禽流感H5N1亞型病毒對五個不同品系小鼠致病性的比較
2010-09-09 09:20:30黃韌王穎彥鄧少嫦黃培根張鈺劉忠華吳玉娥李文德閔凡貴劉香梅
中國實驗動物學報 2010年4期
黃韌,王穎彥,鄧少嫦,黃培根,張鈺,劉忠華,吳玉娥,李文德,閔凡貴,劉香梅
(1.廣東省實驗動物監(jiān)測所廣東省重點實驗室;2.廣東醫(yī)學院,湛江 524023;3.哈佛醫(yī)學院麻省總醫(yī)院放射腫瘤系)
研究報告
禽流感H5N1亞型病毒對五個不同品系小鼠致病性的比較
黃韌1,王穎彥1,鄧少嫦1,黃培根3,張鈺1,劉忠華1,吳玉娥1,李文德2,閔凡貴1,劉香梅1
(1.廣東省實驗動物監(jiān)測所廣東省重點實驗室;2.廣東醫(yī)學院,湛江 524023;3.哈佛醫(yī)學院麻省總醫(yī)院放射腫瘤系)
目的比較了不同遺傳背景小鼠對禽流感H5N1亞型病毒的致病敏感性,為H5N1禽流感模型制作和機理研究提供依據(jù)。方法近交系BALB/c、C57BL/6和封閉群ICR、NIH Swiss和KM Swiss共五個不同品系小鼠。每個品系實驗動物30只,分接毒組20只,空白對照組10只,每組雌雄各半。病毒株為A/Goose/Guangdong/ NH/2003(H5N1),經(jīng)測定TCID50為10-4.875/mL。接毒組通過鼻腔接種0.1 mL病毒液,對照組接種正常雞胚尿囊液。小鼠接毒后連續(xù)觀察14 d,觀察記錄臨床癥狀、體溫、體重變化,對在實驗期間死亡和實驗14 d結束后仍然存活的小鼠均進行組織器官病理取材,進行RT-PCR病毒分離檢測、HE染色及H5N1抗原特異性免疫組化染色。結果①臨床癥狀:H5N1禽流感病毒能感染五個品系的小鼠,引起呼吸急促等癥狀和一過性體重、體溫下降。②死亡情況:小鼠在接毒后第1天即出現(xiàn)死亡,死亡的高峰期集中在接毒后第3~6天。五個品系小鼠死亡率存在差異,BALB/c為70%,ICR為50%,NIH Swiss為40%,C57BL/6為25%,KM Swiss為10%;③病毒分離:各組接毒小鼠在死亡后均進行了病毒分離,死亡小鼠的肺臟均分離到病毒,其他臟器未分離到病毒。④病理變化:實驗期間五個品系死亡小鼠肺臟病理改變相近。大體觀:死亡小鼠肺部淤血,呈暗紅色,體積增大,局部肺組織實變。鏡下觀:死亡小鼠的共同病理改變?yōu)殚g質性肺炎,具體表現(xiàn)為肺泡腔及間質出血、炎性細胞浸潤;間質增生,肺泡隔增寬;肺泡腔中見纖維素性滲出,透明膜形成。⑤免疫組化結果:在死亡小鼠的氣管上皮細胞和肺巨噬細胞可觀察到H5N1禽流感病毒陽性表達。結論小鼠作為H5N1禽流感病毒模型具有普適性,不同品系小鼠感染鵝源H5N1禽流感病毒的臨床癥狀、病程和病理變化與人禽流感病例相似。不同品系小鼠的死亡比例有明顯差別,可以根據(jù)不同的實驗目的,選擇不同品系的小鼠制作H5N1禽流感動物模型。不同品系的遺傳特性對禽流感易感性產生明顯的影響,遺傳背景可能與H5N1禽流感病毒感染應答機理存在聯(lián)系:BALB/c和C57BL/6均為近交系,其中BALB/c小鼠的品系特征之一表現(xiàn)為干擾素產量低,接種H5N1病毒后表現(xiàn)為高死亡率(70%),而C57BL/6小鼠的干擾素產量高,接種H5N1病毒后表現(xiàn)為低死亡率(25%),提示不同遺傳背景小鼠的干擾素水平與H5N1感染致死具相關性。為進一步研究H5N1禽流感病毒易感性相關基因以及其與宿主免疫反應的關系提供了一個研究基礎。
H5N1禽流感病毒;動物模型;小鼠
Data from the World Health Organization(WHO) has shown the mortality rate of human H5N1 virus infections to be greater than 50%[1].To better understand this viral disease,appropriate nonhuman mammalian models of H5N1 virus infection are needed. Recently,mice have been widely considered as one of the most useful mammalian models for studying pathogenesis and immunity to avian influenza A(H5N1)viruses due to their convenience of use and the virus’s predictability in this species[2-9].Our previous results also showed that the mouse is the most sensitive animal to H5N1 infection when compared with the rat,cavia cobaya,and gerbil in a pathogenicity study[10].It has been recently shown that host genetic background might strongly influence the response to a pathogen[11].However,little is known about how response to H5N1 virus infections varies among different strains of mice.Here,in order to provide more data to establish a suitable mammalian mouse model for H5N1 virus infection research,we report the results of a comparative study on the pathogenicity of avian influenza virus A/Goose/Guangdong/NH2003(H5N1)infection in five different genetic background strains of mice.
