鹽脅迫對海棠等3樹種光合能力及色素變化的影響
2010-05-09 06:57:52孫方行孫明高
浙江林業科技 2010年2期
關鍵詞:植物
孫方行,周 勃,孫明高
(1. 河南城建學院,河南 平頂山 467044;2. 山東農業大學,山東 泰安 271018)
鹽脅迫對海棠等3樹種光合能力及色素變化的影響
孫方行1,周 勃1,孫明高2*
(1. 河南城建學院,河南 平頂山 467044;2. 山東農業大學,山東 泰安 271018)
以2年生海棠(Malus spectabilis)、石榴(Punica granatum)和桃樹(Amygdalus persica)的盆栽苗為試驗材料,測定它們在不同濃度NaCl脅迫下的光合能力及色素變化。結果表明,3樹種的耐鹽性依次為海棠 > 石榴> 桃樹;鹽脅迫下3種非鹽生樹種凈光合速率(Pn)和氣孔導度(Gs)下降,胞內CO2濃度(Ci)變化沒有規律,非氣孔因素是導致光合下降的限制因子;鹽脅迫導致3樹種葉綠素含量下降,但葉綠素a/b的變化各不相同。
鹽脅迫;光合能力;色素;桃樹;石榴;海棠
我國是一個人口多、人均土地資源占有相對較少的農業國,在并不豐裕的土地資源中包含有大量的鹽堿地,它們分布在我國西北、東北及沿海地區,如何開發和利用好這些鹽堿地,對于提高人民生活水平,改善生態環境具有十分重要的意義。
植物耐鹽性[1~2]及其對鹽脅迫響應機理研究日趨受到重視[3]。光合作用是植物生產的最基本過程,研究鹽逆境下光合作用變化規律和機理,對于提高植物抗鹽性具有十分重要的意義。鹽逆境會引起光合速率降低、生長減慢、氣孔非均勻關閉等[4~5]。以往關于植物抗鹽性的研究多局限在鹽脅迫后幾天到一周的較短時間內的變化,難以反映植物對鹽分長期脅迫的響應,同時這方面研究多集中于農作物,較少涉及果樹[6]。本文以海棠、石榴和桃樹的2年生實生苗為試驗材料,對它們在鹽脅迫下光合作用及葉綠素含量的變化進行了對比研究,以期探討它們在鹽脅迫下光合作用的變化規律及其與光合色素的關系。
1 試驗材料與方法
1.1 試驗材料
本研究以桃樹(Amygdalus persica)、石榴(Punica granatum)、海棠(Malus spectabilis)的2年生實生苗為材料。2007年12月從山東農業大學教學基地購得上述苗木各120株,假植于該基地林學站苗圃,并作好苗木越冬工作。2008年3月挑選長勢相對一致的苗木植于規格為30 cm×20 cm×26 cm的花盆中,每盆盛土10 kg,每樹種每處理5株苗木,共計60株。
