γ-氨基丁酸在畜牧生產上的應用
2010-09-03 10:32:42內江職業技術學院李靜
中國飼料 2010年3期
關鍵詞:影響
內 江 職 業 技 術 學 院 李 靜
四川農業大學動物營養研究所 李 霞
γ-氨基丁酸 (GABA)是一種非蛋白質氨基酸,廣泛分布于動植物中。在動物體內不僅分布于中樞神經系統和外周神經,起神經鎮定作用;也分布于非神經組織的細胞中,發揮激素或營養因子的功能,對動物機體正常的生理功能起著重要的調節作用(Xu等,2001)。隨著研究深入,GABA的生理功能不斷得到闡明,已發展成為一種新型功能性因子,并伴隨著生產技術的逐步成熟,已逐漸在醫藥、食品保健、化工及農業等行業中廣泛應用。在飼料行業中,GABA作為一種功能性氨基酸類飼料添加劑,具有提高動物生產性能、免疫功能和產品品質等多種功效,已成為研究熱點,因此本文介紹GABA在畜牧生產上的應用情況。
1 GABA的結構與理化性質
GABA在丁酸的γ位上有一個氨基,是一種非蛋白質氨基酸,分子式為C4H9NO2,相對分子質量為103.12,結構式見圖1。
GABA結構與谷氨酸類似,只少了一個羧基,為其脫羧產物。GABA在氣相時主要以中性分子的形式存在,而在晶體狀態下則以兩性離子的形式存在,在GABA水溶液中也存在著大量兩性離子。
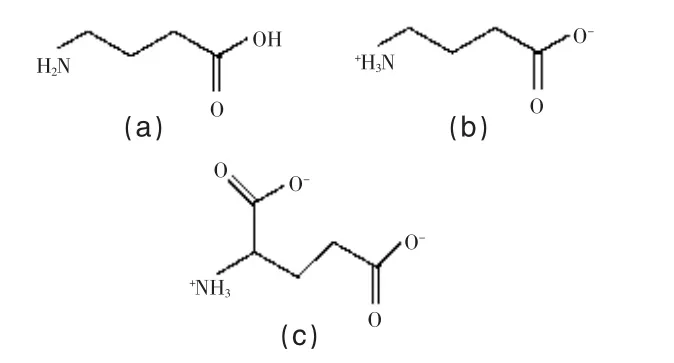
圖1 GABA 的中性(a)、兩性(b)離子結構和谷氨酸的兩性離子結構(c)
GABA極易溶于水,25℃時溶解度為130 g/100 mL,微溶于乙醇,不溶于其他常見有機溶劑,熔點202~204℃,但在195℃時即分解為吡咯烷酮和水。結晶為白色小葉狀(甲醇-乙醚)或針狀(水-乙醇)),味微苦,無旋光性。在水溶液中,易發生兩性離解,也可進行氨基酸的特征性化學反應。
2 GABA在畜牧生產中的應用
2.1 GABA對動物生產性能的影響及其原因
2.1.1 對動物生產性能的影響 韋習會等(2004)在生長豬日糧中添加不同濃度 (0、10、20 mg/kg和40 mg/kg)的GABA,結果表明,10 mg/kg和20 mg/kg的添加水平能顯著提高日增重(分別提高11.39%和9.49%),而添加40 mg/kg對增重無顯著影響。而其增重的提高主要是通過提高日采食量(提高7.82%和9.47%),對料肉比無顯著影響;與對照組相比,10 mg/kg的添加劑量對血清尿素氮含量無顯著影響,而高劑量的GABA則使豬血清尿素氮含量顯著高于對照組。Fan等(2007)在體重為 34 kg生長豬日糧中添加50 mg/kg和100 mg/kg GABA,結果表明,GABA對豬的促生長作用明顯,但隨生長階段和作用劑量不同存在一定差異,主要體現在試驗3~4周,以100 mg/kg的效果最好。劉振軍(2006)在21 d斷奶仔豬日糧中分別添加50、100 mg/kg和150 mg/kg GABA,發現50 mg/kg添加劑量能顯著提高斷奶后28 d的采食量和日增重,但不影響料肉比和粗蛋白質的表觀消化率。隨著劑量的增加,GABA的效果降低,當添加劑量為150 mg/kg時,斷奶仔豬的生產性能有下降的趨勢。謝曉婕(2007)在28日齡斷奶仔豬日糧中添加20 mg/kg GABA,飼喂14 d,提高了仔豬采食量、日增重,降低0~7 d料重比,但差異不顯著。
