堿茅耐鹽堿基因克隆研究進展
2010-06-08 07:52:34任偉王志峰徐安凱
草業學報 2010年5期
任偉,王志峰,徐安凱
(吉林省農業科學院畜牧科學分院,吉林 公主嶺136100)
堿茅(Puccinellia)是生長在草甸草原、鹽漬化土壤堿斑周圍的冷季型多年生禾本科牧草。它的幼苗,在p H值大于10、表土含鹽量5%以上的土壤中,仍能正常生長,被譽為鹽堿地的先鋒植物[1]。作為一種抗鹽堿、耐低溫、飼用價值好的優良牧草,引起了諸多學者的重視。以往的科技工作者對堿茅在鹽堿脅迫下的生理生化反應[2-6]、種子萌發[7-9]、形態解剖[10-12]、耐鹽機制[13-17]、遺傳育種及鹽堿地生態恢復與改良[18-20]等方面做了詳細的研究。近年來,隨著分子生物學的飛速發展,許多新的技術如分子克隆、分子標記、轉基因、差異顯示(DDRT)、擬制性消減雜交(SSH)、c DNA微陣列技術(c DNA microarray)、表達序列標簽(EST)等廣泛應用于堿茅耐鹽機理的研究,取得許多重要的研究成果。獲得了大量的鹽堿誘導基因,主要集中在鹽堿脅迫條件下,與其滲透調節物質合成、抗氧化保護酶、離子通道蛋白、信號轉導蛋白作用有關的基因。對堿茅耐鹽堿基因的挖掘,不僅有助于更好的研究堿茅自身的耐鹽堿機制,而且可以加快牧草及其他農作物的品質改良,具有重要的理論意義和很高的實際應用價值。
1 滲透調節物質合成基因
當植物遭遇干旱、鹽堿、低溫等逆境時,就會在體內積累各種小分子物質,提高細胞液濃度,降低其滲透勢,并保持一定的壓力勢,來維持細胞的正常膨壓。參與滲透調節的物質有很多,大致可分為兩大類。一類是由外界進入細胞的無機離子,一類是在細胞內合成的有機物質,如脯氨酸(proline)、甜菜堿(betaine)、芒柄醇(D-ononitol)、甘露醇(mannitol)、山梨醇(sorbitol)和海藻糖(trehalose)等。
目前,從植物中克隆的參與滲透調節物質合成的基因主要是BADH基因(甜菜堿醛脫氫酶基因,betaine aldehyde dehydragenase)、P5CS基因(Δ’-吡咯啉-5-羧酸合成酶基因,Δ’-pyrroline-5-carboxylate synthase)[21]和Imtl基因(肌醇甲基轉移酶基因,inositol O-methyltrans ferase)[22]。其中BADH基因是目前耐鹽、耐旱基因工程中研究得較深入的基因之一,是公認的截至目前為止,最具抗鹽堿效果的基因。楊錚等[23]和鐘鳴等[24]通過同源序列克隆結合cDNA末端快速擴增技術(rapid amplification of cDNA ends,RACE),在朝鮮堿茅中擴增出BADH基因,共1 502 bp。序列比對后發現,朝鮮堿茅與大麥(Hordeumbrevisubulatum)、羊草(Leymuschinensis)的同源性高達89%,含有醛脫氫酶特有的十肽保守序列(VSLELGGKSP),但是與多數植物中BADH的十肽保守序列(VTLELGGKSP)存在細微的差別,朝鮮堿茅第2位的T被S取代。其后29位點上含有與酶功能有關的半胱氨酸殘基,可能包含NAD+結合位點及酶催化位點,這些微小的變化及內含的結合位點可能與增強堿茅的耐鹽能力有關。C末端的SKL信號肽序列,說明甜菜堿醛脫氫酶定位于朝鮮堿茅體內的過氧化物酶體中,而甜菜堿的合成是在葉綠體中完成,朝鮮堿茅是如何完成這個運輸過程的,會不會影響它的耐鹽能力,以及這個BADH基因的耐鹽生理特性如何,都值得做進一步的研究。