1 M aterials and M ethods
1.1 Virus
The avian influenza virus used in this study was A/Goose/Guangdong/NH/2003(H5N1)virus,which was isolated from a goose in the Guangdong Province of China in 2003.The original virus used in the experiment was generously provided by the Veterinary Research Institute of Southern China Agriculture University.Virus stocks were propagated in the allantoic cavity of 10-day-old specific pathogen-free (SPF)embryonated chicken eggs at 37℃.Infectious allantoic fluid was harvested and stored at-70℃until use for inoculation.The virus titer applied to the study was 104.87550%tissue culture infection dose (TCID50)/m L.All virus stocks were handled in the Animal Experiment Laboratory for Infectious Disease.
1.2 Anim als and virus in fections
A total of 150 SPF mice,18 to 22 g(male: female=1∶1),from 2 inbred strains(BALB/c and C57BL/6)and 3 outbreed stocks(ICR,NIH Swiss,and KM Swiss)(the Laboratory Animal Center of Southern Medical University,Guangzhou,China)were used.These mice were maintained in SPF conditions prior to infection.The absence of H5N1 virus-specific antibodies before infection in these m ice was confirmed by ELISA.All serological results from the tested mice (5%of all experimental m ice tested random ly)were negative.A ll m ice were moved to the Animal Experiment Laboratory for Infectious Disease 5 days prior to infection,where they were housed in microisolator cages within separated negative-pressure isolators.Thirty mice of each strain were subjected to an infected group of 20 mice that were inoculated with 0.1 m L(104.875TCID50)of A/Goose/Guangdong/NH/2003(H5N1)virus intra-nasally.The mock-infected control group(10 m ice)received an equivalent dilution of noninfectious allantoic fluid.All experiments were conducted in the Animal Experiment Laboratory for Infectious Disease.The mice had access to sterile food and water ad libitum.Animal care and use was approved by the Animal Care Comm ittee of the Guangdong Laboratory Animal Monitoring Institute (Guangzhou,China).
1.3 Observation of clinical signs
Clinical signs,including changes in activity,respiratory rate,body temperature and body weight,were assessed daily for 14 days post-infection.Body temperature was taken with a Braun IRT4520 ear thermometer(Braun Co.Germany).The number of mouse deaths was recorded daily and the mortality rates were compared among strains.
1.4 Necropsy and histopathological exam ination
Necropsy was performed on all mice that died during the experiment and those euthanized at the end of the study.Tissues from all major organs(including the lung,tonsil,heart,liver,spleen,kidney,stomach,intestine,and brain)were collected.Tissue samples were either stored at-70℃for viral isolation or fixed in formalin or polyoxymethylene for histopathological or immunohistochemical analysis.The gross lung lesions,quantified as a lung index,was determined for each mouse according to the following formula:Lung index(%)=lung weight(g)/body weight(g)x 100%.Tissue samples were fixed in 10%neutral buffered formalin,embedded in paraffin,cut at 5μm thickness,and stained with hematoxylin and eosin(H&E)for light m icroscopic exam ination.