李愛學(2003)在產蛋高峰期母雞飼糧中添加50 mg/kg GABA,母雞采食量和采食次數均有所增加。吳常信(2005)報道,在肉雛雞前期日糧中添加50、100 mg/kg和200 mg/kg GABA,日增重相對于對照組分別提高3.6%、7.6%和10%,而50 mg/kg和100 mg/kg處理組飼料消耗分別降低了4%和8%,200 mg/kg組料肉比無顯著差異;在肉雛雞后期日糧中添加75、150 mg/kg和300 mg/kg GABA,日增重結果與前期相似,75 mg/kg和150 mg/kg處理組飼料消耗降低15%和18%,而300 mg/kg組體重和飼料消耗沒有改善。陸桂平等 (2007)在產蛋雞日糧中添加10 mg/kg GABA,青年期、產蛋初期和產蛋高峰期蛋雞的采食量分別提高 11.9%(P<0.01)、5.95%(P<0.05)和 7.23%(P<0.01),對于產蛋性能,只顯著或極顯著提高了產蛋初期的產蛋率 (15.59%)和日只蛋重(17.86%)。
從以上結果可以看出,GABA主要是通過提高畜禽的日采食量,且不增加或略微提高飼料轉化效率來實現對增重的提高,GABA的添加效果與添加劑量有關,不同動物和不同階段添加劑量不同。
2.1.2 GABA提高動物生產性能的原因
2.1.2.1 采食作為一個復雜的行為活動,主要受中樞神經系統的控制,飽中樞(下丘腦腹內側核)和攝食中樞(下丘腦外側區)是動物調節攝食的基本中樞。GABA則可通過抑制飽中樞(下丘腦腹內側核)的活動引起動物的采食,增強動物的采食能力。研究表明,將一定劑量范圍的GABA注射于動物的不同腦區,可顯著促進動物攝食,并具有劑量依賴效應。
2.1.2.2 可能與其促進生長激素等的分泌有關。Tacca等(1989)發現,激活離體鼠胃 GABA受體能夠劑量依賴性誘導胃泌素分泌。Weigert等(1998)認為,灌注外源性GABA濃度,劑量依賴性提高胃泌素釋放,而生長抑素(SS)的分泌則被顯著性抑制;并且生長抑素和胃泌素對于GABA的反應能夠完全被γ-氨基酸A受體(GABAAR)拮抗劑荷包牡丹堿和乙酰膽堿能神經元阻斷劑阿托品完全抑制。陸桂平等(2007)研究發現,飼喂10 mg/kg的GABA使青年期蛋雞、產蛋初期和產蛋高峰期母雞血清瘦素水平分別升高4.47%、29.48%(P<0.05)和 21.16%(P<0.05),血清胰島素水平分別升高 21.15%(P<0.01)、15.13%(P<0.05)和 12.85%(P<0.05)。 瘦素具有調節攝食、促進能量消耗及調節相關激素水平和生殖機能等作用。胰島素水平升高可使攝食中樞興奮,促進采食。Fan等(2007)在生長豬日糧中添加50 mg/kg和100 mg/kg GABA,結果表明,GABA顯著促進了試驗豬體內生長激素(P<0.05)和褪黑素(P<0.05)的分泌,明顯提高了小豬體內促甲狀腺激素水平(P<0.1),這與良好的生產性能吻合,但作用程度隨試驗階段和激素種類不同而存在一定差異。
2.1.2.3 與GABA促進動物胃腸道發育有關。GABA 可促進胃酸分泌。Thirlby(1998)等,給安裝胃瘺的狗胃內灌注GABA,發現54 μg/kg·min注射組狗胃內平均酸分泌量在基礎水平上提高了(30±1.4)mmol/h; 另外靜脈給藥 0.5 mg/kg baclofen(GABA激動劑)能夠顯著提高狗胃內酸的分泌量。Laura(2004)在研究外源GABABR激動劑對大鼠胃酸分泌的試驗中發現,靜脈給藥baclofen和SKF-97541(GABA選擇性受體激動劑),能夠對大鼠胃酸分泌起到劑量依賴性的刺激作用。由此可以得出GABA及其受體激動劑具有顯著的促酸分泌的作用。
