急性運動中骨骼肌線粒體移動相關基因表達與線粒體動力學的關系
2010-01-04 02:16:20劉慧君姜寧趙斐翟克敏劉洪濤吉力立張勇
天津體育學院學報 2010年2期
關鍵詞:融合
劉慧君姜寧趙斐翟克敏劉洪濤吉力立張勇
●成果報告Original Articles
急性運動中骨骼肌線粒體移動相關基因表達與線粒體動力學的關系
劉慧君1,2,姜寧2,1,趙斐2,翟克敏2,劉洪濤1,吉力立3,2,張勇2,1
目的:擬探討急性運動中骨骼肌線粒體移動基因miro1、融合基因mfn2及分裂基因drp1的動態變化以及之間的相互關聯。方法:以C57 BL/6小鼠一次中等強度負荷跑臺運動(0°,13m/min)為實驗模型,觀察運動中骨骼肌miro1、mfn2、drp1mRNA表達變化,同時測定骨骼肌H2O2生成。結果:(1)120min急性運動過程中miro1mRNA表達均較安靜組顯著增高,E60~E120組mfn2mRNA表達較安靜組顯著降低;E60~E120組drp1mRNA表達較安靜組增高。(2)在運動中各時間點骨骼肌H2O2均較安靜組顯著增高。結論:線粒體移動基因的動態表達可能作為融合分裂基因表達變化的先導,協同應對細胞能量需求急劇變化產生快速應答。
急性運動;骨骼肌;線粒體移動;線粒體融合分裂
線粒體是具有高度動態結構和能移動的細胞器,為適應細胞不同生理功能、細胞內信息傳遞及不同部位能量需要的變化,其在細胞中持續地進行著移動、分裂與融合、合成與降解,其形態、分布、數量都在不斷地改變[1-3],從而維持適宜的動態平衡。目前已發現一些與線粒體形態動力學密切相關蛋白,如調控線粒體內、外膜融合的線粒體融合蛋白1/2(Mfn1/2)、調控線粒體分裂的動態相關蛋白1(Drp1)、線粒體移動相關蛋白Miro1/2等[4]。研究發現,線粒體形態和結構的動態變化深刻地影響著線粒體功能、細胞能量代謝、發育、凋亡、衰老、線粒體DNA復制等生命活動以及神經系統疾病、代謝性疾病等多種疾病發生過程[5-6]。如線粒體過度分裂將損害線粒體的能量代謝,融合與分裂的動態平衡能力下降直接導致細胞能量代謝紊亂,引發代謝性疾病[7];線粒體移動的變化、線粒體定位異常與機體發育、多種神經系統疾病密切相關[8]。
研究證實,能量需求旺盛的組織如心肌、骨骼肌中線粒體呈豐富的網絡化結構。目前運動中線粒體形態動力學密切相關蛋白的研究尚少,運動過程中骨骼肌細胞能量需求急劇變化時,線粒體移動、融合分裂基因表達及與線粒體能量代謝的關聯和意義仍不清楚。本實驗擬以急性運動為模型,研究骨骼肌在能量需求急劇變化過程中,線粒體移動與融合分裂基因表達的動態變化,并初步探討線粒體移動、融合、分裂基因動態表達之間的可能關系及其生理意義。
1 材料與方法
1.1 實驗動物與分組
雄性C57 BL/6小鼠40只(19~21 g),7~8周齡,購自北京維通利華實驗動物技術有限公司(SPF級),于正式實驗前一周購入,在本實驗室分籠飼養,自由飲食,飼料為標準嚙齒類飼料。飼養環境為21~24℃,相對濕度45%~55%,每日光照12 h。將小鼠隨機分為5組,組間體重無顯著性差異。其中1組為安靜對照組(R,n=8),4組為急性運動組,分別以不同組代表各時間點:運動30 min組(E30,n=8)、運動60 m in組(E60,n=8)、運動90min組(E90,n=8)、運動120min組(E120,n=8)。