1.5 Imm unohistochem istry
Tissue samples were fixed in 4% polyoxymethylene(pH 7.3)for 4 hours at 4℃,washed in phosphate buffered saline(PBS),cryoprotected in 30%sucrose overnight,and frozen in OCT embedding compound(Sakura Finetek U.S.A. Inc.Torrance,Calif.).Frozen sections were cut at 6 μm thickness by a Leica CM1900 freezing microtome (Leica Co.Germany).Sections were stained with an avidin-biotin comp lex(ABC)immunoperoxidase method(Wuhan Boshide Biotech Co.China),using a monoclonal antibody to the nucleoprotein of influenza virus A as the primary antibody(Biodesign International.Saco,ME).Under the light microscope,a visible diffuse or granular brown staining,usually seen darker in the nucleus than in the cytoplasm,was considered as positive staining.
1.6 V irus isolation
Tissue samples from all major organs were homogenized in sterile normal saline containing penicillin(200 U/m L)and streptomycin(200 mg/ m L)using Potter tissue grinders.The homogenate was centrifuged and the supernatant was inoculated into 10-day SPF embryonated chicken eggs.After incubation at 37℃for 48 hours,the identity of the virus from the embryos was confirmed with the viral hemagglutination test and/or hemagglutination inhibition test using standard procedures.
1.7 Statistical analysis
A ll data are expressed as mean±SD.Statistical differences between groups were tested by Student t test and Chi-square X2test using the SPSS statistical software package for Windows,version 11.0(SPSS,Inc.,Chicago,IL).A probability level of P<0.05 was considered to be statistically significant.
2 .Results
2.1 C linical signs
Most H5N1 virus-infected mice of all the five strains started to show inactivity,anorexia,lethargy,ruffled fur,hunched-back posture,and acute dyspnea at 24 hours post-infection.All these clinical signs became more severe from day 3 to day 7 post-infection. During this period,infected m ice exhibited severe respiratory