城市景觀紅竹林無性系分株生長特征
2010-06-04 09:44:06虞敏之陳雙林鄭連喜郭子武黃樹榮邵宏軍汪忠華
浙江林業科技 2010年2期
虞敏之,陳雙林,鄭連喜,郭子武,黃樹榮,邵宏軍,汪忠華
(1. 浙江省龍游縣林業局,浙江 龍游 324400;2. 中國林業科學研究院亞熱帶林業研究所,浙江 富陽 311400)
紅竹(Phyllostachys iridenscens)自然分布于浙江、江蘇、安徽等地,具有生態適應性強,成林速度快,稈形高大(稈高8 ~ 12m,徑可達10 cm),竹筍產量高,品質佳,竹材劈篾性好,經曬不裂等特點,是筍、材皆可利用的優良經濟竹種[1]。可實施林地有機材料覆蓋措施,促進筍芽提早萌發,顯著提高竹林經濟產出[2]。在我國一些主要竹產區有規模化栽培,已成為區域竹產業發展的重要資源。龍游縣屬剛竹屬竹種中心分布區,紅竹是20世紀90年代逐漸發展起來的主栽筍用竹種,全縣有紅竹林面積0.7萬余hm2,已成為區域農村社會經濟發展的重要資源和農民家庭經濟收入的主要來源,也是龍游縣建立地方特色竹產業的重要組成部分,并在城鎮綠化中得到了較大規模的應用。
紅竹形態優美,枝葉茂盛,竹葉碧綠,籜鞘紫紅色,籜葉為顏色鮮艷的彩帶狀,邊緣桔黃色,中部綠、紫色,也是優良的觀賞竹種。紅竹作為一種重要的生態經濟竹種,開展的相關研究較多,涉及生長發育[3]、生產力評價[4]、林地養分補充[5~6]和有機材料覆蓋[2]、新造林地上部分林分結構變化[7]和地下鞭系結構變化[8]、葉片葉綠素含量[9]等。但就城市景觀綠化紅竹林的相關研究尚無涉及。本文對立地條件、初植密度、栽培管理措施相同的,大密度全梢竹一次成景造林的2種造林年份的城市景觀紅竹林進行了連續3年的新竹數、新竹胸徑和枝下高、全高等因子的調查,以分析紅竹林立竹生長特征,旨在為城市景觀紅竹林良好更新生長、景觀維護提供參考。
1 材料與方法
1.1 試驗地概況
試驗地位于浙江省龍游縣城,屬中亞熱帶氣候,四季分明,年均氣溫17.3℃,極端高溫41℃,極端低溫-11.4℃,年均無霜期261.5d,年均降水量1621.9 mm,年均蒸發量1 392 mm,年平均相對濕度80%。地勢平緩,土壤為紅壤,土層厚度100 cm以上,速效N、P、K含量中等。
1.2 試驗竹林
試驗紅竹林位于龍游縣城江濱公園,分別于2003年、2005年11月營造,面積分別為0.3、0.2 hm2。為達到一次成景效果,采用大密度全梢竹造林方法,初植密度2 ~ 3株/m2。母竹采自未進行過林地有機材料覆蓋的紅竹林,選擇1年生生長健壯,胸徑2 cm左右,無病蟲為害的立竹,帶30 cm以上土球,有來鞭、去鞭。母竹不去梢,栽植前剪去母竹2/3枝葉。栽植后1周內,晴天每天澆水補充土壤水分。試驗林自然生長,不實施土壤養分補充和林地墾復,不采筍、伐竹,僅在成林前(栽植后2 a)每年8月進行1次人工除草。栽植后第3年試驗林立竹郁閉成密集狀,不包括母竹的立竹密度達30 000株/hm2以上。
