馬鈴薯青枯病菌生化型研究及菌株接種方法的比較
2010-06-19 08:59:06王曉丹閔凡祥胡林雙董學志李學湛
中國馬鈴薯 2010年1期
關鍵詞:研究
王曉丹,閔凡祥,郭 梅,胡林雙,魏 琪,董學志,李學湛
(黑龍江省農業科學院植物脫毒苗木研究所,黑龍江 哈爾濱 150086)
馬鈴薯青枯病是由Ralstonia Solanacearum(RS)引起的一種世界性的重要細菌性病害,每年因此造成的產量損失大約為10%~15%,發病嚴重的地塊產量損失可達80%甚至絕產,給馬鈴薯生產帶來嚴重威脅。由于青枯病為細菌性病害,植株感病是系統性侵染,青枯病的藥劑防治十分困難。目前,防治青枯病最根本有效的方法是采用抗病品種,但是,馬鈴薯品種的抗病性對于不同的青枯菌系是有差異的[1]。因此鑒定和研究馬鈴薯青枯病菌生化型的組成和分布,將為加速我國馬鈴薯抗青枯病育種和病害綜合防治的工作進程提供必要的理論依據。
1 材料與方法
1.1 供試菌株
供試的10個菌株分別來自我國山東、廣東和廣西3省。菌株全部從送檢至我單位的馬鈴薯青枯病樣品中分離純化獲得。菌株來源情況見表1。用于分離純化的培養基為2,3,5-氯化三苯基四氮唑干酪素選擇性培養基(簡稱TZC)、用于純培養的培養基是基礎培養基(簡稱Kelman)[2]。
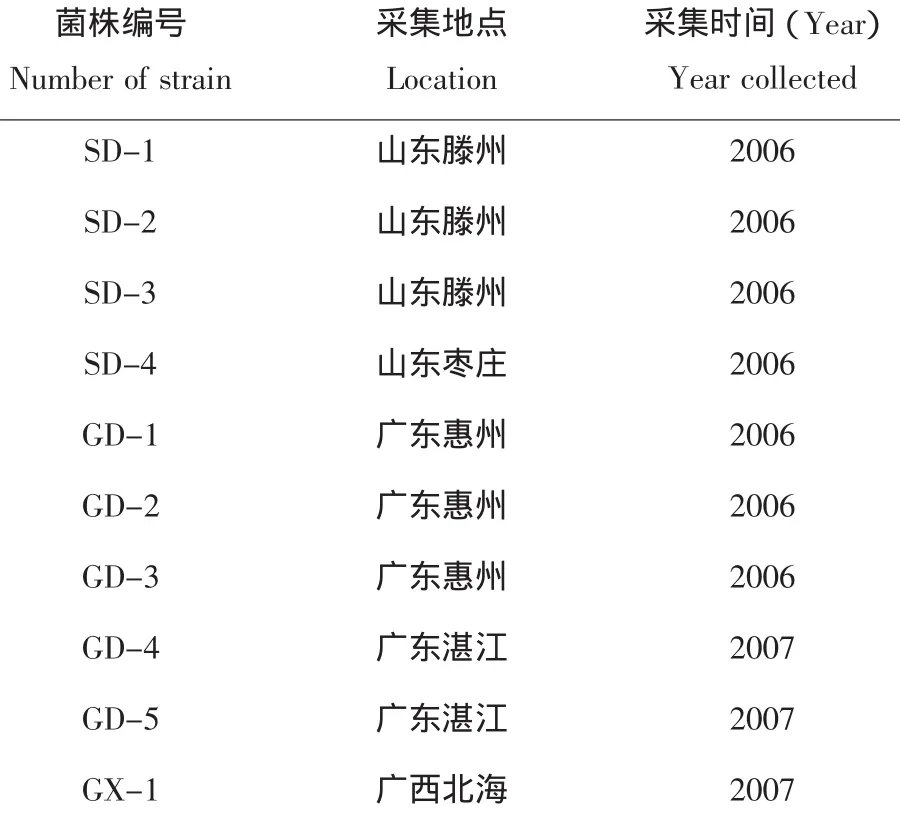
表1 供試菌株Table1 The tested strains
1.2 生化型鑒定
休和利夫森二氏培養基[3]:蛋白胨2 g、NaCl 5 g、K2HPO40.2 g、葡萄糖10.0 g、瓊脂6.0 g、1%溴百里酚藍溶液3 mL、蒸餾水1000 mL,pH 7.0~7.2,分裝于試管中,115℃蒸汽滅菌20 min。滅菌后,分裝試管,高度約為4.5 cm。配置10%濃度的纖維二糖、乳糖、麥芽糖、己六醇、甘露醇和山梨醇溶液,分別過濾滅菌,加入涼至60℃的休和利夫森二氏培養基中,終濃度為1%。
以18~24 h幼齡菌種作種子,穿刺接種至休和利夫森二氏培養基中,每株 3支。同時以不接種的 3管培養基作為陰性對照。30℃培養 1、2、3、7、14 d后,觀察結果。
1.3 生化型鑒定標準