GABA可促進小腸各腸段收縮和舒張運動。Giotti(1983)發現,在 3×10-6~ 3×10-4mmol/L 劑量范圍內,GABA可引起體外培養的豚鼠回腸標本收縮,緊接著又成舒張狀態。Krantis等(2000)對培養的豚鼠結腸末端和回腸的研究證明,GABA對沒有神經支配的腸肌沒有作用。在有神經支配的腸段,GABA所引起的收縮和舒張都可被河豚毒素阻斷,表明GABA通過激活神經性的興奮和抑制機制對腸段發揮作用。
有研究報道,產蛋高峰期母雞飼料中添加GABA可顯著提高十二指腸至盲腸段內容物中的胰蛋白酶總活性(P<0.05)。表明GABA對小腸功能的調節很可能是通過影響其消化酶的分泌來實現(李愛學,2003)。
在豚鼠和大鼠小腸上的試驗已證實GABA能影響上皮的轉運過程。另外,腸道隱窩處纖維與上皮組織中含有GABA,與內分泌細胞存在相似之處,提示在黏膜功能調節上,腸道GABA很可能影響上皮細胞的有絲分裂和遷移(Krantis,2000)。
2.2 對胴體品質的影響 曹德瑞和鄒曉庭(2007)在生長育肥豬日糧中添加10 mg/kg GABA,可使屠宰率提高3.64%,眼肌面積增加11.89%,背膘厚略有降低,但差異不顯著。GABA改善動物胴體品質的原因可能與其促進生長相關激素的合成與釋放有關,因為GABA可促進生長激素的分泌,而大量試驗表明,外源生長激素可改善豬的胴體品質。
2.3 對免疫功能的影響 Adham等(2006)研究表明,GABA對黏膜免疫具有一定增強作用。人口服100 mg/kg GABA后顯著提高唾液中IgA的分泌水平,從而增強機體抵御病原侵入,提高免疫力。具體的研究證明較少,但從旁分析GABA也具有提高免疫的功能,分析其原因可能是:一方面主要是GABA促進胃酸、消化液和生長激素的分泌,提高動物的采食量,增強養分吸收與代謝,提高生長性能從而提高免疫機能;另一方面GABA作為神經遞質,影響內分泌系統激素的分泌,然而激素分泌量的變化必然使存在于免疫細胞上的各種激素受體作出應答,從而引導免疫細胞自身增殖或凋亡、活性增強或抑制;另外,GABA是抑制性神經遞質,與受體結合后被激活,阻止與焦慮或應激有關的信息抵達大腦,鎮靜神經,抵抗焦慮和減輕應激程度,增強免疫力(胡家澄等,2008)。
2.4 GABA對生殖性能的影響 GABA在雄性動物或人性腺和附屬性腺中存在,與精子的運動和類固醇激素的產生有密切關系,對雄性生殖功能有調節作用。據報道,GABA可明顯地引起獲能小鼠及人精子發生頂體反應,且隨精子獲能進程而顯著增加,并存在明顯的量效關系 (Shi和Roldan,1995; 王春年,1995;Roldan 等,1994);明顯提高正常及抗精子陽性精子頂體酶的活性,并使精子的Na+-K+-ATPase和超氧化物歧化酶的活性增加,說明GABA對精子頂體酶活性有顯著影響。
在雌性動物中,在機體30多種含有GABA的外周組織中,以卵巢與輸卵管中的含量最高,超過腦含量的2倍以上,提示GABA可能對性腺的生理功能具有調節作用(Kalia等,1999)。據研究,GABA能夠通過下丘腦-垂體-性腺軸影響垂體及性腺的生理機能,從而參與生殖激素(如孕酮、促黃體素LH及促乳素PRL)的分泌調節。