1.2 急性運動模型
以跑臺運動作為運動應激模型。實驗前所有動物均未進行過跑臺運動。正式實驗前,動物先在跑臺上進行5min跑步以適應跑臺環境(0,5 m/min)。運動組正式實驗采用中等強度運動(0,13m/min)。
1.3 骨骼肌樣品制備
安靜對照組動物于安靜狀態、其他組于運動中不同時間點(E30、E60、E90、E120)即刻頸椎脫臼處死,迅速取出下肢骨骼肌,部分樣本液氮速凍,-80℃凍存備用進行基因測定;另外部分骨骼肌樣本立即進行H2O2測定。以上操作均在冰浴中進行。
1.4 骨骼肌m iro1、m fn2和drp1mRNA熒光定量PCR分析
使用Trizol(Invitrogen)提取骨骼肌總RNA,通過0.8%的瓊脂糖電泳和紫外分光光度計確定總RNA完整性和純度;按照RevertAid First Strand cDNA Synthesis Kit(Fermentas)說明書進行逆轉錄反應。采用Applied Biosystems StepOne Real-Time PCR System(BIO-RAD)進行熒光定量PCR反應,引物由寶生物工程(大連)有限公司合成,熒光定量PCR條件:預變性95℃/30 s;95℃/5 s,60℃/30 s,共40個循環(見表1)。根據SYBR~Premix Ex Taq Kit(BIO-RAD)說明書配制反應體系,目的基因的相對變化量(Fold Change)=2-Δ(ΔCT),其中ΔCT=CTtarget-CT18s,Δ(ΔCT)=ΔCTstimulated-ΔCTcontrol。
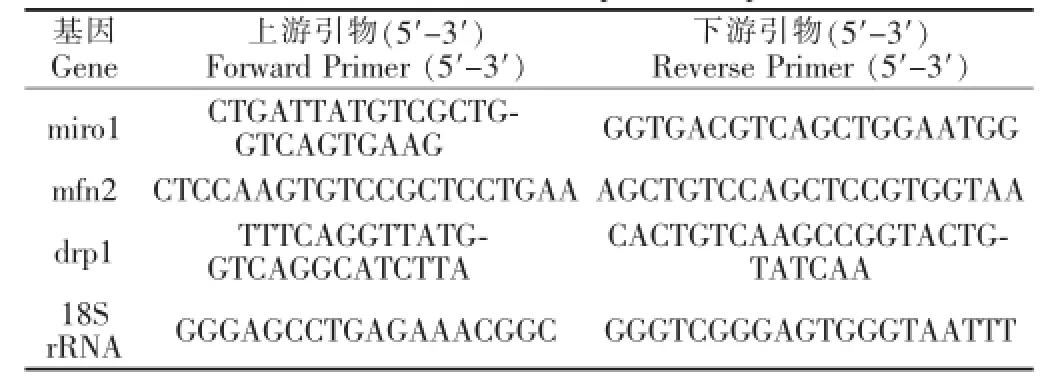
表1 熒光定量PCR引物序列Table 1 Real-time PCR prim er sequences
1.5 骨骼肌H2O 2測定
根據H2O2與鉬酸銨作用生成穩定的黃色過氧鉬酸絡合物的原理,在可見-紫外分光光度計(Beckman Co,USA)波長405 nm測定其絡合物生成量可計算H2O2的濃度。組織剪碎,去除筋膜、脂肪;加入生理鹽水用玻璃勻漿器勻漿;2 500 r/min 4℃離心10min,取上清,嚴格按照南京建成生物工程研究所試劑盒說明書操作進行檢測。
1.6 統計學分析
2 結果
2.1 m iro1和m fn2、drp1 m RNA含量的變化
如圖1所示,120min運動過程中miro1mRNA表達均較安靜組顯著增高,E30、E60、E90、E120組分別較安靜組增高32.8%、107.6%、63.8%及44.8%,E60組達峰值。如圖2所示,E60、E90、E120組m fn2mRNA表達均較安靜組明顯減少,分別較安靜組降低了34.6%、69.3%及60.4%。如圖3所示,E60、E90、E120組drp1 mRNA表達均較安靜組分別增高20.5%、35.3%及22.0%。
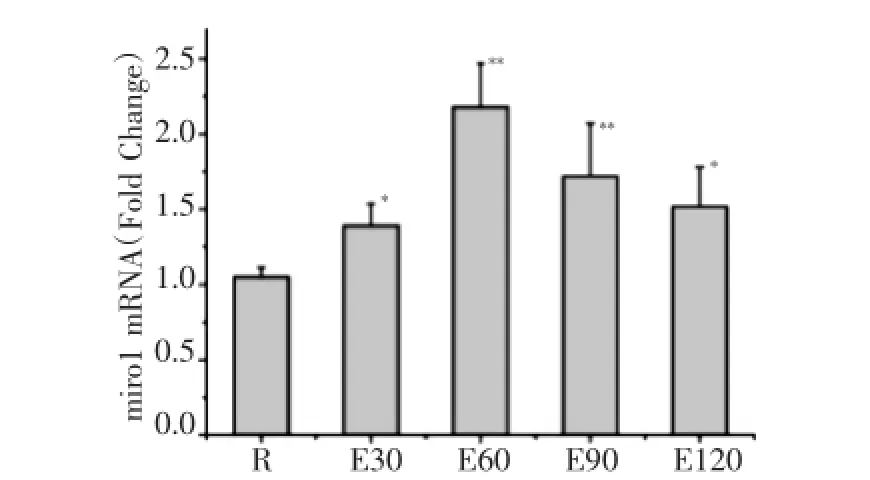
圖1 急性運動中小鼠骨骼肌m iro1m RNA表達的動態變化(與安靜組比較,*P<0.05**P<0.01,n=8,以下同)Fig.1 Dynam ics expression ofm iro1 m RNA in m ouse skeletal muscle during acute exercise