若接種培養基產酸由橄欖綠變為黃色并且陰性對照不變色的判斷為陽性。生化型鑒定參照1964年Hayward[4]的劃分標準進行,見表2。
1.4 青枯菌接種方法
以番茄作為接種植物,采用傷根灌注法和刺莖法2種方法進行人工接種,并設清水對照,重復3次,篩選最佳接種方法。
(1)傷根灌注法:在溫室進行營養穴盤育苗,在5~6片葉時期,在幼苗的一側,距根1 cm處用刀造成傷口,每株番茄苗在傷根部位灌注50 mL菌液(菌液濃度大約為1×109 cfu·mL-1)。將接種后的番茄苗放入人工氣候箱內,設定溫度30℃,光照16 h,黑暗8 h,空氣濕度保持在80%以上,定期澆水,保持土壤濕潤。若濕度無法達到,則可以用一個塑料袋包住植株,植株頂部將塑料袋開一小口,以保持其濕度,促進發病。在接種后7、14、21 d觀察發病情況。
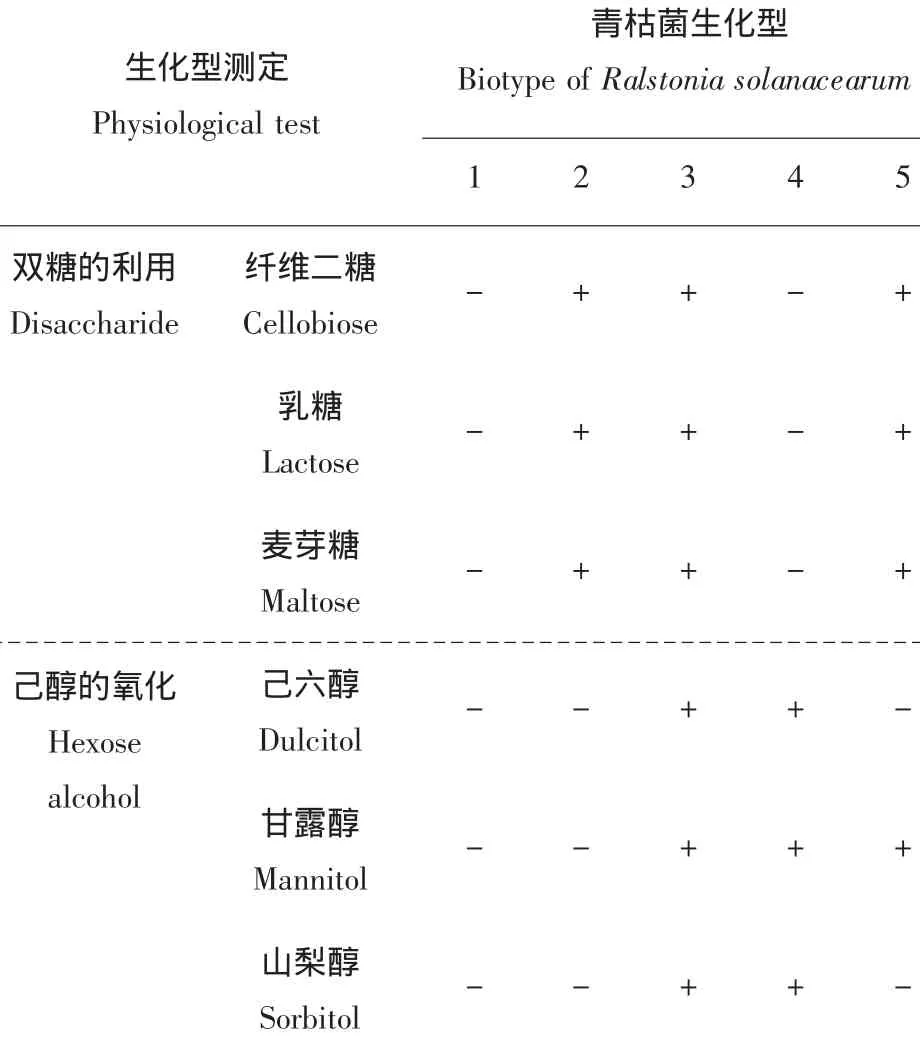
表2 根據對雙糖的利用和己醇的氧化產生酸時為陽性(+)對青枯菌分類Table2 Classification of Ralstonia solanacearum based on the ability to utilize disaccharides and oxidize hexose alcohols producing acid when it is positive(+)
(2)刺莖法:將滅好菌的牙簽挑取青枯菌菌液若干,直接插到第3節葉腋處,每1株插1根,插牙簽時要注意不用插得太深,以免植株在未發病前就萎蔫死亡。放入人工氣候箱培養,培養條件同上。
在接種后7、14、21、28 d,按Winstead等[5]分級標準進行分級調查,共分5級,分級標準為:0級:不發病;l級:1片葉感病;2級:2或3片葉感病;3級:除了頂端2~3片葉其余葉片均感病;4級:所有葉片感病;5級:植株完全死亡。
2 結果與分析
2.1 生化型測定
對來源于我國3個省份的從馬鈴薯寄主上分離的10個青枯病菌的生化型鑒定,結果見表3,測定結果可以看出:生化型Ⅱ和Ⅲ為優勢種群,均占被測菌株的40%,生化型Ⅰ和生化型Ⅳ分別占10%。從地域上來看,被檢樣品中,我國山東省馬鈴薯青枯病菌生化型有3種,分別為生化型Ⅱ、Ⅲ和Ⅳ;廣東省馬鈴薯青枯病菌生化型有2種,分別為生化型Ⅱ和Ⅲ;由于廣西省被檢樣品數量有限,發現1種生化型,為生化型Ⅰ。