2 抗氧化脅迫基因
鹽脅迫下,細胞內會產生氧自由基(O2-)、過氧化氫(H2O2)和羥基自由基(OH-)等活性氧物質(ROS),引起膜脂過氧化,蛋白質變性,核酸降解,進而導致氧化脅迫的產生。植物有2種防止活性氧危害的系統:酶促系統和非酶促系統。非酶促系統主要包括一些直接參與活性氧清除的抗氧化物,如抗壞血酸、谷胱甘肽、多元醇、α-生育酚、類胡蘿卜素和黃酮等。酶促系統是指參與保護反應的酶類,主要有超氧化物歧化酶(super oxide dismutase,SOD),過氧化氫酶(catalase,CAT),抗壞血酸過氧化物酶(aseorbate peroxidase,APX),谷胱甘肽過氧化酶(glutathione peroxidase,GPX),谷胱甘肽硫轉移酶(glutathione s-transferase,GST),單脫氧抗壞血酸還原酶(monodehydro aseorbate reduetase,MDHAR)及脫氧抗壞血酸還原酶(dehydroascorbic acid reductase,DHAR)等[25]。在堿茅中,目前研究比較多的是酶促系統中的一些關鍵酶類。
2.1 脫氫抗壞血酸還原酶(DHAR)基因和鐵蛋白(Fer)基因
張曉磊[26]根據已有的堿茅EST片段設計基因特異性引物,克隆到了脫氫抗壞血酸還原酶(DHAR)和鐵蛋白(ferritin,Fer)2個抗氧化基因,并研究了不同鹽堿處理條件下這2個基因的表達變化情況。PtDHAR基因,開放閱讀框(ORF)639 bp,編碼213個氨基酸,以谷胱甘肽為底物,催化脫氫抗壞血酸還原為抗壞血酸,在保護細胞組分抵御氧化損傷及循環利用抗壞血酸中起重要作用[27]。鐵蛋白(PtFer)基因開放閱讀框為336 bp,編碼111個氨基酸。在高氧環境下,氧與鐵反應會產生氧自由基,對植物產生巨大的毒性而損害生物有機體[28]。而鐵蛋白具有很強的貯鐵功能,可以將鐵以無毒的形式貯存起來,為依賴于鐵的生理生化過程起著暫時性緩沖作用,減少植物體內各種氧化脅迫產生的傷害作用,增強植株對逆境脅迫的抗性,提高植物的生長速度[29]。Northem雜交的結果表明,鹽堿脅迫后,這2個基因都有不同程度的表達,但是隨著處理時間的延長,DHAR的表達量先增加后減少,而鐵蛋白的表達量持續增加。隨著處理濃度的遞增,DHAR的表達量變化不大,而鐵蛋白的表達量在200 mmol/L處理后達到最大,并且堿茅地上部與根部對脅迫響應也存在時空上的差異[26]。說明同一植物不同基因以及同一基因在堿茅不同部位對鹽堿脅迫的響應機制是不同的,它們之間可能相輔相成,也可能相互制約。今后,應該注重不同耐鹽堿基因之間互作效應的研究。
2.2 抗壞血酸過氧化物酶(APX)基因
作為抗氧化系統中清除鹽堿脅迫產生的H2O2的主要酶類——抗壞血酸過氧化物酶(ascorbate peroxidase,APX)[30],它催化H2O2還原為H2O的反應,對抗壞血酸具有很高的特異親和性[31]。它的基因在堿茅中也得到分離克隆[32]。序列分析表明,開放讀碼框(ORF)為876 bp,編碼291個氨基酸。它是細胞質過氧化物體的分泌蛋白,在細胞質中合成,最后通過蛋白轉運系統定位于過氧化物體。過量表達PutAPX基因的酵母在含H2O2的SD培養基上生長良好,對組織和細胞起到了保護作用,體現了抗壞血酸過氧化物酶在氧化脅迫下的作用。為進一步研究APX在堿茅體內的轉運機制以及逆境誘導下氧化脅迫的作用機理奠定了基礎。