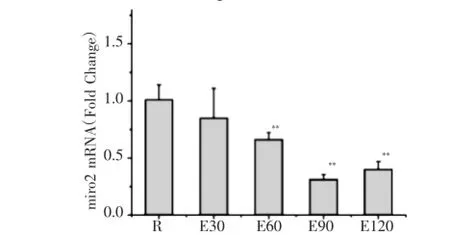
圖2 急性運動中小鼠骨骼肌m fn2mRNA表達的動態變化Fig.2 Dynamics exp ression ofm fn2m RNA inmouse skeletal muscle during acute exercise
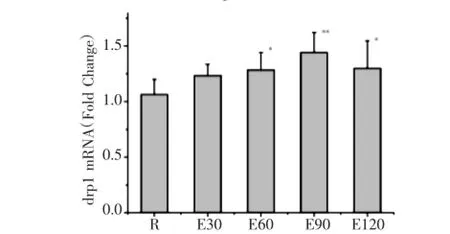
圖3 急性運動中小鼠骨骼肌drp1 m RNA表達的動態變化Fig.3 Dynam ics expression ofd rp1m RNA inmouse skeleta l m uscle during acute exercise
2.2 骨骼肌H2O2濃度變化
如圖4所示,120 min運動過程中骨骼肌H2O2均較安靜組顯著增高,其中E30、E60和E90組H2O2持續增高,分別較安靜組增高了22.1%、26.7%、37.6%,E90組達峰值,E120組比E90組略有降低但仍明顯高于安靜組。
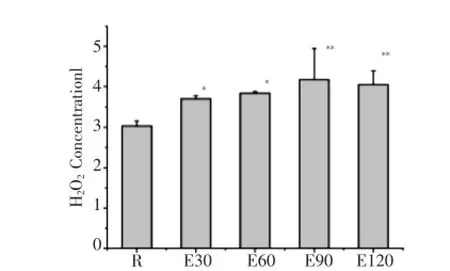
圖4 急性運動中小鼠骨骼肌H2O2濃度動態變化Fig.4 Dynamics ofH2O2 concentration inmouse skeleta lmuscle during acute exercise
3 討論
近年來的研究表明,線粒體在細胞中不斷地移動、融合與分裂,其形態分布不斷地變化,以應對不同生理功能和細胞內不同區域的能量需要,形成動態平衡的“整合性應答”。線粒體形態分布的動態變化對線粒體功能、細胞能量代謝、凋亡等生命活動具有重要的生理意義。如L6E9肌管細胞中,mfn2表達下調可降低呼吸鏈復合體I、Ⅱ、III、V亞單位的表達,而m fn2表達上調則增加這些基因的表達,促進線粒體能量代謝[9]。通過RNA干擾減少HeLa細胞中drp1表達,使線粒體呼吸速率降低、ATP合成減少,線粒體膜流動性增加[10]。線粒體傾向于聚集在特殊的亞細胞部位提供能量、參與如鈣緩沖等胞內信號傳遞[3,[12]。Waterham HR等報道了首例Drp1突變的病例,該突變為致死突變,可引起線粒體和過氧化物酶體形態改變,并引起出生后大腦發育障礙、視神經萎縮、高乳酸血癥等一系列嚴重的并發癥[13]。由此可見,線粒體形態動力學變化參與細胞內信息傳遞、能量代謝,與細胞生理病理密切相關。
目前,運動過程中骨骼肌這一能量代謝旺盛的組織中線粒體移動、融合分裂基因表達的研究鮮見報道。本研究發現:120 m in急性運動過程中E60~E120組m fn2 mRNA表達均較安靜組明顯降低;E60~E120組drp1mRNA表達均較安靜組增高。并且急性運動過程中m fn2 mRNA表達顯著下調,drp1 mRNA表達上調。以上融合分裂基因動態變化的結果與本實驗室前期報道首次發現急性運動中大鼠骨骼肌Mfn1/2 mRNA表達及蛋白含量減少,分裂蛋白Fis1mRNA表達及蛋白含量增加的結果趨勢一致[14],再次證實了線粒體的動態變化可能適應了細胞能量代謝的需求。正常的線粒體分裂融合動態變化直接影響線粒體的氧化磷酸化功能[9-10]。一般認為線粒體融合成網絡利于能量和信息在不同線粒體中傳遞,線粒體內容物及mtDNA交換互補[15];線粒體分裂利于線粒體根據能量需求重新分布,提高線粒體能量代謝速率,合成更多ATP,同時保護線粒體免于遭受過多損傷[10,16]。以上研究提示:急性運動中線粒體融合受抑制而趨于分裂,從而可能提高線粒體能量代謝速率,以滿足細胞對能量的需求。