2.2 病原菌接種
試驗結果表明,利用傷根灌注法接種馬鈴薯青枯病菌的效果好于刺莖法。前者接種的番茄苗14 d內均發病,發病較后者快;傷根灌注法接種的番茄苗發病級數都為4級,而刺莖法發病級數為2~3級。從操作難易程度來看,傷根灌注法比較簡單,而刺莖法操作時要掌握好力度,容易傷及幼苗。因此,綜合分析,馬鈴薯青枯病接種方法以傷根灌注法較好。
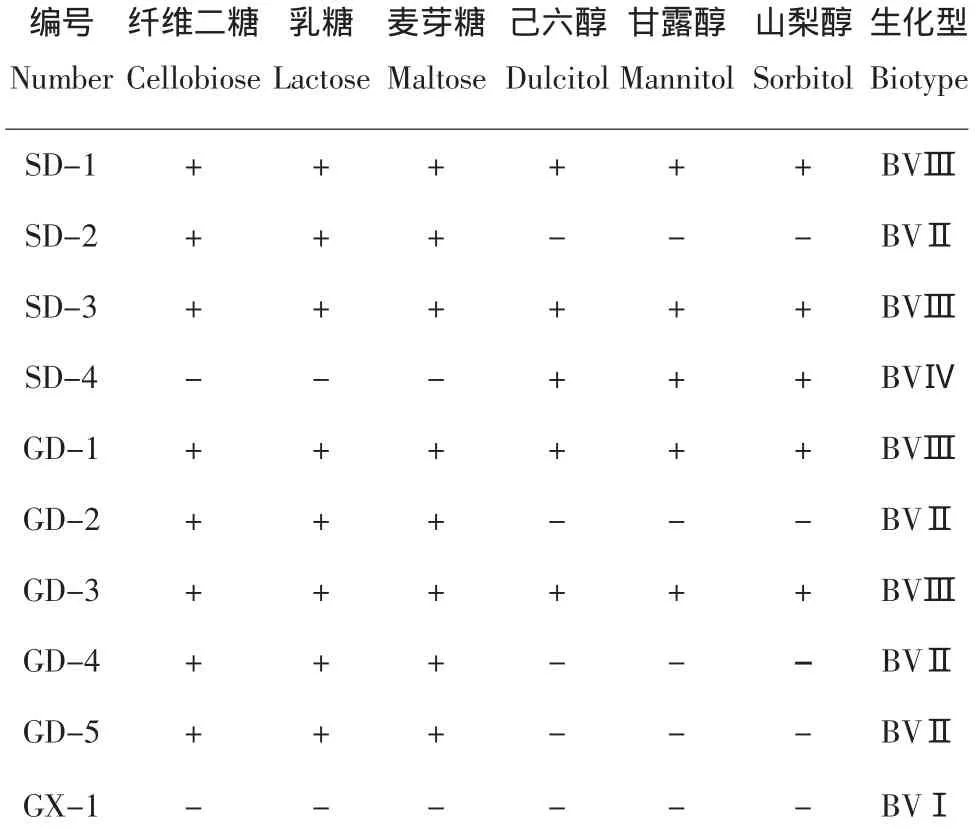
表3 被測菌株生化型測定結果Table3 The classification results of tested strains
3 討論
青枯病菌生化型在我國不同地區和不同植物上的分布狀況是相當復雜的,據國外資料記載在馬鈴薯上4種生化型都有存在[2]。本研究的被測馬鈴薯青枯病菌菌株包含了上述全部4種生化型。1985年,華靜月等[1]對我國8個省市的43個青枯菌菌株測定生化型,結果表明,供試的絕大部分菌株屬生化型Ⅱ,少數屬于生化型Ⅲ和Ⅳ。本研究中的優勢種群為生化型Ⅱ和Ⅲ。從地域上看,本研究的被測廣東省菌株中,生化型Ⅲ為優勢種群,占60%。曾憲銘等[6]測定廣東農作物青枯病菌生化型的結果是其所測青枯病菌在馬鈴薯寄主上的生化型為Ⅱ、Ⅳ和Ⅲ-2。本研究中山東省馬鈴薯青枯病菌以生化型Ⅲ為優勢種群,占50%,與丁愛云等[7]報道的山東省馬鈴薯青枯病菌以生化型Ⅲ為主的結果一致。
1976年Harris[8]認為,可以把Hayward模式中的生化型進一步劃分為亞型,自1984年華靜月等[9]研究報道了我國青枯病菌有生化亞型的存在,之后又有曾憲銘等[6]、丁愛云等[7]、徐莉莉等[10]報道了我國青枯病菌生化亞型的存在。在本研究中沒有測定到被測菌株中有生化亞型。
由于本研究的菌株數量有限,特別是廣西省收集到的菌株數量很少,因此本研究的結果尚不能代表上述3省份的所有馬鈴薯青枯病菌的生化型種類及優勢種群,有待于今后加大菌株的收集量,做更詳細地研究。
[1]華靜月,張長齡,何禮遠.我國馬鈴薯青枯菌菌系和初步研究[J].植物病理學報,1985,15(3):181-184.