distress,inappetence,and weight loss (Fig.1).The BALB/c and ICR m ice displayed more serious clinical signs than NIH Swiss,C57BL/6,and KM Swiss mice.Especially the KM Swiss mice only showed slight clinical signs.As shown in Figure 1,the difference in weight loss between the infected and uninfected control groups of all five mouse strains was significant by day 3 post-infection.The infected vs. control weights were:BALB/c:17.5±2.1 vs.22.9±1.8;ICR:17.5±1.0 vs.22.9±2.0;NIH Swiss: 16.8±2.0 vs.23.6±2.0;C57BL/6:17.2±2.3 vs. 23.1±1.5;and KM Swiss:16.9±1.3 vs.22.8± 1.7.The difference was significant(P<0.05)in all cases.Significant weight loss was continually observed in all strains of mice except the KM Swiss strain,in which the m ice gradually recovered to normal body weight after days 5 to 6 post-infection(Fig.1,Page 271).
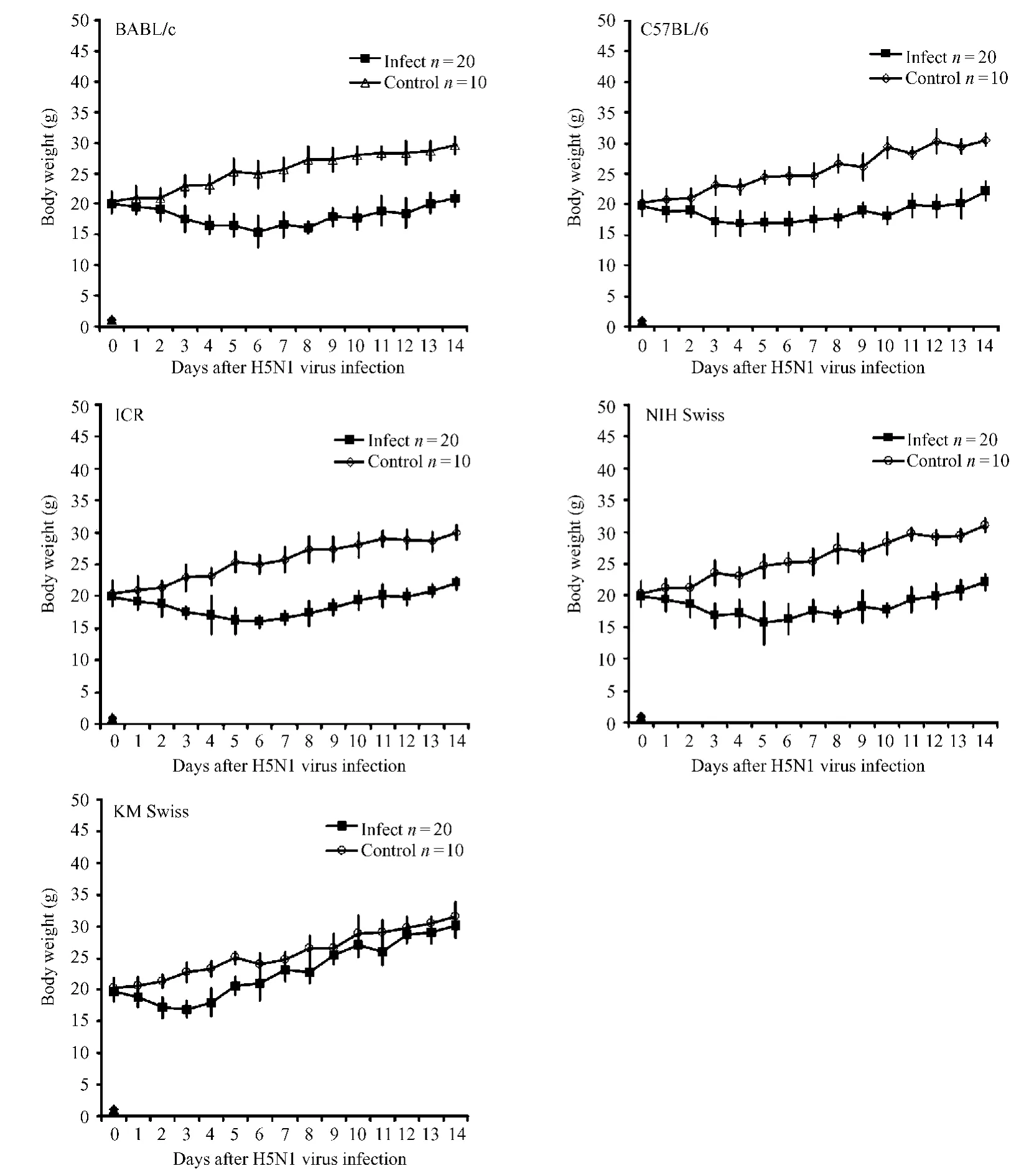
Fig.1 Curves show the changes in body weights of the five mouse strains after A/Goose/NH/2003(H5N1)virus infection
No fever developed in any H5N1 virus-infected mouse in our study;instead,the body temperature of all infected m ice,except the KM Swiss m ice,decreased significantly from about 37.4℃(±0.2)to below 34℃by days 3 to 8 in most strains(Fig.2).
About 80%(31 animals)of the in total 39 dead m ice,died between days 3 to 6 post-infection.Only 4 infected mice died before day 3,and 4 died after day 6 (Tab.1 and Fig.3).Figure 3 shows that the mortality rate varied drastically among the five strains of infected m ice:70%(14/20)in BALB/c,50% (10/20)in ICR,40%(8/20)in NIH Swiss,25% (5/20)in C57BL/6,and only 10%(2/20)in KM Swiss m ice.This H5N1 virus(A/Goose/Guangdong/NH/2003)demonstrated highest lethality for BALB/c m ice and led to significantly different mortality rates between BALB/c and all other tested mouse strains except the ICR(BALB vs.NIH Swiss,C57BL/6,or KM Swiss;P<0.05 or P<0.01,X2test).No mice in the mock-infected control group died during the study.