1.3 調查方法
2009年12月在2種造林年份的試驗紅竹林中分別隨機設置3個5 m×5 m樣地,調查樣地2007-2009年出筍成竹的新竹數、新竹胸徑和枝下高、全高。年度新竹數按照立竹稈色來分辨調查,立竹胸徑用Guanglu電子數顯游標卡尺(分辯率0.01 mm)測量,立竹枝下高用鋼卷尺測量,立竹全高用測高儀測量,精度均為0.01 m。
1.4 數據處理
試驗數據在Excel 2003統計軟件中進行整理和作圖,在DPS.v2.0統計軟件中進行LSD法多重比較,分析不同造林年份不同年度新竹數、新竹胸徑、枝下高、全高的差異性。為避免立竹胸徑差異對立竹枝下高、全高的比較影響,以立竹胸徑為自變量,立竹枝下高、全高為因變量,建立2種造林年份各個年度的新竹枝下高、全高與新竹胸徑的線性回歸關系方程,再分別將6個樣地各個年度的新竹平均胸徑代入方程,對所得的數據組進行差異性分析。
2 結果與分析
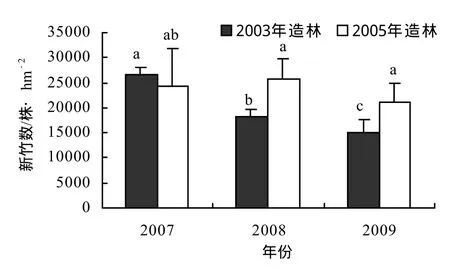
圖1 城市景觀紅竹林年度新竹數變化Figure 1 New bamboo amount of different year in Ph. iridescens plantation for urban landscape
2.1 年度新竹數
由圖1可知,2005年冬季造林的試驗林2007-2009年林分均有大量出筍成竹,雖然 2009年新竹數較前 2年有12.4% ~ 18.1%幅度的下降,但年度間新竹數并無顯著差異。2003年冬季造林的試驗林 2008-2009年新竹數無顯著差異,但均較2007年新竹數顯著減少,降幅達31%以上,并呈隨造林年限的延長,林分新竹數逐年下降的趨勢。比較2個造林年份試驗林,2003年造林林分2007年新竹數與2005年造林林分2007-2009年新竹數差異不顯著,即造林后出筍成竹的前3年林分出筍成竹能力無顯著差異,2008年新竹數顯著下降,即造林后出筍成竹的第4年是林分新竹數顯著變化的轉折點。可以認為,高密度造林的紅竹景觀林從造林后第5年開始,林分出筍成竹能力顯著降低,這與林分密度逐年大幅度提高,地下鞭系延伸生長空間受到限制,林內光照不足,無性系分株間競爭加劇,林分立竹年齡結構不合理,逐漸趨于老化狀態等有關,影響了竹林自我更新和景觀效果維持。2005年龍游縣夏秋季出現嚴重干旱,影響了2006年的出筍成竹,但2007年仍有較大的新竹數,并且較距離造林年份相同年限的2005年造林林分2009年新竹數大,這與竹林林分密度相對較小,林內環境較好,鞭系生長空間較大,養分供應相對充足,營養退筍減少等有關,也說明紅竹生態適應性強,環境資源利用效率高。
