RGA法標記植物抗病基因的研究進展
2009-12-28 05:46:44張榮陳歐王振英
天津農業科學 2009年1期
張 榮 陳 歐 王振英
摘要:簡述了植物抗病基因的結構特點,介紹了利用RGA法克隆的抗病基因同源序列及其應用,對RGA法的應用前景進行了展望。
關鍵詞:RGA;抗病基因;結構域
中圖分類號:Q78文獻標識碼: A 文章編號:1006-6500(2009)01-0010-03
Research Advance on Disease-resistant Gene of Plant by RGA Labeling
ZHANG Rong,CHEN Ou,WANG Zhen-ying
(Tianjin Key Lab of Cyto-genetical and Molecular Regulation,College of Chemistry and Life Science,Tianjin Normal University,Tianjin 300387,China)
Abstract:The structure characteristics about plant disease-resistant gene was summarized, the same sequences of cloned disease-resistant genes by RGA and its applications were introduced. At last, the application prospect of RGA was expected.
Key words: RGA;resistance gene;construction domain
植物在長期的發展進化過程中,不可避免的會遭到各種病害的侵襲,在不斷地抵抗其危害的過程中,植物本身形成了一套比較完整的抗病基因序列,然而在不同的植物品系之間,這類抗病基因之間存在著很大的同源性,這類基因被通稱為植物抗病基因(Resistance gene,R基因),并在1992年被首次克隆成功[1]。針對作物、園藝等不同方面的需求,人們對大量植物種類進行了廣泛的實驗驗證,均證明植物的抗病基因同源性確實存在且相對穩定,這逐漸成為實驗研究的一種重要引證和新的實驗方法的源泉。
1 植物抗病基因的結構特點
R基因是在植物抗病過程中抵抗病菌浸染及擴展的有關基因,也就是Flor經典遺傳學的基因對基因(gene-for-gene)假說中所指的與病原菌無毒基因相對應的基因,存在于植物的特定品種中,在植物生長的整個周期或其中某個階段表達的植物抗病品種所特有的一類基因。研究表明,這些抗病基因的氨基酸序列之間具有較高的同源性。
1.1 NBS-LRR結構域的特點
富含亮氨酸重復序列(Leucine Rich Repeats, LRR),與蛋白質之間的互作及信號的轉導存在密切的聯系,包括兩種:一種存于細胞外,一種存在于細胞內。核苷酸結合位點(Nucleotide Binding Site, NBS)—真核生物中具有結合ATP和GTP活性的蛋白,含有三個保守的特征結構域,一是激酶la,即磷酸結合環(P-loop),二是激酶2,三是激酶3a,這兩類保守的區域多同時存在,形成抗病基因的保守結構域,大多數已克隆得到的R基因都有NBS-LRR區域。2004年孫學輝等人利用HMM(Hidden Markov Model)對源于日本晴的粳稻基因組和9311的秈稻基因組的蛋白數據庫進行了搜索,分別獲得了325個和344個富含NBS-LRR類抗病基因的蛋白序列,并得到了與它們相對應的cDNA序列。它們的共同特點是:在它們編碼蛋白的近N端存在NBS,而在它們的近C端則存在LRR[2]。
1.2 TIR、STK及LZ結構域的特點