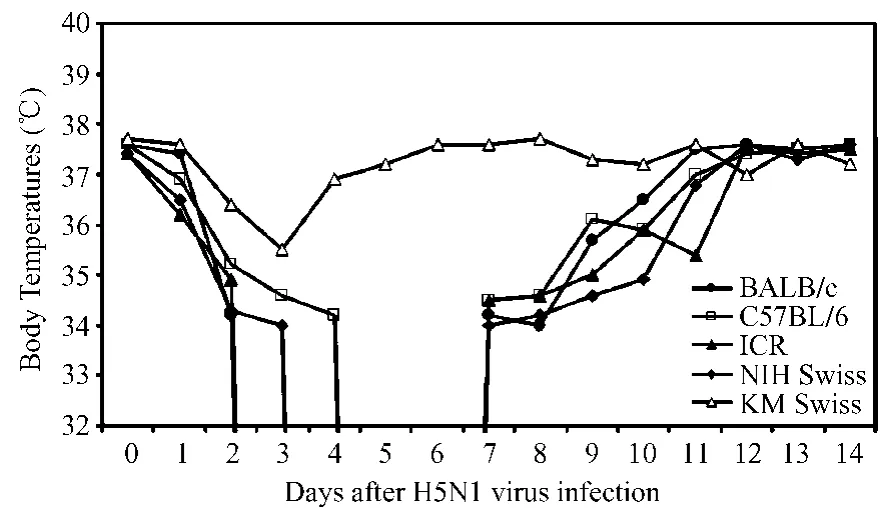
Fig.2 Curves show the changes in body temperature of the five mouse strains after A/Goose/ NH/2003(H5N1)virus-infection
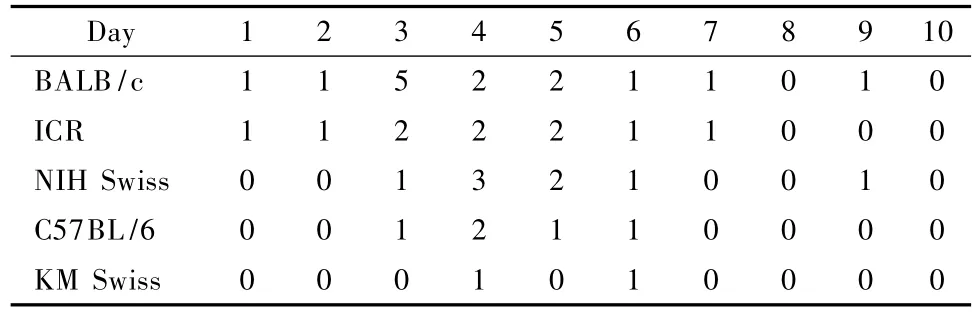
Tab.1 Number of deaths of mice in the five strains after H5N1 virus infection
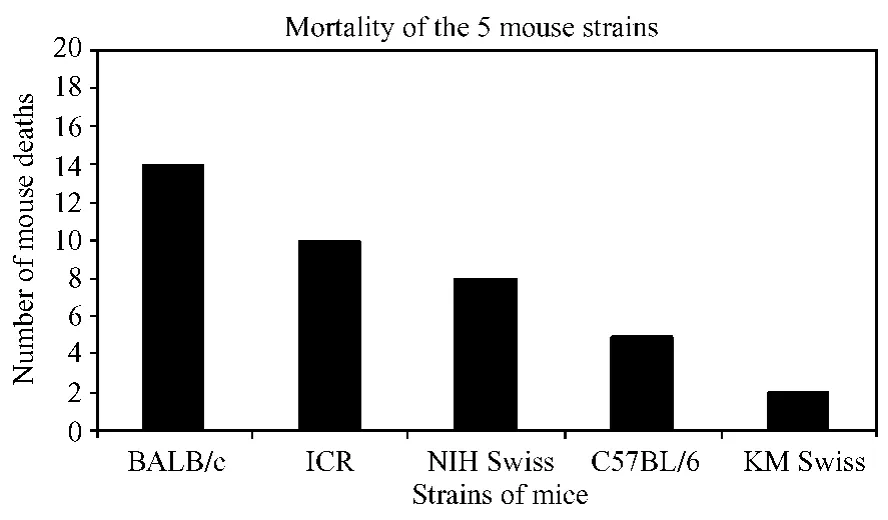
Fig.3 The mortality of the 5 mouse strains
2.2 Pathology
Gross lesions were observed mainly in the lungs of m ice from the infected groups,including pulmonary hyperem ia,hemorrhage,consolidation,and marked ly increased lung volume.Tab.2 shows the increased lung index in the H5N1 virus-infected mice.The lung indexes of the infected groups were significantly increased compared with those of their matched noninfected control groups in all the 5 strains(all P<0.01).The increase in the lung index usually correlated with highly edematous and hemorrhagic lungs.No gross lesions in the other major organs,such as the heart,liver and kidney,were observed in the infected mice.