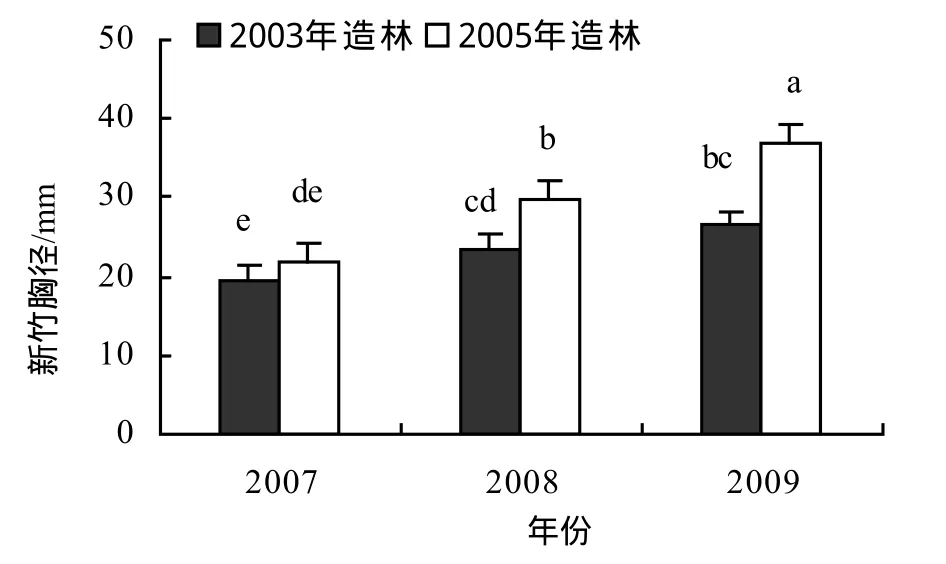
圖2 城市景觀紅竹林年度胸徑變化Figure 2 DBH of new bamboos of different year in Ph. iridescensplantation for urban landscape
2.2 立竹胸徑
由圖2可知,2003年、2005年造林的試驗林年度新竹胸徑均有逐年增大的趨勢,造林后第6年高密度一次成景造林的景觀紅竹林雖然新竹數減少,但新竹胸徑仍在增粗。2005年造林試驗林2007-2009年新竹胸徑均有顯著差異,2008年新竹胸徑比2007年新竹胸徑增大36.2%,2009年又比2008年增大23.4%。2003年造林試驗林2008-2009年新竹胸徑顯著大于2007年,2008-2009年間差異縮小,2008年新竹胸徑比2007年新竹胸徑增大21.7%,2009年比2008年增大13.7%,說明隨著造林年限的延長,新竹胸徑年際間增大的幅度逐漸減小,并趨于相對穩定。造林后第1個出筍成竹年新竹胸徑較母竹小,但第2個出筍成竹年新竹胸徑即可達到母竹胸徑。2003年造林林分2007年新竹與2005年造林林分2009年新竹距離造林年份的年限相同,但新竹胸徑差異極顯著(p < 0.01),這與2005年夏秋季嚴重干旱,影響了2006年的成竹質量有關。試驗林年度新竹最大平均胸徑為36.84 mm,個體最大胸徑49.51 mm,表明如進行適當的林分結構調控,控制林分密度,個體徑級仍有較大的增長空間。
2.3 立竹枝下高
由表1可知,2003年造林試驗林2007年、2009年的新竹枝下高差異顯著,其他年度間新竹枝下高無顯著差異。2005年造林試驗林2007-2009年新竹枝下高均有顯著差異。比較2種造林年份試驗林,2005年造林試驗林2009年新竹枝下高與2003年造林試驗林3個年度的新竹枝下高均差異顯著,2008年新竹枝下高與2003年造林試驗林2009年新竹枝下高無顯著差異,與2007-2008年新竹枝下高差異顯著,2007年新竹枝下高與2003年造林試驗林3個年度的新竹枝下高均無顯著差異。已有研究表明,立竹枝下高是立竹胸徑的從屬因子[10],而2種造林年份試驗林不同年度間新竹枝下高、新竹胸徑差異性不一致,除立竹胸徑對立竹枝下高的決定性影響外,是否不同年度差異顯著的林分密度對新竹胸徑也可能產生一定的影響呢?