[2]French E R,Gutarra L,Aley P,et al.Culture media for Pseudomonas solanacearum:Isolation,identification and maintenance[J].Fitopatologia,1995,30:126-130.
[3]東秀株,蔡妙英.常見細菌系統鑒定手冊[M].北京:科學出版社,2001.
[4]Hayward A C.Characteristic of Pseudomonas solanacearum[J].Appl Bact,1964,27(2):265-277.
[5]Winstead N,Kelman A.Inoculation technique for evaluating resistance to Psedomonas solanacearum[J].Phytopathology,1952,42:628-634.
[6]曾憲銘,董春.廣東農作物青枯病菌的生化型[J].華南農業大學學報,1995,16(1):50-53.
[7]丁愛云,鄭繼發,時呈奎,等.山東省植物青枯菌生化型研究[J].山東農業大學學報,2002,33(1):50-52.
[8]Harris D C.Bacterial wilt in Kenya with particular reference to potatoes[M]//Sequeira L,Kelman A.Proceedings of the first international planning conference and workshop on the ecology and control of bacterial wilt caused by Pseudomonas solanacearum.North Carolina State University,Raleigh,1976:84-88.
[9]華靜月,張長齡,何禮遠.我國植物青枯菌的生化型和其他生理差異[J].植物保護學報,1984,11(1):43-50.
[10]徐莉莉,尹燕妮,王勇明,等.發生于中國的青枯病菌生化變種研究[J].江蘇農業科學,2008(5):102-10.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