2.5 GABA對畜禽的抗缺氧能力的影響 GABA可提高海馬腦的耐缺氧能力,其機制可能與GABA通過GABA受體提高Cl-內流有關 (趙彤等,2003)。GABA作為體內一種主要的抑制性氨基酸,在腦缺血時其合成與釋放均增加,這與興奮性氨基酸、自由基等毒性物質含量的增高有關。腦缺血后針對興奮性氨基酸的毒性作用,機體建立相應的“抑制性保護”機制,這一保護性機制的建立,主要是通過GABA的作用來完成 (Shuaib等,1997)。魏智清等(2006)試驗證明,適當濃度的GABA可以提高鯽魚的抗缺氧能力,其中質量分數0.3%GABA使鯽魚在缺氧環境下的存活時間明顯延長。
2.6 GABA對動物熱應激的影響 由于畜禽皮膚上缺少甚至沒有汗腺,在炎熱的夏季高溫所引起的熱應激給畜禽生產帶來嚴重的損害。GABA為中樞神經系統抑制性神經遞質,可抑制腦干呼吸中樞的整合作用,使呼吸頻率減慢,維持各中樞對機體各系統正常調節機能。陳忠等(2002)試驗證明,在肉仔雞飲水中添加適量GABA,可以減少雞只的活動量和產熱量,降低代謝率,從而緩解高溫對肉雞生產性能的影響,增強肉雞的抗應激能力和日增重。吳東輝等(2003)試驗證實,在急性應激時可通過海馬內興奮性氨基酸抑制劑GABA含量的變化來影響興奮性氨基酸含量的改變,從而對動物機體起到保護作用。曹德瑞等(2008)試驗表明,在生長育肥豬日糧中添加10 mg/kg的GABA可提高高溫下豬的采食量和增重;提高豬的血糖含量,維持機體熱應激狀態下新陳代謝所需的血糖;降低血液乳酸脫氫酶(LDH)和肌酸激酶(CK)水平,有效減輕熱應激對豬只的損害;提高豬血清谷胱甘肽過氧化物酶水平,緩解熱應激。
3 GABA的制備
3.1 化學合成方法 化學合成GABA的方法主要有以下幾種(趙炳超等,2004):(1)以鄰苯二甲酰亞氨鉀和γ-氯丁氰或丁內酯為原料在強烈條件下反應,所得產物與濃硫酸作用后再經過水解制得產品GABA;(2)以吡咯烷酮為起始原料,經氫氧化鈣、碳酸氫銨水解開環制得產品GABA;(3)以丁酸和氨水為原料,在γ射線照射條件下,可以得到GABA;(4)以丙胺和甲酸為原料,經輝光放電電解可以合成出GABA;(5)以溴代乙酸甲酯和乙烯為原料,經聚合反應得到4-溴丁酸甲酯,再經氨解和水解可制得。GABA化學方法反應速度快,得率高,但脫除產品中的有毒成分比較復雜,成本較高,反應條件不易控制,安全性較差。
3.2 分離提取方法 以天然產物或天然產物經過加工后的副產物為原料,經過提取、分離、提純制得。如以稻谷加工后的副產物米糠為起始原料,經過檸檬酸緩沖液抽提、過濾,所得濾液經冷凍干燥得到黃棕色的固體粗產物,然后經層析柱分離提純制得純品GABA。分離提取方法雖然比化學方法易操作,但成本仍然較高。
3.3 富集技術 高等植物中GABA的合成主要由 L-谷 氨 酸 脫 羧 酶 (Glutamatedecarboxylase,GDC,EC 4.1.1.15)催化 L-谷氨酸(L-Glu)的脫羧反應生成,而GABA的降解是在GABA轉氨酶的作用下與α-酮戊二酸發生轉氨作用,形成谷氨酸和琥珀酸半醛(SSA),琥珀酸半醛經脫氫氧化形成琥珀酸,進入三羧酸循環。因此有利于L-谷氨酸脫羧反應,不利于GABA轉氨作用的條件均有利于植物中GABA的合成。例如,細胞質pH值降低,有利于谷氨酸脫羧,不利于GABA轉氨作用(最適pH 8.9),從而積累GABA;Ca2+或鈣調蛋白(CaM)可激活谷氨酸脫羧酶;L-谷氨酸含量增多可使GABA支路中碳流量增加,并調節谷氨酸脫羧酶的活性。