1.2 材料處理
2008年7月1日做好苗木的防雨措施。2008年7月5日分別用0(ck)、2、4和6 mg/g的NaCl溶液處理苗木,每隔3 d澆1次NaCl溶液,每次每盆1 L。加鹽過程中盡量減少鹽分的損失,每次澆水時滲漏出的水分要重新回灌入花盆中。桃樹加8次NaCl溶液后,6 mg/g處理苗木出現死亡停止加鹽。在第8次加鹽結束后3 d隨機選取植株相同部位葉片進行試驗測定。
1.3 試驗方法
1.3.1 光合參數的測定 采用美國產LI-6200型光合測定儀,于10:00測定各處理植株的4 ~ 7片成熟葉片的光合能力,測定時光強為1 000 umol·m-2·s-1,CO2濃度為450 umol/mol,溫度為29℃,O2濃度為21%,測定指標包括光合速率(Pn)、胞內CO2濃度(Ci)、氣孔導度(Gs)。Ls利用Berry等[7]的方法計算:
Ls=1-Ci-Ca
式中,Ca為空氣中的CO2濃度。
1.3.2 葉綠素的測定 每樹種每處理隨機取葉15片,用直徑為0.6 cm的打孔器打出葉圓片15片。用10 mL 96%乙醇溶液浸提葉綠素。浸泡48 h后,再以參比作為對照,用UV-160紫外分光光度計測其吸光值,求得葉綠素的濃度、葉綠素和類胡蘿卜素的含量。
1.3.3 土壤中NaCl含量的測定 在苗木采樣的同時,采集土樣,并立即測定土壤含水量。用去離子水提取土壤中可溶性鹽分(m水∶m干土=5∶1)。Na+用原子吸收法測定,Cl-用AgNO3法測定,結果見圖1。
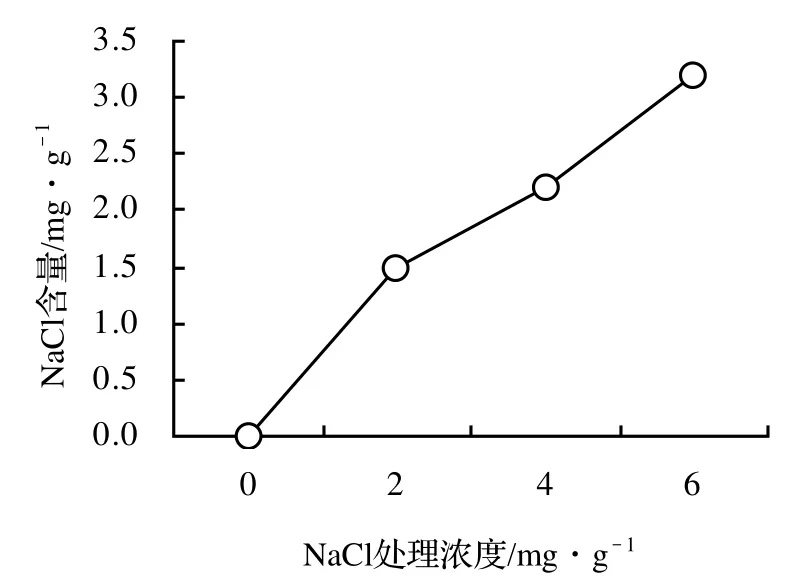
圖1 各處理土壤中NaCl的含量Figure1 NaCl concentration in the soil of former planting location
2 結果
2.1 鹽脅迫對3樹種Gs、Pn和Ci的影響
3樹種的Gs都隨著鹽分處理濃度的升高而下降,并且三者的變化趨勢相似(圖2)。其中桃樹的Gs下降最大,達到75.8%,而海棠的Gs下降最小,為57.2%。但是3樹種在2 mg/g水平時無明顯受害癥狀,表明氣孔部分關閉是這幾種非鹽生樹種對鹽脅迫的一個快速而敏感的反應。
鹽脅迫下3樹種的凈光合速率如圖3所示。3樹種Pn隨著鹽分處理濃度的變化趨勢與Gs相似。桃樹和石榴的Pn隨著鹽分處理濃度升高而變化較大,分別降低了99.3%和91.1%,而海棠的只降低了62.7%。3樹種凈光合速率在低鹽脅迫上就已明顯降低,說明凈光合速率和氣孔導度一樣,對鹽脅迫反應非常敏感。