2.3 超氧化物歧化酶(SOD)基因
超氧化物歧化酶(super oxide dismutase,SOD)是一種含金屬的抗氧化酶。對于清除氧自由基,防止氧自由基破壞細胞的組成、結構和功能,保護細胞免受氧化損傷具有十分重要的作用[33],與植物的抗旱、抗鹽堿、耐高溫等多種逆境有密切關系[34],主要可分為3類:Cu/Zn-SOD、Mn-SOD和Fe-SOD[35]。吳建慧等[36]從堿茅根c DNA文庫中分離得到Put-Cu/Zn-SOD基因,全長cDNA,開放讀碼框長615 bp,所編碼的蛋白由204個氨基酸組成。對轉Put-Cu/Zn-SOD酵母進行鹽堿、氧化脅迫的實驗結果表明,堿茅Put-Cu/Zn-SOD基因對Na2CO3、Na HCO3和H2O2有著良好的抗性,顯著提高了酵母的抗鹽堿和耐氧化能力。也有一些研究表明,單個抗氧化脅迫基因的能力往往是有限的,所以要從堿茅中克隆更多的抗氧化脅迫基因,研究多個基因在增強植物抗逆能力中的作用機理及應用。
3 離子脅迫相關基因
植物消除離子脅迫主要有3種機制,即降低離子吸收、離子外排和離子區隔化[37,38],這3種機制的關鍵在于一些離子通道蛋白和逆向轉運蛋白對離子的選擇性吸收或轉運作用。目前,在堿茅中克隆到的編碼此類蛋白的功能基因主要與 HKT(high-affinity K+transporter)轉運蛋白、NHX(Na+/H+exchanger)逆向轉運蛋白、CAX(Ca2+/H+exchanger)反向轉運蛋白和質子泵蛋白的作用有關。
3.1 Na+/H+逆向轉運蛋白基因(NHX)
植物Na+/H+逆向轉運蛋白(Na+/H+exchanger,NHX)分為質膜型和液泡膜型2類,依靠H+-ATPase和H+-PPase產生的質子驅動力介導Na+/H+跨膜運輸,分別負責細胞質內的Na+外排和Na+區隔化,可以調節植物的耐鹽能力[39]。程玉祥[40]分離了堿茅質膜型Na+/H+逆向轉運蛋白基因(PtSOS1),Southern分析表明PtSOS1是單拷貝基因。半定量RT-PCR結果顯示,PtSOS1受鹽脅迫上調表達。說明PtSOS1可能在星星草(Puccinelliatenuiflora)的抗鹽堿能力中起作用。也有研究表明PtSOS1在擬南芥(Arabidopsisthaliana)中的過表達,提高了轉基因植株的耐鹽性[41]。但是堿茅質膜型Na+/H+逆向轉運蛋白是否參與了體內K+的運輸,與水稻(Oryzasativa)、擬南芥等植物的運轉功能是否相同,還不清楚。可以采用過表達PtSOS1和反義RNA擬制表達的方法來研究它的結構與功能。
3.2 Ca2+/H+反向轉運蛋白基因(CAX)