近年來研究發現,線粒體移動與線粒體融合分裂之間具有密切關聯。如HeLa細胞中,Dynein/dynactin復合體(線粒體移動馬達蛋白)通過影響drp1在胞內重新分布調控線粒體形態分布,當Dynein功能破壞導致線粒體分布于核周且形成長的分支狀結構[17]。H9C2及原代腦皮層神經細胞中,細胞胞漿中游離鈣離子濃度([Ca2+]i)靜息水平時,過表達Miro增強線粒體移動并促進線粒體趨于融合;[Ca2+]i升高或鈣振蕩時,過表達Miro促進線粒體趨于分裂[18]。以上研究提示,線粒體移動很可能作為線粒體融合與分裂介導的網絡結構變化的基礎,通過影響融合分裂,進而影響線粒體能量代謝。本研究發現:120min急性運動中miro1mRNA表達均較安靜組顯著增高,且m iro1基因表達的改變先于m fn2、drp1基因表達變化。提示:急性運動中線粒體移動增加,且很可能通過影響線粒體融合與分裂基因,促進線粒體網絡結構趨于分裂以利于線粒體重新分布,以應對細胞能量需求的急劇變化。
目前,有關線粒體形態分布的調控機制尚不清晰。相關研究發現活性氧(ROS)可以調控線粒體移動融合分裂動態改變。ROS是把雙刃劍,較低濃度的H2O2等活性氧在調節細胞增殖、分化、凋亡以及細胞信號轉導中起著重要作用[19];高濃度的ROS能夠誘導細胞死亡[20],氧化脂質、蛋白、DNA等多種細胞成分,從而對細胞完整性造成威脅[21]。Barsoum發現NO可以誘導線粒體分裂使線粒體活性下降,ATP產生減少,ROS增多,對神經元細胞產生毒性引起神經損傷病變[22]。原代前腦神經細胞中,NO誘導線粒體移動停止,同時線粒體膜電位降低[23]。本實驗結果表明,120min急性運動過程中各組骨骼肌H2O2均較安靜組顯著增高。根據前述實驗結果及相關研究分析提示:一次急性運動中H2O2生成急劇增多,同時線粒體移動基因表達迅速增加,線粒體通過移動分布的變化應對能量需求改變產生“早期快速應答”,在此基礎上線粒體融合分裂網絡結構動態平衡變化產生“整合性應答”。在這一應答過程中,線粒體的動力學變化導致細胞內線粒體不斷地進行重構,可能有助于線粒體與細胞胞漿和其他細胞器之間的物質、信息和能量交換以滿足細胞對能量的快速需求。
目前,對運動過程中線粒體移動、融合分裂的研究還不夠深入,線粒體融合分裂之間的偶聯機制及線粒體移動與融合分裂的關系以及具體的調控機制和生理意義尚不清楚。骨骼肌線粒體移動基因變化是否作為融合分裂基因表達變化的先導影響線粒體動力學;ROS是否作為信號分子參與調控線粒體動力學改變及其與線粒體能量代謝的關系等問題還有待進一步深入探討。
[1]Heymann JA,Hinshaw JE.Dynaminsataglance[J].Journalof Cell Science,2009,122(19):3427-3431.