為避免立竹胸徑差異對立竹枝下高的比較影響,建立了 2種造林年份不同年度的新竹胸徑(D)與枝下高(h)的關系方程:

式1 ~ 3、式4 ~ 6分別是2003年、2005年造林試驗林2007-2009年新竹胸徑與枝下高的關系方程。將6個樣地 2007-2009年新竹平均胸徑分別代入方程式,得到相同胸徑條件下的不同年度新竹枝下高數據組。經LSD法方差分析,在相同的立竹胸徑條件下,2種造林年份不同年度的立竹枝下高并無顯著差異(表2),可見,在試驗林林分密度范圍內,立竹枝下高是由紅竹固有的生物學和生態學特征所決定的。

表1 城市景觀紅竹林立竹枝下高、全高變化Table 1 Total height and clear bole height of Ph. iridescens plantation for urban landscape

表2 城市景觀紅竹林相同胸徑條件下立竹枝下高、全高變化Table 2 Variation on total height and clear bole height of Ph. iridescens with the same DBH
2.4 立竹全高
2種造林年份試驗林3個年度的新竹全高差異性與新竹枝下高總體上相同,主要差別在于2003年造林試驗林2007年新竹全高與2008-2009年新竹全高差異顯著,與各年度的立竹胸徑差異性一致,說明立竹全高是立竹胸徑的從屬因子。與立竹枝下高相同的方法建立立竹胸徑與立竹全高的關系方程,得出試驗林不同年度相同立竹胸徑條件下的立竹全高(表2),從中可知,2種造林年份試驗林不同年度相同胸徑立竹的全高總體上無顯著差異,其中,2005年造林試驗林2007年、2009年新竹全高差異顯著,這可能與林分密度和竹林養分供應狀況有關。

式7 ~ 9、式10 ~ 12分別是2003年、2005年營造的試驗林2007-2009年新竹胸徑(D)與全高(H)的關系方程。
3 結論與討論
紅竹與散生類竹種和一些克隆植物一樣,其盤根錯節的地下鞭系具有強烈的生理整合能力[11~14],營養物質通過分株間的轉移使資源利用有效性提高,系統收益遠超過耗費[15],并且能集團抵御不良環境。但是這種生理整合效果受制于系統結構狀況,如無性系分株的繁殖力會隨著密度的增加而降低[16]。本研究表明高密度造林的景觀紅竹林造林后前3個出筍成竹年新竹數無顯著差異,而第4個出筍成竹年新竹數則顯著下降,反映出林分密度對分株繁殖能力的影響,也說明為了促進景觀紅竹林的良好更新和景觀效果維持,需要實施人工干擾措施,如疏筍、選擇性伐竹等,如仍沿襲目前的自然生長狀況,到一定年限后,竹林自我更新能力會進一步地急劇下降,甚至不出筍成竹,使竹林趨于老化和退化。紅竹是中大型竹種,筍用林經營的立竹胸徑一般為4 ~ 5 cm,但作為景觀林,需要有較大的立竹胸徑,才能充分體現出觀稈、觀筍的美學價值。本試驗表明,景觀紅竹林造林后前3個出筍成竹年新竹胸徑逐年顯著增大,從第4個出筍成竹年開始,新竹胸徑趨于相對穩定,試驗林最大立竹胸徑不超過5 cm,這也是受林分密度的影響,高林分密度使無性系分株間資源競爭激烈,地下鞭系向上層土壤分布,林內光照減弱,光合作用能力下降,并且竹林易滋生病蟲害,這也說明景觀紅竹林實施林分結構調控的必要性。對此,需進一步開展包括立竹密度、立竹胸徑和年齡結構等的景觀紅竹林合理的林分結構建立研究。
植物具有表型可塑性,器官在復雜環境中會產生一系列不同的相對適合的表現型潛能[17],表型可塑性是植物種群適應環境異質性的重要途徑[18],植物在高林分密度條件下,一般具有促進高生長,抑制徑級生長的趨勢[19~20],較大密度的林分,在林分生長過程中會出現自然稀疏現象[21]。對于竹子而言,立竹枝下高、全高是反映表型可塑性的重要因子,竹林自然稀疏主要表現在出筍減少或退筍增多、新竹抽枝長葉不完全、弱小竹枯死、立竹中下部位枝葉自然整枝等。本試驗表明2種不同造林年份試驗林不同年度相同胸徑的立竹枝下高、全高總體上無顯著差異,反映出立竹枝下高、全高是立竹胸徑的從屬因子,而且除新竹數在造林后一定年限顯著減少外,并沒有出現其他的林分自然稀疏現象,是否試驗林林分密度仍未達到發生立竹表型變化和自然稀疏的密度要求,需進一步開展研究。
[1] 朱石麟,馬乃訓,傅懋毅,等. 中國竹類植物圖志[M]. 北京:中國林業出版社,1994. 128-129.