與果蠅Tol1蛋白及哺乳動物白細胞介素-1受體的細胞外相似的區域(TIR),在植物內部免疫反應過程的信號轉導中起作用。它們能夠與相應的免疫反應的啟動子相結合,從而使相關的基因表達,以達到對病原物的抗性作用[3]。此外,還包括絲氨酸/蘇氨酸蛋白質激酶(Serine/ Threonine Kinase, STK)和亮氨酸拉鏈結構(Leucine Zipper, LZ)等結構域,其中STK包含兩個特征結構域,即DaKXXN和GTaGYXAP(N/E)[4],在抗病過程的信號轉導途徑中,此種激酶借助磷酸化其它的信號分子而達到傳遞其抗性信息的目的。而LZ則是存在于蛋白質一側的亮氨酸殘基結構,每7個氨基酸形成一個重復,而在最后一位上則是亮氨酸或異亮氨酸,在形成二級結構的α-螺旋時,借助疏水作用兩兩相連而形成類似于拉鏈的結構[5]。針對這樣的特點,RGA方法應運而生,通過R基因的保守結構域擴增分離RGA,目的是為了找到更接近于R基因的分子標記并得到更新的植物抗病基因。作為一種簡便易行的分子標記方法,RGA逐漸被大家認可并廣泛地應用于抗病基因的尋找過程中。
2 利用RGA法克隆的抗病基因同源序列及其應用
2.1 RGA技術原理及其特點
RGA分析主要是針對抗病基因的表達產物存在保守區域的特性,人工合成相應的簡并引物,進而以植物的總體DNA或cDNA為模板進行PCR擴增,分離出與其他植物或本種類植物與抗病基因類似的序列,再以PCR擴增所得的產物作為探針,運用RFLP等其他的分子標記方法進行分析,將該克隆產物進行定位,通過在抗病的近等基因系文庫內進行篩選,獲得試驗預期的抗病基因的克隆產物,或進一步明確這些新獲得的基因與已知的抗病基因間的連鎖關系,從而為接下來的克隆或轉化該基因奠定了一定的基礎。
RGA既可以是一種DNA 的分子標記,也可以是個體本身所攜帶的特異抗病基因,在植物中是普遍存在的,多以成簇的方式隨機分布于植物基因組中。它與R基因間可能存在3種關系:第一,分離的RGA可能本身就是一種新的R基因;第二,分離的RGA可能是已經得到的某種R基因的緊密連鎖;第三,還有一種可能就是分離的RGA與R基因根本沒有關系,只是在某些非主導型序列上存在相似性而已,因此分離后期的連鎖性鑒定就顯得尤其重要了。早期的RGA主要是從基因組DNA 中得到的,不可避免地會受到DNA中大量存在的非表達區域和重復區域的干擾,進而影響克隆的效果而得到錯誤的結果。近些年來,研究者逐漸改為從cDNA中克隆得到RGA,再利用特異引物返回到基因組DNA 中獲得基因全長。
雖然在之前的工作中采用RGA法已經在抗病基因克隆方面取得了較好的成績,但是此種方法還存在自身的缺陷:(1)NBS、LRR和STK等保守序列并非只存在于抗性基因中,導致分離得到的RGA并不是都與R基因有關,使得克隆R基因的過程變得復雜;(2)克隆所需的引物在設計時要考慮到抗病基因中的較高同源性,引物設計的好壞嚴重影響PCR產物的實用效果;(3)因R基因在植物中多以成簇的方式存在,因此所得到的與其連鎖的RGA,還要在基因簇中進行篩選,而不是全部接受整個基因簇的基因。
2.2 RGA方法的應用
RGA標記與一般的分子標記如RAPD或RFLP相比,存在其自身的優越性,不僅用于揭示品種的遺傳差異,而且還可以反映品種的功能,有助于選擇品種組合和控制病害。它的應用主要包括:首先,可以作為分子標記用于抗病基因的標記;其次,構建連鎖的遺傳圖譜和輔助植物的抗病育種;再次,RGA也可作為探針用于基因組文庫的篩選或抗病基因的克隆;此外也有直接應用RGA 的簡并引物分析種質資源間的遺傳關系的。
目前,RGA分子標記的方法已經在多種植物中廣泛應用,分離得到的RGA在GeneBank上記錄的已達到200多種。涉及的植物種類廣泛,主要集中在水稻、麥類等糧食作物中。隨著分子生物學技術的飛速發展,植物的抗病性逐漸被大量地應用于育種實踐中,進而避免化學藥物防治對環境和人類造成的破壞和危害。
首先,水稻作為我國的首要糧食類作物,針對其易感染的葉枯病[6]、稻瘟病[7]等主要病害而進行的抗病基因同源序列分析,目前已經成為許多研究者的實驗重點,旨在獲得更接近于R基因的抗病基因序列,從根本上提高其抗病性,進而提高水稻的產量和質量。
