南洞庭湖黃土包河不同年齡洲灘生態特征的初步研究
2010-06-14 08:58:04謝永宏陳心勝
作物研究 2010年4期
潘 瑛,謝永宏,陳心勝,李 峰
(1中國科學院亞熱帶農業生態研究所亞熱帶農業生態過程院重點實驗室,湖南長沙 410125;2中國科學院亞熱帶農業生態研究所洞庭湖濕地生態系統觀測研究站,湖南長沙 410125;3中國科學院研究生院,北京 100039)
濕地普遍存在泥沙淤積現象,主要是由河流的沖積作用造成[1]。一般來說,由于水文地理特征的特殊性及其時空變化的不均勻性,各濕地泥沙淤積存在差異,如美國伊利諾斯州每年僅淤積0.5~ 1.0 cm[2],而淤積活躍地區如美國福爾河甚至達60~120 cm[3]。泥沙淤積使濕地土壤理化性質和土壤-植物微環境發生了一系列的變化,是影響濕地演替方向的重要驅動因素之一[4]。此外,地上植被能改善土壤物理結構,促進環境中營養物質的循環及影響濕地泥沙淤積過程等[5],因此植物因素也是決定濕地發育和演替的重要環境因素[6,7]。近年來,關于泥沙淤積對濕地植被演替的研究逐漸成為國際熱點[8,9],但是濕地演替過程中泥沙淤積、土壤理化性質及植物群落三者間的關系還遠沒有研究清楚,決定濕地演替的關鍵因子尚不明確。本研究選擇洞庭湖為研究對象對以上問題進行探討。
洞庭湖是我國第二大淡水湖泊,具有調節江河、凈化水質、溝通航運、繁衍水產和改善生態環境等重要生態功能[10]。研究表明,1956~ 1995年間,洞庭湖多年平均沉積泥沙量達 1.27億噸,占入湖泥沙量的81.2%[11],其中荊江三口及湘、資、沅、澧四水帶來的泥沙沉積是形成洞庭湖洲灘濕地的主要原因。洞庭湖局部地區泥沙沉積的不均衡性,引起洲灘濕地發育與演替的差異[12]。調查發現,洞庭湖濕地演替依泥沙淤積的速度和方式而異。前人對洞庭湖泥沙淤積與洲灘濕地的形成與發育已有大量的研究[13~15],但是對不同演替階段洲灘生態特征變化規律及其關鍵調控因子的研究尚未見報道。本文以空間代替時間的方法對不同年齡洲灘濕地土壤特征和植被特征的空間分布規律及其相關性進行了研究,以揭示洲灘濕地演替的關鍵環境因子,為洞庭湖濕地的保護和合理利用提供依據。
1 材料和方法
1.1 研究區概況