Ca2+/H+反向轉運體(Ca2+/H+exchanger,CAX)與Ca2+-ATP酶同是Ca2+外向轉運器,負責將胞質中的Ca2+運出細胞,或運入細胞內的液泡及其他細胞器[42]。CAX是 Ca2+/cation反向轉運體(Ca2+/cation antiporter,CaCA)的家族之一,在植物、真菌、細菌和低等脊椎動物中均有存在。它是不直接需求三磷酸腺苷(adenosinetriphosphate,ATP)的次級轉運器,利用質子驅動力(pmf)來驅動鈣離子的運輸[43],對植物體內的離子平衡有著極其重要的作用。Liu等[44]在堿茅中分離到的PutCAX基因,具有提高酵母對Ca2+或Ba2+抗性的功能,并且首次提出CAX基因具有逆向轉運Ba2+的功能。通過GFP熒光蛋白標記和FM4-64(分子探針)染色,初步證明PutCAX定位于釀酒酵母的液泡膜上。通過觀察PutCAX基因N、C端缺失酵母轉化體的生長情況,發現N端和C端對PutCAX運輸Ca2+或Ba2+具有一定的調控作用。但是,在轉化試驗中發現,一些轉PutCAX擬南芥植株的生長反而受到擬制,是不是由于PutCAX的過表達,打破了擬南芥體內原有的離子平衡穩態,還是與其他重金屬離子產生了離子拮抗作用,值得做進一步詳細的研究。
3.3 Na+/K+轉運蛋白基因(HKT)
植物HKT家族轉運蛋白(high-affinity K+transporter)的主要功能是參與Na+/K+在植物體內的選擇性轉運,已有的研究結果表明K+/Na+在細胞內的選擇性轉運,是堿茅體內一個非常重要的耐鹽機制[17],說明堿茅HKT基因在耐鹽轉基因工程中可能具有重大應用價值。植物HKT蛋白的一些跨膜區和螺旋區的氨基酸組成是非常保守的[45],這為利用同源克隆方法分離植物的HKT基因提供了方便。但是HKT基因在總體上的保守性比較差,即使單個氨基酸殘基的變化也會顯著改變選擇性轉運Na+的活性[46]。因此Ardie等[47]將從堿茅中分離的PutHKT2與水稻中克隆的OsHKT2同時轉化酵母和擬南芥,來研究2個HKT基因的表達差異性。結果表明,轉堿茅PutHKT2基因的酵母表現出高親和性的K+-Na+共轉運系統。轉PutHKT2擬南芥可以抵抗外界Na+、K+和Li+多種離子的脅迫,而轉OsHKT2擬南芥僅僅對外界Na+表現敏感。這也許是堿茅比水稻更耐鹽堿的原因之一。此外,同一蛋白家族的基因,表達效果也不盡相同。Zhang等[48]對堿茅質膜家族蛋白3(plasma membrane protein 3)的2個基因PutPMP3-1、PutPMP3-2的研究發現,后者的抗逆能力更強。
4 參與脅迫信號轉導途徑的基因
干旱、鹽堿、低溫等外界逆境因子實質上是一種體外信號,當植物感知體外信號后,可以引發一系列的體內信號,進而誘導相關基因表達調控的改變。在長期進化過程中,植物擁有完整的信號網絡用以調節各種環境脅迫引起的響應。隨著研究的逐步深入,脫落酸(ABA)被普遍認為參與了在鹽脅迫下的滲透脅迫信號轉導。相應地,調控ABA信號響應基因的表達也被證實有助于提高植物的耐鹽性[49]。ABA可以引起氣孔的關閉,維持植物體內水分平衡,保護質膜結構和功能,提高植物的抗鹽能力[50]。于雪飛和楊傳平[51]在堿茅中分離到受ABA和高鹽誘導的Put-R40g3基因,全長588 bp,編碼195個氨基酸,與水稻,小麥和擬南芥的氨基酸序列有較高的同源性,綠色熒光蛋白(GFP)定位研究結果顯示Put-R40g3蛋白存在于酵母細胞質中。顯著提高了酵母對鹽、干旱及氧化等逆境條件的適應能力。