其次,小麥作為僅次于水稻的第二類主要作物,研究其抵抗病害的基因作用原理也具有深遠意義。麥類作物普遍感染的葉銹病、白粉病[8],已經成為困擾廣大科學家和全國糧農的首要病害,寄希望于找到更好的抗病基因從而較為徹底的解決此類問題。在麥類植物中應用抗病基因同源序列法,也已經在一定程度上取得了驕人的成績。胡楠等人[9]針對抗白粉病基因Pm4b進行的RGA分析,獲得了穩定的長度為1 321 bp的RGA序列的多態性條帶。
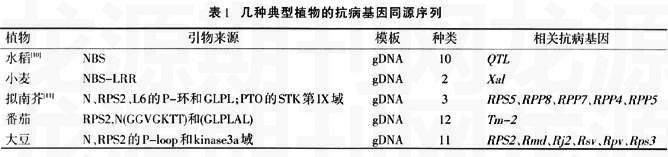
另外,利用抗病基因同源序列法在擬南芥、番茄、大豆等的R基因中都成功地建立了特異的分子標記。幾種典型植物的抗病基因同源序列見表1。
3 RGA研究展望
RGA法作為近些年發展起來的新型分子標記方法,憑借其簡便的技術以及與抗病基因的較高相關性,已得到諸多科學工作者的認可和支持,獲得了不同植物的大量RGA,為進一步的實用價值的創造提供基礎。通過分離RGA的方法克隆R基因或得到與R基因緊密連鎖的分子標記,特別是針對復雜的基因組來說,是相對簡便的一種標記方法。在接下來的研究中應當努力克服其各方面的缺陷,結合其他更加先進的分子生物學技術,從植物體內獲得更多的RGA,進而獲得更加有效的植物抗病基因,為植物育種做出更進一步的探索。
參考文獻:
[1] Johal G S, Briggs S P. Reductase activity encoded by the HM1 disease resistance gene in maize[J].Science,1992,258:985-987.
[2] 孫學輝,路鐵剛,賈士榮,等.水稻富含亮氨酸的重復序列和核苷酸結合位點(LRR-NBS)基因家族的生物信息學分析[J].中國農業科學,2004,37(1):1-7.
[3] 唐軍.Tol1樣受體—一個新發現的介導天然免疫的古老家族(下)[J].國外醫學:免疫學分冊,2001,24(2):59-61.
[4] 王海燕,劉大群,楊文香.植物抗病基因類似序列研究進展及展望[J].河北農業大學學報,2002,25(S1):164-168.
[5] Bostjan K, Johann D. The leucine-rich repeats: a versatile binding motif[J].TIBS, 1996 (8):1757-1771.
[6] 姬廣海,張世光,魏蘭芳,等.云南抗白葉枯病稻種的RGA初析[J].作物學報,2004,30(10):969-974.
[7] 李曄,范靜華,何月秋,等.水稻品種RGA分析與抗瘟性鑒定[J].江西農業大學學報,2007,29(1):11-15.
[8] 王春梅,別同德,陳全戰,等.簇毛麥6V染色體短臂特異分子標記的開發和應用[J].作物學報,2007,33(10):1595 -1600.
[9] 胡楠,伊艷杰,劉紅彥,等.小麥抗白粉病基因Pm4b的RGA分析[J].安徽農業科學,2007,35(21):6379-6380,6430.
[10] Tada Y. PCR-amplified resistance gene analogs link to resistance loci in rice [J]. Breeding Science,1999,49(4):267-273.
[11] Aarts M G, te Lintel H B, Holub E B, et al. Identification of R-gene homologous DNA fragments genetically linked to disease resistance loci in Arabidopsis thaliana [J].Mol Plant Microbe Interact, 1998(11):251-258.