[2]Liu X G,Weaver D,Shirihai O,et al.Mitochondrial'kiss-and-run':interplay betweenmitochondrialmotility and fusion-fission dynamics[J].The EMBO Journal,2009,28(20):3 074-3 089.
[3]Yi M,Weaver D,Hajnóczky G.Control ofmitochondrialmotility and distribution by the calcium signal:ahomeostatic circuit[J].Journal of Cell Biology,2004,167:661-672.
[4]馬國棟,張勇.運動與解偶聯蛋白3(UCP3)介導的線粒體活性氧代謝調控研究進展[J].西安體育學院學報,2007,(4):71-78.
[5]夏云健,時慶德,蔣春筍,等.運動性疲勞狀態下大鼠心肌線粒體氧化磷酸化偶聯的變化[J].天津體育學院學報,2002,(2):22-24.
[6]常波.運動源性自由基對大鼠腎臟線粒體游離鈣和ATP合成能力的影響[J].上海體育學院學報,2006,(2):76-79,83.
[7]Chan DC.MitochondrialFusion and Fission inMammals[J].AnnualReview of Celland Developmental Biology,2006,22:79-99.
[8]Guo X,Macleod G.T,Wellington A,etal.The GTPase dMiro is required for axonaltransportofmitochondriatoDrosophilasynapses[J].Neuron,2005,47(3):379-393.
[9]Pich S,Bach D,Briones P,et al.The Charcot-Marie-Tooth type 2A gene product,Mfn2,up-regulates fueloxidationthrough expression of OXPHOS system[J].Human Molecular Genetics,2005,14(11):1 405-1 415.
[10]Benard G,Bellance N,James D,et al.Mitochondrial bioenergetics and structural network organization[J].Journal of Cell Science,2007,120:838-848.
[11]Park M K,Ashby M C,Erdemli G,et al.Perinuclear and sub-plasmalemmal mitochondria have distinct functions in the regulation of cellular calcium transport[J].The EMBO Journal,2001,17:1 863-1874.
[12]Bach D,Pich S,Soriano FX,etal.Mitofusin-2 determinesmitochondrial network architecture and mitochondrialmetabolism.A novel regulatory mechanism altered in obesity[J].Journal of Biological Chemistry,2003,278:17190-17197.
[13]Waterham H R,Koster J,van Roermund CW,et al.A lethal defect of mitochondrial and peroxisomal fission[J].The New England Journal of Medicine,2007,356:1 736-1 741.
[14]Ding H,Jing N,Liu H J,et al.Response of Mitochondrial Fusion and Fission Protein Gene Expression to Exercise in Rat SkeletalMuscle[J].Biochim Biophy Acta,2009,8:7.
[15]Legros F,Lombes A,Frachon P,etal.Mitochondrial fusion in human cells is efficient,requires the inner membrane potential,and ismediated by mitofusins[J].Molecular Biology of the Cell,2002,13(12):4343-4 354.
[16]Pletjushkina O Y,Lyamzaev K G,Popova E N,et al.Effect of oxidative stresson dynamicsofmitochondrial reticulum[J].Biochim Biophys Acta,2006,1757:518-524.
[17]Varadi A,Johnson-Cadwell L I,Cirulli V,et al.Cytoplasmic dynein regulates the subcellular distribution ofmitochondria by controlling the recruitmentof the fission factor dynamin-related protein-1[J].Journal of CellScience,2004,117:4 389-4 400.
[18]Saotome M,Safiulina D,Szabadkai G.Bidirectional Ca2+-dependent controlofmitochondrialdynamicsby theMiroGTPase[J].Proceedings of the National Academy of Sciences of the United States of Americ(PNAS),2008,105(52):20 728-20 733.