本研究選擇南洞庭湖黃土包河洲灘濕地為研究對象。黃土包河位于南洞庭湖的北沿,西起沅江縣赤山,東連湘陰縣毫巴 ,研究區域地理坐標:E:112°28′07.4″~ 112°35′54.7″,N:28°54′05.7″~ 28°55′28.3″,全 長58.92 km,是一條重要的泄洪、輸沙河道,承擔著自長江流入的20%~30%的輸沙任務。由于該區高洪時期河湖不分,下游分支頗多,水情復雜,加劇了泥沙淤積程度,故洲灘形成非常活躍,為空間代替時間來研究洲灘濕地演替奠定了基礎。
1.2 研究方法
(1)野外調查取樣
調查于 2009年 4月底進行,此時為洪水來臨前植物最活躍的時期。在試驗地選擇不同形成年份(10,20,30,50 a)的9塊洲灘(洲灘基本狀況見表1,其形成時間通過走訪當地老漁民和行政部門確定)。在每個洲灘挖取 2~ 3個剖面,各剖面均按 0~ 20,20~ 40,40~ 60 cm分層,用封口袋裝好帶回實驗室。每個采樣點設置2~3個 1 m×1 m樣方,對植被特征進行調查。采樣的過程中對每個樣點精確定位。此次取樣共含樣點 25個,樣方總數為 51個。在調查過程中,因當地大面積種植楊樹(外來種)有可能改變洲灘的固有特性,因此以原生特征(周圍無大面積楊樹)為樣點選擇依據。
(2)生物特征
以每平方米地上部分的鮮重作為評價生物量的標準;物種豐富度采用Menhinick豐富度指數(1964):R表示每一個樣方中的物種總數,N為全部種的個體數目;多樣性指數采用 Shannon-Wiener指數:H=-∑ Pi×log2Pi,Pi表示物種重要值=(相對密度+相對高度+相對蓋度)/300。
(3)土壤理化特征測定
土樣帶回實驗室后,除去植物殘體及土壤動物。將土樣分為兩份,一份用于測定土壤還原性物質總量和土壤含水量;一份用于測定土壤顆粒組成、pH、有機質、全氮、全磷、全鉀。土壤機械組成采用吸管法,分為粗 砂 (直 徑大于 0.25 mm)、細 砂 (直徑 0.25~ 0.05 mm)、粗 粉粒 (直徑 0.05~ 0.02 mm)、細粉粒 (直徑0.02~ 0.002 mm)、粘粒 (直徑小于 0.002 mm);全 N采用半微量開氏定氮法;全磷采用氫氧化鈉堿熔—鉬銻抗比色法;全鉀采用氫氧化鈉堿熔—火焰光度法;有機質采用重鉻酸鉀-外加熱法;pH值采用電位法;還原性物質總量采用 Al2(SO4)3浸提-K2Cr2O7滴定法;土壤含水量采用烘干法。具體操作參照《土壤理化分析與剖面描述》[16]。