3樹種的Ci都表現出先下降后上升的趨勢(圖4)。這種趨勢正好與氣孔限制值(Ls)的變化趨勢相反(圖5)。由此可知鹽分脅迫對Ci和Ls的影響存在差異。
2.2 鹽脅迫下3樹種葉片色素的變化
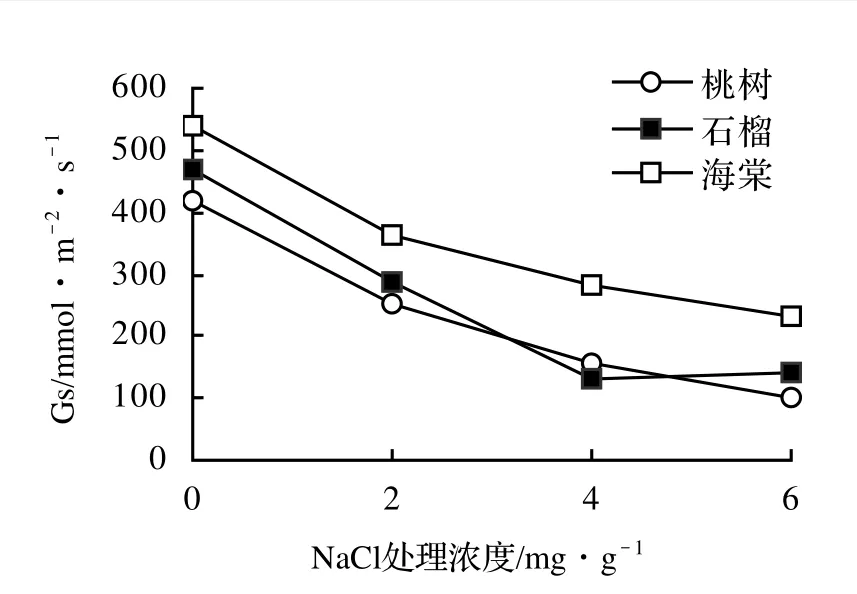
圖2 鹽脅迫下3樹種Gs的變化Figure 2 The changes of Gs of three tree species under different salt stress
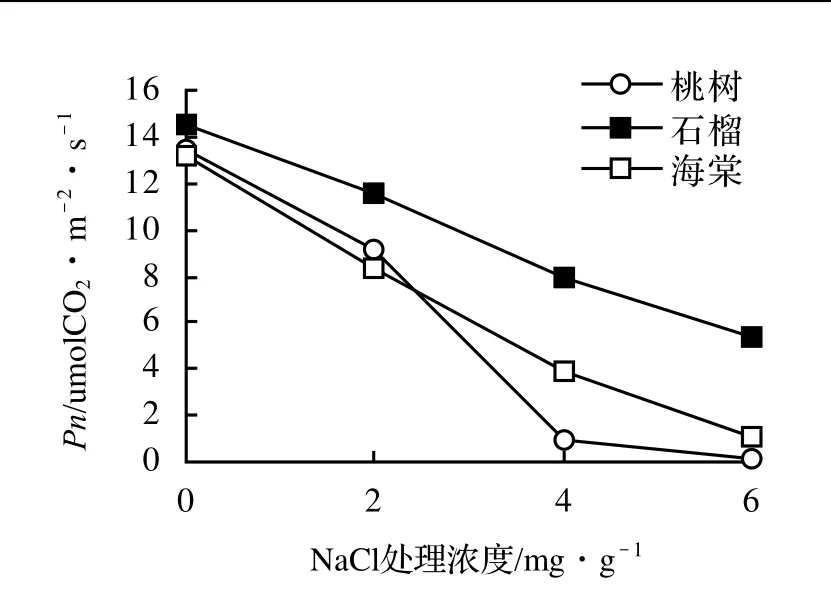
圖3 鹽脅迫下3樹種Pn的變化Figure 3 The changes of Pn of three tree species under different salt stress
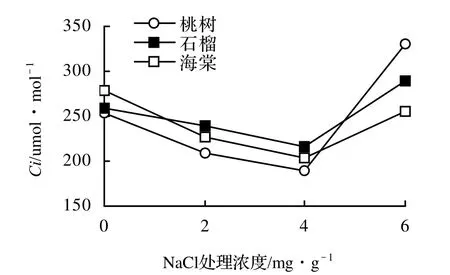
圖4 鹽脅迫下3樹種Ci的變化Figure 4 The changes of Ci of three tree species under different salt stress
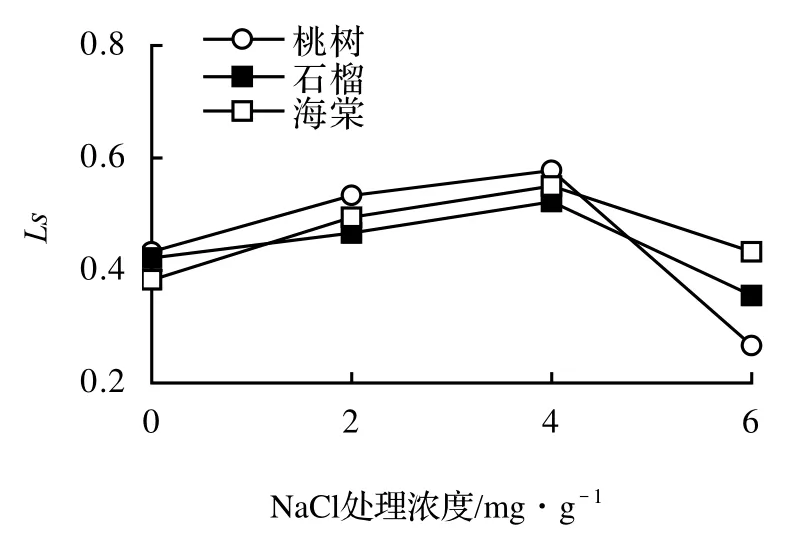
圖5 鹽脅迫下3樹種Ls的變化Figure 3 The changes of Ls of three tree species under different salt stress
由表1中可知,鹽脅迫下3樹種葉綠素含量比對照降低,鹽脅迫強度越大,葉綠素含量越低。張凌云[8]關于蘋果砧木的研究中都發現鹽脅迫下葉綠素含量的降低與鹽脅迫下葉綠素酶的分解作用有關。本試驗中低鹽處理(2 mg/g)水平時葉綠素含量與對照相比只有少量下降,但是當鹽分處理濃度達到6 mg/g時,3樹種的葉綠素含量都有明顯的下降,海棠、石榴和桃樹分別只有對照的80.0%、68.8%和71.7%。
隨著鹽分處理濃度的升高,3樹種類胡蘿卜素(Car)含量變化趨勢與葉綠素含量的變化趨勢相似,都是隨著鹽分處理濃度的升高而降低。葉綠素a/b值因樹種而不同,桃樹葉片在鹽脅迫下無變化,石榴在高鹽濃度處理6 mg/g水平上降低,海棠在受鹽脅迫后葉綠素a/b值升高。
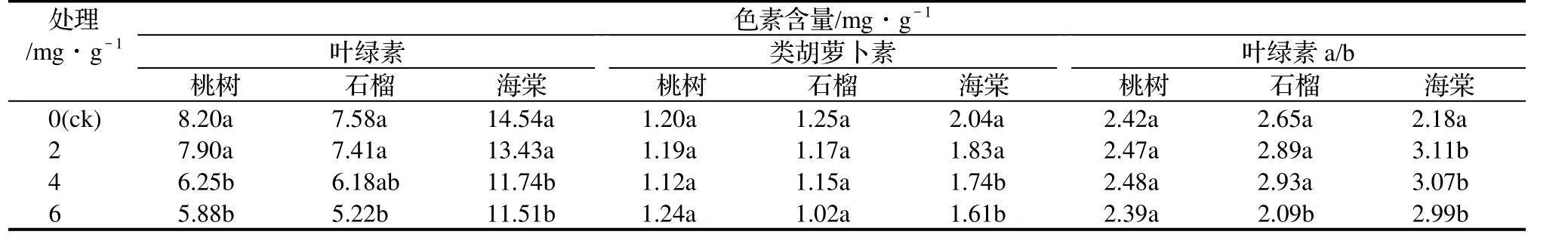