除了ABA信號轉導之外,植物還有很多響應鹽脅迫的信號途徑。已經證實鹽過敏感(salt overly sensitive,SOS)信號途徑在植物耐鹽中起調控作用,在這條途徑中,鈣離子作為第2信使與鈣離子結合蛋白結合并激活下游一系列蛋白,進而調控植物的耐鹽性[52,53]。此外,蛋白質磷酸化和去磷酸化也是調控細胞響應各種外源信號的重要機制,MAP激酶(促分裂原活化蛋白激酶)級聯途徑在內源、外源信號傳導過程中均發揮著重要的作用。MAP激酶級聯途徑包括3種蛋白激酶:MAPKK激酶(MAPKKK)、MAPK激酶(MAPKK)和 MAP激酶(MAPK),構成三級激酶模式[54-56]。
大量的證據表明,這些脅迫信號轉導途徑間存在聯系和交叉作用。鹽脅迫的早期信號轉導事件以及早期誘導表達的基因是一個關鍵所在,分離堿茅體內此類脅迫響應基因是今后植物耐鹽性研究的一個重點方向。在鹽堿脅迫下,堿茅體內相應的細胞受體是什么?從堿茅感知脅迫信號到合成相關基因的表達蛋白,具體的信號轉導過程是什么?這些問題的解決將有助于加深對堿茅耐鹽堿機理的研究。
5 基因組學在堿茅耐鹽堿機理研究中的應用
隨著植物基因組學與生物信息學的飛速發展,為堿茅耐鹽堿機理的研究及相關基因的克隆提供了許多新的技術,如:差異顯示(DDRT)、擬制性消減雜交(SSH)、c DNA微陣列技術(c DNA microarray)和表達序列標簽(EST)等。已有許多學者應用其中一種,或是幾種技術相結合的方法,對不同濃度、不同時間鹽堿脅迫處理下,堿茅相關基因的表達情況,作了詳細的研究[57-60]。其中已知的同源EST序列,大多數是與活性氧清除、滲透調節、基因調控、離子轉運過程有關的基因片段。DNA芯片(DNA chip)的研究結果顯示,一些基因在鹽脅迫下表達下調,另一些基因在鹽脅迫下表達上調,這些基因的功能主要涉及了信號傳導、轉錄調控、細胞防御、細胞代謝等方面,這些差異表達的基因可能在星星草抗鹽堿過程中具有關鍵作用。植物的抗鹽堿過程是一個復雜的多因子作用體系[61],通過新興的基因組學研究手段,可以有效獲取鹽堿脅迫下堿茅基因表達的大量特征信息,從而為系統闡明堿茅抗鹽堿分子機理奠定堅實的基礎,也為尋找未知的抗逆基因提供了新途徑。
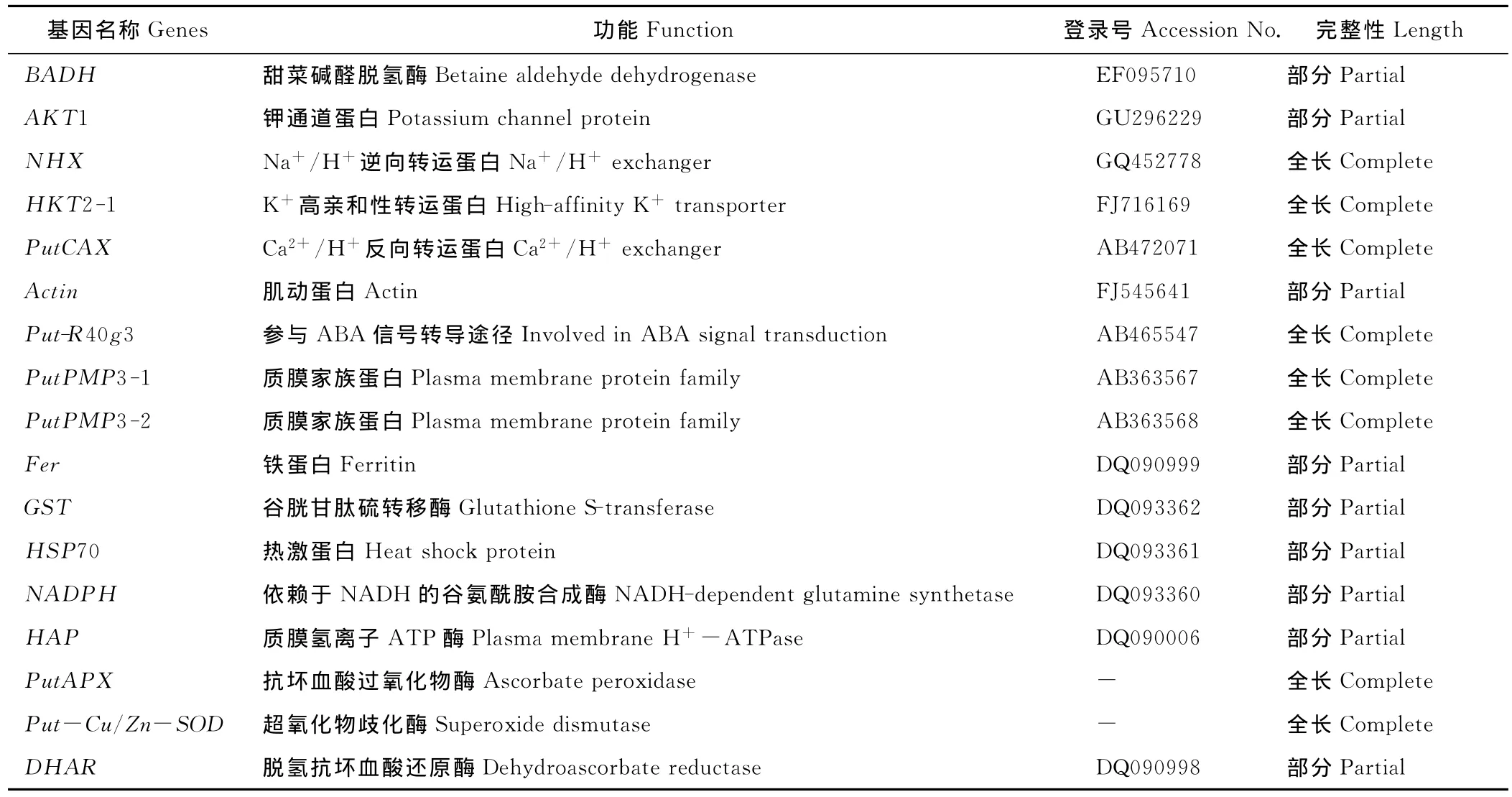
表1 NCBI基因庫中已登錄的堿茅鹽堿誘導基因或片段Table 1 The salt and alkali inducible gene or cDNA segment registered in Gene bank
6 問題與展望
目前,在美國國立生物技術信息中心(NCBI)基因庫中登錄的與堿茅鹽堿誘導相關的基因或片段總共有17種(表1)。不得不承認我國牧草分子生物學的研究要遠遠落后于水稻、小麥等農作物的研究[62-64]。但是,農業生產面臨的首要逆境脅迫就是土壤鹽漬化,而培育耐鹽堿轉基因農作物有望解決這一世界難題。分子選育抗鹽堿植物新品種的關鍵,就是從鹽生植物里分離抗鹽堿功能基因,堿茅,作為一種寶貴的鹽生種質資源、為相關研究提供了豐富的耐鹽堿基因。加大對堿茅分子生物學的研究,不僅有助于培育耐鹽堿轉基因農作物、牧草和花卉,而且可以為我國鹽堿草地的改良和治理提供理論基礎,具有重大的經濟價值和生態效應。
[1]中國飼用植物志編輯委員會.中國飼用植物志(第一卷)[M].北京:農業出版社,1978:21-22.