[19]Peus D,BeyerLe A,VasaM,et al.Antipsoriatic drug anthralin induces EGF receptor phosphorylation in kerationocytes:requirement for H2O2 generation[J].Experimental Dermatology,2004,13(2):78-85.
[20]Orzechowski A,Grizard J.Jank M.et al.Dexamethasone-mediated regulation ofdeathand diferentiation ofmusclecells Ishydrogen peroxide involved in theprocess[J].Reproduction Nutrition Development,2002,42(3):l97-216.
[21]Scherz-Shouval R,Elazar Z.ROS,mitochondria and the regulation of autophagy[J].Trends in Cell Biology,2007,17(9):422-427.
[22]Barsoum M J,Yuan H,Gerencser A A,et al.Nitric oxide-induced mitochondrial fission is regulated by dynamin-related GTPases in neurons[J].The EMBO Journal,2006,25(16):3 900-3 911.
[23]RintoulGL,BennettV J,PapaconstandinouNA,etal.Nitricoxide inhibits mitochondrialmovementin forebrain neuronsassociatedwith disruption of mitochondrialmembranepotential[J].JournalofNeurochemistry,2006,97(3):800-806.
The Relationship of M itochond rial M otility-related Gene Expression and M itochond rial Dynam ic during a Bout of Acute Exercise
LIU Huijun1,2,JIANG Ning2,1,ZHAO Fei2,ZHAIKemin2,LIU Hongtao1,JILili3,2,ZHANG Yong2,1
(1.Institute of Health and Environmental Medicine,Academy of Military Medical Sciences,Tianjin 300050,China;2.Tianjin Key Laboratory of Exercise Physiology and Sports Medicine,Tianjin University of Sport,Tianjin 300381,China;3 Dept.of Kinesiology,University of Wisconsin-Madison,Madison,WI 53706,USA)
Objective:The purpose of this study was to investigate the gene expressions ofmiro1,m fn2,drp1 during an acute bout of prolonged exercise and to analysis the relations of the gene expressions.Methods:C57 BL/6 mouse underwent amidd le intensity treadmill running with 0,13m/min.Gene expressions ofmirol,mfn2 and drp1 mRNA were detected,skeletalmuscle H2O2concentration was alsomeasured.Results:First,compared with resting group(R), miro1 mRNA contents were significant increased during 120 min of exercise;mfn2 mRNA contentswere significant decreased in groups E60~E120;drpl mRNA expression increased in groups E60~E120.second,H2O2contents of skeletalmuscle were progressively increased during 120min of exercise.Conclusion:Mitochondrialmotilitymaybe the base ofmitochondrial fusion-fission,the gene expression ofmitochondrial fusion-fission and motility proteins in skeletalmusclemight respond rapidly tomatch the energy demand during exercise.
acute exercise;skeletalmuscle;mitochondrialmotility;mitochondrial fusion-fission
G 804.2
A
1005-0000(2010)02-0118-04
2010-01-07;
2010-01-31;錄用日期:2010-02-01
國家自然科學基金項目(項目編號:30771048);天津市社會發展重大科技攻關項目(項目編號:05YFGDSF02100)
劉慧君(1973-),女,天津市人,天津體育學院助理研究員,軍事醫學科學院在讀博士研究生。通訊作者:張勇(1956-),男,湖北人,天津體育學院教授,軍事醫學科學院博士生導師。Email:yzhang@tjus.edu.cn
1.軍事醫學科學院衛生學醫學研究所,天津300050;2.天津體育學院天津市運動生理與運動學重點實驗室,天津300381;3.Deportmentof kinesiology,University ofWisoonsin-Madison,Madison,WI53706,USA。
猜你喜歡
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
數學年刊A輯(中文版)(2022年4期)2022-02-16 08:17:34
今日農業(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
無線電通信技術(2021年4期)2021-07-13 08:58:28
無線電通信技術(2021年3期)2021-06-08 03:33:48
中學生數理化(高中版.高考數學)(2021年1期)2021-03-19 08:28:38
無線電工程(2020年11期)2020-10-29 01:25:46
現代出版(2020年3期)2020-06-20 07:10:34
福利中國(2015年4期)2015-01-03 08:03:38