表1 鹽脅迫下3樹種光合色素的變化Table 1 The changes of photosynthetic pigment of the three tree species under salt stress
3 結論與討論
3.1 對3樹種光合限制因子的分析
在鹽脅迫下,植物光合速率降低的因子一般認為有氣孔限制和非氣孔限制兩種[7]。采用鄒琦[9]方法分析,桃樹等3樹種葉片胞內CO2濃度在2、4 mg/g水平時Pn、Gs和Ci比對照降低,并且Ls是升高的趨勢,說明低鹽脅迫引起的氣孔關閉是光合降低的限制因子。而在高鹽處理Pn比對照降低,Ci又出現升高的趨勢,如桃樹的Ci值達到330 umol/mol,比對照增加了30.0%,而此時Ls又出現下降趨勢,說明非氣孔因素是Pn下降的主要因素。鹽脅迫影響了光合作用其它環節,雖然胞內CO2積累,其濃度值升高,但是并不能被植物充分利用,具體原因有待于進一步研究。
3.2 對3樹種色素的分析
植物的類囊體膜是葉綠體光能吸收、傳遞和轉換的結構基礎,而色素是類囊體膜的重要組成成分,是光能的受體。葉片中光合色素含量是反映植物光合能力的一個重要指標,其中葉綠素a有利于吸收長波光,葉綠素b有利于吸收短波光,Car既是光合色素,又是內源抗氧化劑,除在光合作用中具有一定的功能外,在細胞內還可吸收剩余光能,淬滅活性氧,從而防止膜脂過氧化,葉綠素a/b值的變化,能反映葉片光合活性的強弱[11]。許多研究證實,環境因子的改變可以引起光合色素含量的變化,進而引起光合功能的改變[12]。
本試驗通過控制土壤中鹽分濃度從而改變其生長環境,結果發現3樹種的葉綠素含量呈現下降趨勢。3樹種的葉綠素含量下降表明鹽脅迫影響了樹種的葉綠素合成從而導致葉片失綠。一般研究表明,鹽脅迫下植物葉片中葉綠素含量下降[13],其主要原因是由于NaCl能促進葉綠素酶活性,使葉綠素分解[14];另外由于類胡蘿卜素含量的降低,減少了對活性氧的淬滅,導致細胞內積累較多的氧自由基,破壞葉綠體膜結構,加速葉綠素的分解,從而降低光合作用中光能的吸收和傳遞。3個樹種中僅有海棠在高鹽分濃度(6 mg/g)脅迫下的葉綠素a/b仍大于對照,其增幅為37%,同時在低鹽濃度(2、4 mg/g)下,其葉綠素a/b增幅更大,表明在鹽分濃度脅迫下海棠通過維持其高的葉綠素a/b,盡量減小鹽分脅迫對其生長、光合等的影響。
[1] 夏富才,姜貴全,陸靜梅. 鹽生植物抗鹽結構機理研究進展[J]. 通化師范學院學報,2002,23(2):67-69.