[2]王蘋,李建東,歐勇玲.松嫩平原鹽堿化草地星星草的適應性及耐鹽生理特性的研究[J].草地學報,1997,5(2):80-84.
[3]石德成,殷麗娟.鹽與堿對星星草脅迫作用的差異[J].植物學報,1993,35(2):144-149.
[4]王鎖民,朱興運,趙銀.鹽脅迫對拔節期堿茅游離氨基酸成分及脯氨酸等含量的影響[J].草業學報,1994,3(3):22-26.
[5]王鎖民,朱興運,王增榮.滲透調節在堿茅(Puccinelliatenuiflora)幼苗適應鹽逆境中的作用初探[J].草業學報,1993,2(3):40-46.
[6]劉延吉,張蕾,田曉艷.鹽脅迫對堿茅幼苗葉片內源激素、NAD激酶及Ca2+-ATPase的效應[J].草業科學,2008,25(4):51-54.
[7]徐安凱.堿茅發芽生理及恒溫發芽選育效果研究[D].蘭州:甘肅農業大學,1995.
[8]李艷波,陳月艷,孫國榮,等.鹽堿脅迫下星星草種子萌發過程中氮代謝的初步研究[J].植物研究,1999,19(2):153-158.
[9]陳月艷,孫國榮,李景信.Na2CO3脅迫對星星草種子萌發過程中水分吸收及膜透性的影響[J].草業科學,1997,14(2):27-30.
[10]朱宇旌,張勇,胡自治,等.小花堿茅根適應鹽脅迫的顯微結構研究[J].中國草地,2001,23(1):37-40.
[11]韋存虛,王建軍,王建波.Na2CO3脅迫對星星草葉肉細胞超微結構的影響[J].生態學報,2006,26(1):108-114.
[12]楊春雪,申家恒.星星草受精作用及其胚與胚乳早期發育的觀察[J].武漢植物學研究,2004,22(2):91-97.
[13]閻順國,朱興運,郭樹林.堿茅草對土壤鹽分動態及鹽量平衡的影響[J].水土保持學報,1990,4(1):44-48.
[14]朱興運,王鎖民,閻順國.堿茅屬植物抗鹽性與抗鹽機制的研究進展[J].草業學報,1994,3(3):9-15.
[15]Wang S M,Zhao G Q,Gao Y S.Puccinelliatenuifloraexhibits stronger selectivity for K+over Na+than wheat[J].Journal of Plant Nutrition,2004,27:1841-1857.
[16]Peng Y H,Zhu Y F,Mao Y Q.Alkali grass resists salt stress through high K+and an endodermis barrier to Na+[J].Journal of Experimental Botany,2004,55:939-949.
[17]Wang C M,Zhang J L,Liu X S.Puccinelliatenuifloramaintains a low Na+level under salinity by limiting unidirectional Na+influx resulting in a high selectivity for K+over Na+[J].Plant Cell and Environment,2009,32:486-496.
[18]吳青年,徐安凱.堿茅草改良堿斑草場綜合技術[J].中國農業科學,1987,20(1):92.
[19]徐安凱.“吉農朝鮮堿茅”與“三北”地區鹽堿地的改良[J].牧草與飼料,2007,1(2):3-5.