1.3 統計分析
數據處理采用統計軟件 SAS8.2,多重比較采用Duncan檢驗,顯著度水平為0.05。因子間相關分析采用雙變量相關分析,選擇pearson相關系數。選擇單峰模型排序(CCA)用于反映環境因子對植物群落的影響,由 Canoco for windows 4.5軟件實現。
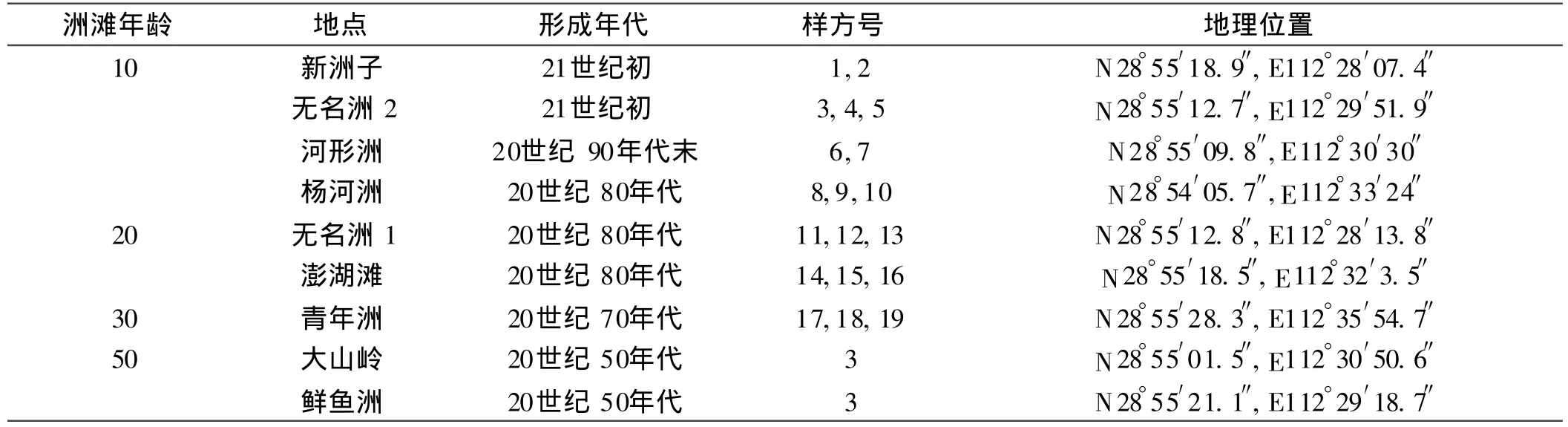
表1 試驗地概況
2 結果
2.1 植被特性
在9個洲灘51個樣方內共含種子植物67種,隸屬于 26科,57屬;其中物種數最多的科有禾本科(Gramineae)11種,菊科 (Compositae)8種,唇形科(Lamiaceae)6種。不同年齡洲灘相對密度、相對蓋度、相對高度、重要值及主要伴生種見表2。調查發現,該地以濕生植物為主,間或有少量中生植物。
黃土包區域不同年齡洲灘植被特征如表3。經方差分析,10年洲灘植物群落的生物量(2.96±1.66 kg/m2,p<0.05)、豐富度指數 (0.43± 0.09,p<0.05)和生物多樣性指數 (0.83±0.07,p<0.01)最低,益草和水芹為其建群種。而在20,30和50年洲灘中,植物群落生物量、豐富度指數和多樣性指數顯著增加,Wienner指數在 30年洲灘中達到最大值 0.75±0.13,而生物量和 Shannon and Wienner指數在20,30和50年洲灘中無顯著性差異(p>0.05),蘆葦和荻逐漸代替益草和水芹成為洲灘建群種。
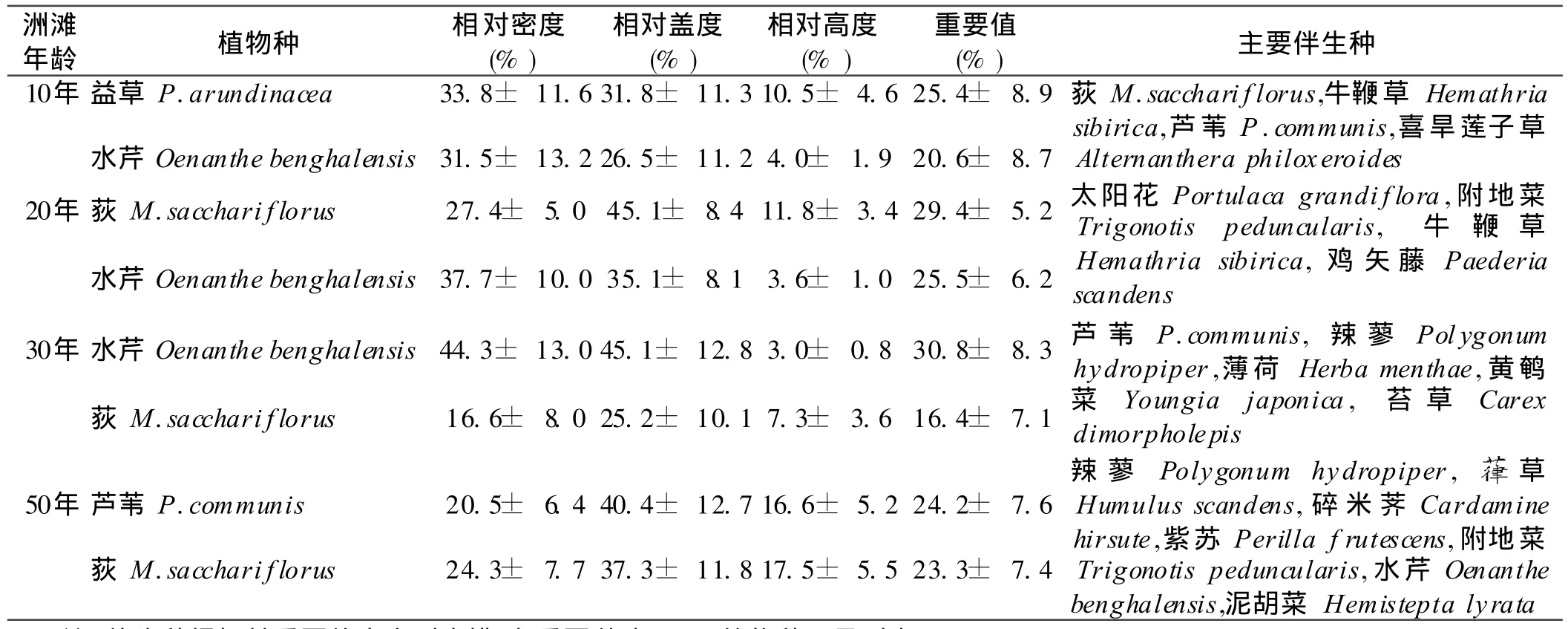
表2 黃土包河洲灘植被組成概況
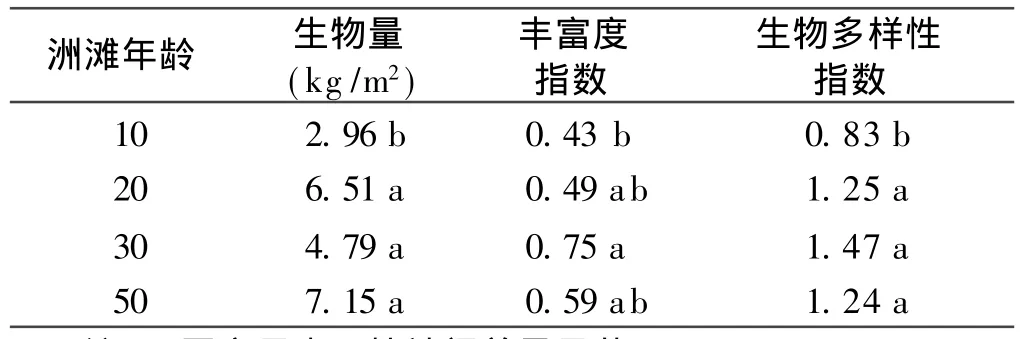
表3 不同演替階段洲灘濕地生物學特征
2.2 土壤特征
2.2.1 土壤物理特征
土壤物理特征如表4所示。在10年洲灘中,細砂粒所占百分比最大(62.2±4.4%),其次為細粉粒(21.6±2.7%)和粘粒(6.9± 1.2%)。隨著形成年份的增加,細砂粒和粗砂粒(>0.05 mm)極顯著減少,而細粉粒和粘粒百分比顯著增加。細砂粒百分比分別減少27.4%,47.2%和 47.6%,而細粉粒分別增加 14.6%,27.5%和 27.4%,粘粒分別增加 8.7%,22.9%和14.3%。但經方差分析,在20,30和50年洲灘間土壤顆粒組成無顯著性差異(p>0.05)。不同形成年份土壤間土壤含水量同樣存在顯著性差異(p<0.05),其在10年洲灘中含量最低(20.1±1.5%)。