[2] 陳雙林,吳柏林,張德明. 林地覆蓋對紅竹林溫度和出筍時間分配的影響研究[J]. 林業科學研究,2004,17(專刊):90-94.
[3] 馬乃訓,黃樹田,張文燕. 紅竹生長發育的研究[J]. 林業科學研究,1990,3(4):358-362.
[4] 陳雙林,董建新. 優良經濟竹種紅竹生產力評價[J]. 林業科技開發,2001,15(4):11-16.
[5] 蔣式洪,鐘傳聲. 磷鉀硼肥對紅竹筍品質效應的研究[J]. 竹子研究匯刊,2000,19(1):48-51.
[6] 吳明,吳柏林,曹永慧,等. 不同施肥處理對筍用紅竹林土壤特性的影響[J]. 林業科學研究,2006,19(3):353-357.
[7] 陳雙林,洪游游,張德明,等. 退化紅壤區筍用小徑竹幼林結構變化規律研究[J]. 浙江林學院學報,2005,22(3):296-299.
[8] 洪游游,陳雙林,吳柏林. 新造紅竹林地下結構年際變化規律初步研究[J]. 浙江林業科技,2005,25(4):24-26.
[9] 李建華,楊清平,郭子武,等. 紅哺雞竹葉片葉綠素含量的影響因子初步研究[J]. 浙江林業科技,2009,29(4):65-68.
[10] 陳雙林,吳柏林,虞敏之,等. 毛竹材積主要構件因子關系研究及材積表編制[J]. 福建林業科技,2008,35(2):30-33.
[11] 李睿,鐘章成,M J A 維爾格. 毛竹的無性系生長與立竹密度和葉齡結構的關系[J]. 植物生態學報,1997,21(6):545-550.
[12] Marshall C. Source-sink relations of interconnected ramets[A]. Van Groenendael J,De Kroom H. Clonal growth in plants. regulation and function[M]. The Hague: SPB Acad Publ,1990. 23-41.
[13] 王昱生. 關于無性系植物種群整合作用研究的現狀及其應用前景[J]. 生態學雜志,1994,13(2):57-50.
[14] 董文淵. 竹類無性系種群生態學研究現狀及其應用前景[J]. 林業科學研究,2002,15(2):235-241.
[15] Stuefer J F,During H J,De Kroom H. High benefits of clonal integration in two stoloniferous species in response to heterogeneous light environments[J]. J Ecol,1994(82):511-518.
[16] De Kroom H,Kwant R. Density-dependent growth responses in two clonal herbs: regulation of shoot density[J]. Oecologia,1991(86):298-304.
[17] Dewitt T J. Costs and limits of pheno typic plasticity[J]. Trends Ecol Evol,1998,13(2):77-81.
[18] Hutchings M J,Price E, Sih A, et al. Dose physiological integration enable clonal herbs to integrate the effects of environmental hetero geneity?[J]. PlantSpec Biol,1993(8):95-105.
[19] 黃寶靈,呂成群,蒙鈺釵. 尾葉桉人工林種群密度的研究[J]. 應用生態學報,2000,11(1):30-32.
[20] 樸順姬,楊持,黃紹峰,等. 羊草種群密度與生長動態研究[J]. 植物生態學報,1997,21(1):60-66.
[21] 鄭海水,曾杰,翁啟杰,等. Nelder試驗:大葉相思樹生長與密度、年齡的相關研究[J]. 林業科學研究,1996,9(2):158-164.