[2] 王文卿,林靜. 紅樹植物體內元素分布特點與抗鹽機理[J]. 林業科學,2003,39(4):30-36.
[3] Story R,Walker R R. Citrus and salinity [J]. Sci Hor,1998(78):39-81.
[4] 張川紅,沈應柏,尹偉倫,等. 鹽脅迫對幾種苗木生長及光合作用的影響[J]. 林業科學,2002,38(2):27-31.
[5] 惠紅霞,許興,李守明. 鹽脅迫抑制枸杞光合作用的可能機理[J]. 生態學雜志,2004,23(1):5-9.
[6] 杜中軍,翟衡,潘志勇,等. 鹽脅迫下蘋果砧木光合能力及光合色素的變化[J]. 果樹學報,2001,18(4)200-203.
[7] 鄭炳松,謝芳,潘惠萍. 無土栽培觀葉植物凈光合速率及環境因子動態變化的初步研究[J]. 浙江林業科技,2001,21(1):13-16.
[8] 張凌云,翟衡,張憲法,等. 鐵鹽脅迫對蘋果砧木葉中色素和離子含量的影響[J]. 植物生理學通,2002,38(1):17-19.
[9] 鄒琦. 作物在水分逆境下的光合作用[J]. 作物雜志,1994(5):1-4.
[10] 許大全. 氣孔的不均勻關閉與光合作用的非氣孔限制[J]. 植物生理學通訊,1995,31(4):246-252.
[11] 蔣文偉,劉彤,溫國勝,等. 城市觀賞樹種桂花的光合特性初步研究[J]. 浙江林業科技,2003,23(5):18-21.
[12] 卓仁英,陳益泰. 五種愷木屬植物的光合特性初步研究[J]. 浙江林業科技,2004,24(6):1-4.
[13] 刁豐秋,章文華,劉友量. 鹽脅迫對大麥葉片類囊體膜組成和功能的影響[J]. 植物生理學報,1997,23(2):105-110.
[14] 劉家堯,衣艷君,張其德. 鹽脅迫對不同抗鹽性小麥葉片熒光誘導動力學的影響[J]. 植物學通報,1998,15(2):46-49.
Changes of Photosynthetic Capability and Pigment Contents of Malus spectabilis, Punica granatum and Amygdalus persica Under Salt Stress
SUN Fang-xing1,ZHOU Bo1,SUN Ming-gao2*(1. Henan University of Urban Construction, Pingdingshan 467044, China; 2 Shandong Agriculture University, Tai’an 271018, China)
The changes of photosynthetic capability and pigment content of 2-year potted Malus spectabilis, Punica granatum and Amygdalus persica were determined under different salt stress. The result showed that salt resistance was as follows: M. spectabilis>A. persica>P. granatum. The Pn and Gs of tested seedlings decreased, but the Ci had no change rule. Chlorophyll content of the seedlings was increased slowly, but the changes of Chla/Chlb were different.
salt stress; photosynthetic capability; pigment; Amygdalus persica; Punica granatum; Malus spectabilis
S718.43
1001-3776(2010)02-0036-05
2009-06-12;
2010-02-18
國家科技攻關計劃重大專項“中國森林網絡體系的研究”(2002BA516A15-04)
孫方行(1981-),男,山東萊蕪人,講師,碩士研究生,從事林木栽培生理研究;*通訊作者。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