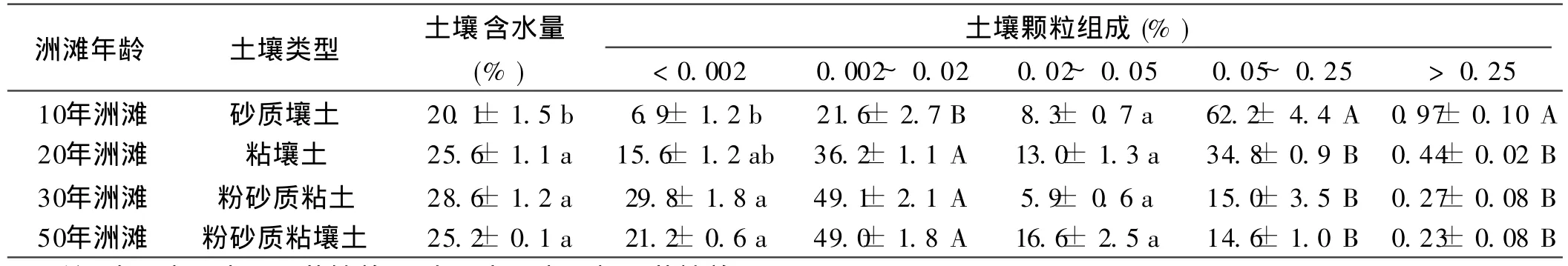
表4 不同年齡洲灘濕地土壤物理性質特征的空間分布特征
2.2.2 土壤化學特征
土壤化學性質如表5所示。10年洲灘土壤營養物質含量明顯低于20,30和50年洲灘。隨著洲灘年齡的增加,有機質含量從 0.68± 0.046%(10年洲灘 )分別增加至 1.30± 0.057%,1.53± 0.103%和 1.67±0.041%,呈極顯著增加的趨勢;全氮 (0.083±0.006%)和全鉀(20.51±0.44 g/kg)在 10年洲灘中含量最低,50年洲灘中含量最高;還原性物質總量和全磷在不同年齡洲灘間無顯著性差異。10年和20年洲灘土壤pH值大于 30和 50年洲灘,兩階段存在極顯著性差異。
2.3 土壤特征與生物學特征間的相關分析
相關分析表明,土壤細砂粒與絕大部分環境變量相關,例如,與pH和粗砂粒顯著正相關,而與有機質、粘粒、細粉粒、生物多樣性指數極顯著負相關,與土壤含水量、全鉀及生物量顯著負相關。這表明隨著土壤細砂粒百分比的減少,土壤理化性質及植被狀況得到改善,因此通過分析土壤細粉粒含量可以間接的得知不同年齡洲灘土壤和植被狀況,從而可以作為洲灘濕地演替的指示因子。
表5 不同演替階段洲灘濕地土壤化學性質特征的空間分布特征
2.4 CCA排序
將25個樣點及植被分別進行CCA排序。經分析,CCA的前兩個排序軸特征值分別為 0.639和 0.506,總特征值2.584。前兩軸累積貢獻率分別為 33.9%和61.7%。這表明本分析中環境變量可以較好的解釋群落結構變化,反映物種分布與性質之間的關系。根據前兩個排序軸做出二維排序圖(圖1),與第一軸相關性最高的環境變量是細砂粒百分比,同時還與有機質、粗砂粒、含水量、全鉀及粗粉粒極顯著或顯著相關。第二軸與全鉀相關性最高,此外還與pH、有機質、細粉粒、細砂粒及總還原性物質總量極顯著或顯著相關。