[20]齊寶林,高國臣,趙云鵬.耐鹽抗寒優質牧草——堿茅[J].吉林林業科技,2005,(9):4-7.
[21]Hong Z.Removal of feedback inhibition ofΔ1-pyrroline-5-carboxylate synthase results in increased proline accumulation and protection of plants from osmotic stress[J].Plant Physiology,2000,122:1129-1136.
[22]Vernon D M,Bohnert H J.A novel methy transferase induced by osmotic stress in the facultative Mesembryanthemum crystallium[J].Embo Journal,1992,11:2077-2085.
[23]楊錚,鐘鳴,郭志富.鹽脅迫下朝鮮堿茅的甜菜堿醛脫氫酶活性變化及其基因保守區的克隆[J].植物生理學通訊,2007,4(3):430-434.
[24]鐘鳴,張佳,郭志富.朝鮮堿茅BADH基因3′端及5′端部分序列的擴增[J].華北農學報,2009,24(3):46-50.
[25]方允中,鄭榮梁.自由基生物學的理論與應用[M].北京:科學出版社,2002:30-31.
[26]張曉磊.星星草PtDHAR及PtFer基因的克隆與表達分析[D].黑龍江:東北林業大學,2008.
[27]Yabuta Y,Motoki T,Yoshimura K.Thylakoid membrane-bound ascorbate peroxidase is a limiting factor of antioxidative systems under photo-oxidative stress[J].The Plant Journal,2002,32:915-925.
[28]Guenriot M L,Yi Y.Iron:Nutritions,noxious and not readily avaliable[J].Plant Physiology,1994,104(33):815-820.
[29]Balla G,Jacob H S,Balla J.Ferritin:A cytoprotective antioxidant strategem of endothelium[J].Journal of Biology Chemistry,1992,267:18148-18153.
[30]Asada K.Ascorbate peroxidase:A hydrogen peroxide scavenging enzyme in plants,Physiologia[J].Plantarum,1992,85(2):235-241.
[31]Shigeoka S,Nakano Y,Kitaoka S.Purification and some properties of L-ascorbic acid peroxidase inEuglenagracilisZ[J].Archives of Biochemistry and Biophysics,1980,201(1):121-127.
[32]管清杰,李琳,高野哲夫.朝鮮堿茅抗壞血酸過氧化物酶的基因克隆和表達[J].基因組學與應用生物學,2009,28(4):631-639.
[33]Song F N,Yang C P,Liu X M.Effect of salt stress on activity of superoxide dismutase(SOD)inUlmuspumilaL[J].Journal of Forestry Research,2006,17(1):13-16.
[34]張海娜,李小娟,李存東.過量表達小麥超氧化物歧化酶(SOD)基因對煙草耐鹽能力的影響[J].作物學報,2008,34(8):1403-1408.
[35]Alscher R G,Erturk N,Heath L S.Role of super oxide dismutases(SODs)in controlling oxidative stress in Plants[J].Journal of Experimental Botany,2002,53:1331-1341.
[36]吳建慧,高野哲夫,柳參奎.堿茅(Puccinelliatenuifolra)Put-Cu/Zn-SOD基因的克隆及在酵母中的表達[J].基因組學與應用生物學,2009,28(1):10-14.
[37]Ward J M,Hirsi K D.Plants pass the salt[J].Trends in Plant Science,2003,8(5):200-201.
[38]Teste R M,Daenport R.Na+tolerance and Na+transport in higher plants[J].Annals of Botany,2003,91(5):503-527.
[39]Blumwald E,Aharon G S,Apse M P.Sodium transport in plant cells[J].Biochimica et Biophysica Acta,2000,1465:145-151.
[40]程玉祥.星星草質膜型Na+/H+逆向轉運蛋白基因的克隆和特性分析[J].植物生理學通訊,2008,44(1):59-64.
[41]程玉祥.過量表達星星草PtSOS1提高擬南芥的耐鹽性[J].植物生理學通訊,2008,44(6):1125-1130.
[42]Kendal D,Hirschi,Victor D.Expression of Arabidopsis CAX2 in tobacco altered metal accumulation and increased manganese tolerance1[J].Plant Physiology,2000,124:125-134.
[43]Takehiro K,Masayoshi M.Residues in internal repeats of the rice cation/H+exchanger are involved in the transport and selection of cations[J].The Journal of Biological Chemistry,2004,279:812-819.