目前已開發一些GABA富集產品,比如利用糙米或米糠自身的谷氨酸脫羧酶,提供發揮此酶活性最佳的溫度、pH、壓力及底物等條件,使發芽糙米或米糠中GABA含量升高。還可利用茶葉、果品、蔬菜、酒糟等植物來生產GABA富集產品。另外,通過遺傳和基因工程手段,也可培育高產GABA的作物品種,生產GABA富集產品。
3.4 生物合成方法 GABA的生物合成主要是以谷氨酸或其衍生物(谷氨酸鈉,或富含谷氨酸的物質等)為原料,利用微生物發酵制得。具有操作條件溫和、成本低、產品含量高的優點,在實際生產中,與化學合成法和分離提取法相比具有很大的優勢,但高效的微生物菌種通常較難獲得。早先主要利用大腸桿菌作菌種來發酵,但大腸桿菌存在著許多安全方面的隱患,目前發現一些安全性很高的微生物菌種,如乳酸菌、酵母菌以及曲霉菌等也可分泌谷氨酸脫羧酶,因此提高了生物合成GABA產品的安全性。劉清等(2004)得到一株可高產谷氨酸脫羧酶的乳酸菌菌株,在33℃進行發酵培養3 d后,發酵液中GABA含量可達到310 g/L以上,比優化前提高了約4倍。
通常發酵法制備高GABA含量的工藝流程如下:米胚芽提取物→添加碳源和氮源等配制培養基→接種酵母菌或乳酸菌→發酵→過濾→干燥→GABA成品。
4 GABA毒性及安全
關于GABA毒性研究較少。楊海峰等(2008)采用改良寇氏法測定GABA的急性毒性,及劑量遞增法測定其蓄積毒性,結果表明,GABA對小鼠的經口LD50為15.55 g/kg,屬實際無毒物質,基本無蓄積毒性。GABA作飼料添加劑使用時,在飼料中正常添加量僅為10~100 mg/kg(Fan等,2007;韋習會等,2004)。由于大多數報道生物合成方法制備GABA所用的微生物大都具有潛在的不安全因素,因此選用生物合成的GABA仍然需要注意。
5 小結
隨著GABA生理功能研究的不斷深入,GABA在畜牧生產中的應用已成為新的研究熱點。GABA作為飼料添加劑可以顯著提高畜禽的生產性能,因此,加強GABA的安全性能測試及研制開發將豐富飼料添加劑的種類,有益于畜牧業的發展。
[1]曹德瑞,鄒曉庭,顧林英.γ-氨基丁酸對溫熱環境中生長肥育豬生產性能和抗氧化功能的影響[J].中國飼料,2008,5:21~26.
[2]曹德瑞,鄒曉庭.γ-氨基丁酸對生長肥育豬生長性能和胴體品質的影響[J].飼料工業,2007,28(23):31 ~ 34.
[3]陳忠,王婷,黃麗明.γ-氨基丁酸對熱應激仔雞生產性能影響的研究[J].海南師范學院學報(自然科學版),2002,15(1):82 ~ 83.
[4]胡家澄,鄒曉庭,董金格.γ-氨基丁酸對肥育豬免疫機能的影響及其作用機理[J].飼料工業,2008,29(14):3 ~ 5.
[5]李愛學.γ-氨基丁酸和氟安定對產蛋高峰期母雞攝食行為以及有關內分泌的影響[D].碩士學位論文,南京農業大學,2003.
[6]劉清,姚惠源,張暉.生產γ-氨基丁酸乳酸菌的選育及發酵條件優化[J].氨基酸和生物資源,2004,26(1):40 ~ 43.
[7]劉振軍.γ-氨基丁酸對早期斷奶仔豬生長性能和血液指標的影響[D].碩士學位論文,四川農業大學,2006.
[8]陸桂平,左偉勇,陸熹.γ-氨基丁酸對蛋雞生產性能的影響[J].中國家禽,2007,29(23):51 ~ 52.
[9]王春年,劉海衛,袁玉英,等.γ-氫基丁酸誘發人精子頂體反應及其對若干離子轉運影響的研究[J].生殖與避孕,1995,16(2):118 ~ 121.