樣方和物種 CCA排序圖見圖 1,2。除河形洲(6,7)外,10年洲灘樣方集中在排序軸右側,此區間建群種為益草和水芹,它們對低營養條件、高砂粒含量及堿性環境有很強的忍耐力,能夠適應洲灘形成早期極度惡劣的環境。50年洲灘樣方集中在左下方,此區間土壤環境狀況最好,如營養物質含量最高,砂粒百分比最低。益草和水芹逐漸從群落中消失,蘆葦和荻最終成為該區間的建群種。20,30年洲灘樣方處在條件適中的左上方,為10年洲灘向 50年洲灘演變的過渡階段。
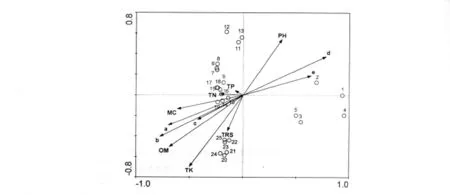
圖1 25個采樣點與土壤性質的CCA排序
3 討論
李景保等人研究表明[17],四水粒徑為 0.005~0.025 mm的泥沙含量占 53.8%~ 67.9%,而三口該粒徑級泥沙含量只占31.8%~ 42.6%;>0.05 mm粒徑級泥沙含量四水僅占 31.8%~42.4%,而三口卻占54.9%~68.7%。10年洲灘土壤顆粒組成狀況與其一致,結合黃土包河地理特征(承擔長江來沙20%~ 30%的輸沙任務),我們推斷 10年洲灘土壤顆粒組成主要由長江來沙的性質決定。隨著洲灘形成年份的增加,洲灘地勢逐漸抬高,灘地出露的時間逐漸增長,落淤的時間縮短,泥沙淤積對洲灘顆粒組成的影響逐漸減弱,而生物因素逐漸增強。相關分析表明,土壤細砂粒百分比與植物群落多樣性指數極顯著負相關,與生物量顯著負相關,而細粉粒和粘粒百分比與生物多樣性指數顯著正相關。
植物對土壤顆粒組成的影響主要表現在兩個方面:1)植物根系分泌的代謝物質進入土壤改變土壤的物理性質,同時植物細根不斷地生長、死亡,其分解產物極大地促進了土壤團聚體的形成[18,19];2)由于植物對侵蝕土壤的攔截作用,減弱了雨水對細粒徑顆粒的沖刷作用,使土壤中細粒徑顆粒含量增加[20]。土壤含水量的變化與土壤狀況的改善有關。相關分析表明,土壤含水量與粘粒和細粉粒顯著正相關,而與細砂粒顯著負相關。這是因為粘粒顆粒小,比表面積大,吸附能力強,保水也較強。此外,粘粒含量增加還促進了土壤的膨脹、可塑性及離子交換等物理性質[21]。綜上所述,泥沙淤積是造成 10年洲灘新生洲灘土壤物理性質和生物特征的主要因子,然而隨著洲灘形成時間的增加,生物因素對土壤的物理改良作用逐漸增強。
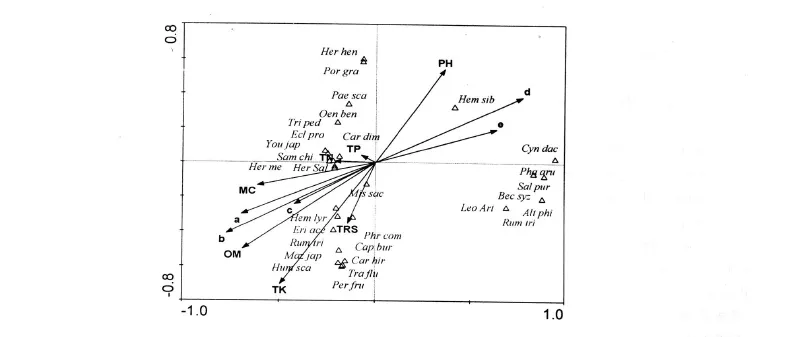
圖2 物種與土壤性質間的CCA排序