[44]Liu H,Zhang X X,Takan T.Characterization of aPutCAX1 gene fromPuccinelliatenuiflorathat confers Ca2+and Ba2+tolerance in yeast[J].Biochemieal and Biology Physieal Research Communieations,2009,383:392-396.
[45]Arciadeb G,Nme S,Uelosma B.Sodium transport and HKT transporters:The rice model[J].Plant Journal,2003,34(6):788-801.
[46]Aser P M,Sooy H O,Shimas G O.Glycine residues in potassium channel-likes electivity filters determine potassiums electivity in four-loopper-subunit HKT transporters from plants[J].Proceedings of the National Academy of Sciences of the united states of america,2002,99(9):6428-6433.
[47]Ardie S W,Xie L,Takahashi R.Cloning of a high-affinity K+transporter genePutHKT2;1 fromPuccinelliatenuifioraand its functional comparison withOsHKT2;1 from rice in yeast and Arabidopsis[J].Journal of Experimental Botany,2009,12:1-12.
[48]Zhang C Q,Nishiuchi S,Liu S K.Characterization of two plasma membrane protein 3 genes(PutPMP3)from the alkali grass,Puccinelliatenuiflora,and functional comparison of the rice homologues,OsLti6a/b from rice[J].Biochemistry and Molecular Biology reports,2008,41(6):448-454.
[49]Moons A,Gielen J,Vandekerckhove J.An abscisic-acid and salt stress responsive rice cDNA from a novel plant gene family[J].Planta,1997,202(4):443-454.
[50]Huang M D,Wu W L.Over expression of TMAC2,an ovel negative regulator of abscisic acid and salinity responses,haspleio tropic effects inArabidopsisthaliana[J].Plant Molecular Biology,2007,63(4):557-569.
[51]于雪飛,楊傳平.堿茅(Puccinelliatenuiflora)Put-R40g3基因的分離及其與逆境的應答[J].分子植物育種,2009,7(2):251-256.
[52]Botella J R,Arteca J M,Somodevill A M.Calcium-dependent protein kinase gene expression in response to physical and chemical stimuli in Mungbean(Vignaradiata)[J].Plant Molecular Biology,1996,30:1129-1137.
[53]Zhu J K.Genetic analysis of plant salt tolerance using Arabidopsis[J].Plant Physion,2000,124:941-948.
[54]Jona K C,Okresz L,Bogre L.Complexity,cross talk and integration of plant MAP kinase signaling[J].Current Opinion in Plant Biology,2002,5:415-424.
[55]Zhang S Q,Daniel F.MAPKcascades in plant defense signaling[J].Trends in Plant Science,2001,6(11):520-527.
[56]Hackett R M,Oh S A,Morris P C.A tomato MAP kinase kinase gene differentially regulated during fruit development leaf senscence and wounding[J].Plant Physiology,1998,117:1526-1531.
[57]劉桂豐,褚延廣,王玉成.cDNA微陣列技術研究Na HCO3脅迫下星星草基因表達譜[J].西北植物學報,2005,25(5):887-892.
[58]Wang Y C,Yang C P,Liu G F.Microarray and suppression subtractive hybridization analyses of gene expression inPuccinelliatenuifioraafter exposure to Na HCO3[J].Plant Science,2007,173:309-320.
[59]Wang Y C,Yang C P,Liu G F.Development of a cDNA microarray to identify gene expression ofPuccinelliatenuifioraunder salineealkali stress[J].Plant Physiology and Biochemistry,2007,45:567-576.
[60]Wang Y C,Yang C P,Liu G F.Identification of expressed sequence tags in an alkali grass(Puccinelliatenuifiora)cDNA library[J].Journal of Plant Physiology,2007,164:78-89.
[61]周學麗,周青平,顏紅波,等.NaCl脅迫對同德小花堿茅苗期生理特性的影響[J].草業科學,2009,26(6):101-105.
[62]張一弓,張麗靜,傅華.植物維生素E合成酶基因克隆及其逆境生理研究進展[J].草業學報,2009,18(5):235-242.
[63]梁哲,姜三杰,未麗,等.三葉草基因工程研究進展[J].草業學報,2009,18(2):205-211.
[64]鄭軼琦,劉建秀.草坪草分子遺傳圖譜的構建與應用研究進展[J].草業學報,2009,18(1):155-162.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