[10]韋習會,漆興桂,夏東,等.日糧添加γ-氨基丁酸對育肥豬生長和飼料利用的影響[J].家畜生態,2004,25(2):10 ~ 12.
[11]魏智清,楊涓,趙紅雪,等.牛磺酸,γ-氨基丁酸對鯽抗缺氧能力的影響[J].淡水漁業,2006,36(1):7 ~ 10.
[12]吳常信.中國家禽研究[M].成都:四川科學技術出版社,2005.354~355.
[13]吳東輝,楊權,許崇濤.應激對大鼠海馬谷氨酸、天冬氨酸和γ-氨基丁酸含量的影響[J].中華精神科雜志,2003,36(2):98 ~ 100.
[14]謝曉婕.γ-氨基丁酸和噻路酮對斷奶仔豬生產性能和消化道發育的研究[D].碩士學位論文,四川農業大學,2007.
[15]楊海峰,葛竹興,郁杰.γ-氨基丁酸的急性毒性和蓄積毒性的研究[J].安徽農業科學,2008,36(13):5464 ~ 5491.
[16]趙炳超,石波,李秀波,等.新型飼料添加劑-γ-氨基丁酸的制備及應用研究進展[J].中國畜牧獸醫,2004,31(12):13 ~ 14.
[17]趙彤,王福莊,黃燕華.GABA對大鼠海馬腦片缺氧損傷的保護作用[J].中國應用生理學雜志,2003,19(1):16 ~ 19.
[18]Adham M,Higashiguchi S,Horie K,et al.Relaxation and immunity enhancement effects of gamma-Aminobutyric acid(GABA)administration in humans[J].Biofactors,2006,3(26):201 ~ 208.
[19]Fan Z Y,Deng J P,Liu G H,et al.Effects of γ-Amino butyric acid on the performance and internal hormone levels in growing pigs[J].Chinese Journal of Animal Nutrition,2007,19(4):350 ~ 356.
[20]Giotti.GABAAand GABABreceptor mediated effects in guinca-pig ileum[J].Br J Phar-macol,1983,78:469 ~ 478.
[21]Kalia V,Hole D R,Willson C A.Effect of gonadal steroids and gammaaminobntyric acid on LH Release and dopmine expressionand activity in the zona in certa in rats[J].Reproduction and Fertility,1999,117:189 ~ 197.
[22]Krantis A.GABA in the Mammalian Enteric Nervous System[J].News Physiol Sci,2000,15:284 ~ 290.
[23]Laura.Peripheral GABABagonists stimulate gastric acid secretion in mice[J].British Journal of Pharmacology,2004,142:1038 ~ 48.
[24]Roldan E R S,Murase T,Shi Q X.Exocytosis in spermatozoa in response to progestorene zona pellueida[J].Science,1994,266:1578 ~ 1581.
[25]Shi Q X,Roldan E R S.Evidance that a GABAA-like receptoris involved in progesterone-induced acrosomal exocytosis in mouse spermatozoa[J].Biol Reprod,1995,52:373 ~ 381.
[26]Shuaib A,Ijaz M S,Miyashita H,et al.GABA and gluamate levels in the substantia nigra reticulation following repeatitive cerebral ischemia in gerbils[J].Exp Neurol,1997,147:311 ~ 315.
[27]Tacca M D,Blandizi C,Bemardinictal M C.GABA-A-mediated gastrin release induced by baclofen in the isolate dvascularly Perfosed rat stomach[J].Experientia,1989,45(8):734 ~ 736.
[28]Thirlby.Effect of GABA on basal and vagally mediated gastric acid secretion and hormone release in dogs[J].Physiol Gastrointest Liver Physiol,1988,254:723 ~ 31.
[29]Weigert N,Schepp W,Haler A,et al.Regulation of gastric,somatostatin and bombesin release from the isolated rat stomach by exogenous and endogenous garmma-amino butyric[J].Digestion,1998,59(1):16 ~ 25.
[30]Xu X G,Yang Z F,Huang S H,et al.Promotive effects of GABA on acid secretion from isolated mouse stomach in vitro[J].Acta Zool Sin,2001,47(2):170 ~ 175.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