洲灘土壤化學性質的變化同樣受泥沙淤積和植被因素影響。從總的趨勢看,早期砂粒比重過大使土壤養分極易隨水體流失,同時強烈的淤積也不利植物的生長,植物凋落物很少,使土壤有機質、全鉀和全氮含量低。隨著洲灘濕地演替時間的延長,有機質、全氮、全鉀含量逐漸增加。這是因為:1)與地上植被有關。土壤有機質和全氮含量與植物凋落物的分解直接相關[22,23],相關分析表明,土壤有機質和全鉀含量與群落生物量顯著正相關;2)土壤溶液中的K+和 P含量與土壤粘粒含量相關[24]。相關分析發現土壤全鉀與粉粒百分比顯著正相關。可見,植被狀況的改善不僅直接增加了土壤營養物質的含量,還通過改良土壤性質增加了其保肥能力,從而間接促進了營養物質的積累。綜上所述,決定洲灘演變的因素有兩個,一個是泥沙淤積,決定新生洲灘的土壤和植被特征;另一個是植被因素,決定 10年后洲灘土壤理化性質。因此,決定洲灘演變的關鍵因子是不斷轉變的。由于本研究只對形成時間小于50年的洲灘進行了調查,50年后的洲灘其演替關鍵因子是否還會改變有待進一步研究。
[1]Maun M A.Adaptations of plants to burialin coastal sand dunes[J].Can J Bot,1997,76:713-738.
[2]Fennessy MS,Brueske CC,Mitsch WJ.Sediment deposition patterns in restored freshwater wetlands using sediment traps[J].Ecological Engineering,1994,3:409-428.
[3]Spencer DF,Ksander GG.Sedimentation disrupts natural regeneration ofZannichellia palustris in FallRiver,California[J].Aquatic Botany,2002,73:137-147.
[4]Owen NW,Kent M,Dale M P.Plant species and community responses to sand burial on the machair of the Outer Hebrides,Scotland[J].Journal of Vegetative Science,2004,15:669-678.
[5]Somé A.Les systè mes écologiques post-culturaux de la zone soudanienne(Burkina Faso)[A].Structure spatiotemporelle des communauté s vé g é tales et évolution des caracté ristiques pé dologiques[C].Paris:Doctoral Thesis,Pierre et Marie Curie Paris V I,1996.250.
[6]Torak LJ,Davis GS,Strain GA,et al.Geohydrology and evaluation of the water-resource potential of the upper Floridan aquiferin the Albany area,Southwestern Georgia[R].US Geological Survey Open-File Report,1991.91-52.
[7]Katherine Kirkman L,Charles Goebell P,Larry West,et al.Depressional wetland vegetation types:A question of plant community development[J].Wetlands,2000,20(2):373-385.
[8]Kent M,Owen N,Dale P,et al.Studies of vegetation burial:a focus for biogeography and biogeomorphology[J].Progress in Physical Geography,2001,25:455-482.
[9]Li F,Xie YH.Spacer elongation and plagiotropic growth are the primary clonal strategies used by Vallisneria spiralis to acclimate to sedimentation[J].Aquatic Botany,2009,91:219-223.
[10]謝永宏,李 鋒,侯志勇,等.澧水河口區于目平湖心區的基本生態特征[J].生態科學,2008,27(6):439-445.
[11]黃金國.洞庭湖區濕地退化現狀及保護對策 [J].水土保持通報,2005,4(12):261-263.
[12]彭佩欽.洞庭湖濕地形成演替與濕地生態研究建議[J].科學新聞,2007,(17):7-8.
[13]周樂新.三峽工程對洞庭湖河道、湖泊的影響及其演變趨勢研究[A].見:陳長嘉.三峽工程與洞庭湖關系研究[C].長沙:湖南科學技術出版社,2002.85-102.
[14]余德清,皮建高.洞庭湖區洲土變化特征與地殼沉降遙感研究 [J].湖南地質,2002,21(1):49-50.
[15]姜加虎 ,竇鴻身,黃 群.湖泊資源特征及與其功能的關系分析 [J].自然資源學報,2004,19(3):387-391.
[16]劉光崧.土壤理化分析與剖面描述 [M].北京:中國標準出版社,1996.
[17]李景保,尹 輝,盧承志,等.洞庭湖區的泥沙淤積效應[J].地理學報,2008,63(5):514-523.
[18]Marschner P,Rengel Z.Contributions of rhizosphere interactions to soil biological fertility[A].In:Abbott LK,Murphy DV.Soil Biological Fertility:A Key to Sustainable Land Use in Agriculture[C].ordrecht:Kluwer Academic Publishers,2003.81-89.
[19]魯素云.植物根際生態學與根病生物防治進展 [M].北京:中國人民大學出版社,1990.
[20]林超文,涂仕華,黃晶晶,等.植物籬對紫色土區坡耕地水土流失及土壤肥力的影響 [J].生態學報,2007,27(6):2191-2198.
[21]朱志梅,楊 持,曹明明,等.多倫草原土壤理化性質在沙漠化過程中的變化 [J].水土保持通報,2007,27(1):1-5.
[22]Van Oijen D,Feijen M,Hommel P,et al.Effects of tree species composition on within-forest distribution of understory species[J].Appl Veg Sci,2005,8:155-166.
[23]Greenson PE,Clark JR,Clark JE.Wetland Functions and Values:The State of Our Understanding [M].Minneapoils,Minn:American Water Resoures Assoc,1979.468-478.
[24]陳增奇 ,金 均,陳 奕.中國濱海濕地現狀及其保護意義 [J].環境污染與防治,2006,28(12):930-933.
猜你喜歡
數學小靈通·3-4年級(2024年2期)2024-05-15 02:02:28
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:36
科技傳播(2019年22期)2020-01-14 03:06:54
瘋狂英語·新策略(2019年10期)2019-12-13 08:43:28
當代陜西(2019年10期)2019-06-03 10:12:04
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
數學小靈通·3-4年級(2017年9期)2017-10-13 08:10:54