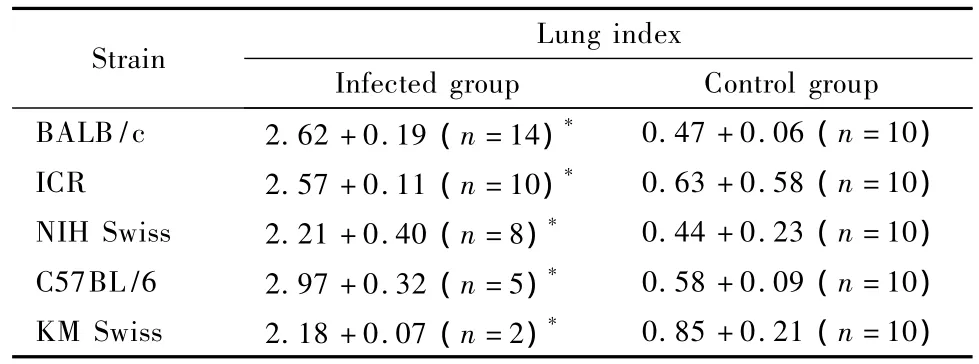
Tab.2 Lung index of the H5N1 virus-infected mice in the five strains(Mean±SD)
Under microscopic examination,necrotizing interstitial pneumonia was usually found in all cases of death in the infected groups.The pulmonary tissue exhibited hyperemia,hemorrhage,and inflammatory exudation,leading to consolidation.The lumina of alveoli and bronchioles were variably filled with protein-rich edema fluid,fibrin,erythrocytes and cell debris,adm ixed with many neutrophils,macrophages and lymphocytes(Fig.4).
2.3 V irus isolation and imm unohistochem ical findings
The virus was isolated from the lung tissues of all dead mice in the H5N1 virus-infected groups,but could not be detected in other organs.Positive immunohistochem ical staining was lim ited to the respiratory tract,such as epithelial cells,sloughed off epithelial cells,macrophages,neutrophils in the bronchial lumen and alveolar epithelial cells(Fig.5).
3 Discussion
Laboratories studying avian influenza have been in need of an ideal strain of virus that not only sensitively infects animal models to elicit pathological changes reflecting those of humans infected with H5N1 viruses,but also displays low pathogenicity towards human researchers.In our previous study,we used the A/ Goose/NH/2003(H5N1)virus isolated from a goose in our home province of Guangdong and successfully established the H5N1 virus infection mammalianmodels in mice,rats,and rhesus macaques[2,8,10,12]. Our results indicated that the A/Goose/NH/2003 (H5N1)virus meets the biological and high bio-safety standards that are valued in a suitable laboratory model.
In this study,we compared the pathogenicity of the H5N1 virus infection in 5 different strains of m ice,including 2 inbred strains(BALB/c and C57BL/6) and 3 outbred stocks(ICR,NIH Swiss,and KM Swiss).Our results showed that an A/Goose/NH/ 2003(H5N1)virus infection inoculated by 0.1× 104.875TCID50of virus intra-nasally can induce respiratory illness in all 5 strains of m ice and disp lay severe to minor clinical signs,including acute respiratory distress symptom(ARDS),and differences in lethality.As the results showed in Table 1,31 infected mice(about 80%of the total number of dead m ice)died between days 3 to 6 post-infection. Necropsy showed that all infected m ice had highly edematous and hemorrhagic lungs,and a significantly increased lung index when compared with that of corresponding non-infected control m ice(Table 2,all P<0.01).Histopathological examination showed that the lung lesions of infected mice were usually necrotizing interstitial pneumonia,characterized by inflammatory cell infiltration,interstitial hyperemia,and alveolar and bronchiolar edema and hemorrhage. In addition,some of their epithelial cells showed degenerative or necrotic characteristics.All these pathological changes seen in the present study are sim ilar to those observed in a previous study that reported ARDS in a BALB/c mouse model induced by an A/Chicken/Hebei/108/2002(H5N1)virus[7]. Furthermore,the results of our study demonstrate the efficiency with which H5N1 virus rep licates in the respiratory tracts of infected mice,especially BALB/c mice(confirmed by virus isolation and immunohistochem ical staining).Infected m ice could also represent most of the clinical symptoms and pathological changes found in humans infected with H5N1 viruses[1].In total,these results combined to confirm the mouse H5N1 virus infection as a suitable model for this viral disease studies.Moreover,our study clearly showed that the mortality rate was diverse in these 5 strains of m ice(from 10%to 70%).Thus,our research acts to further refine mouse models studying H5N1 virus infections by allowing the selection of mouse strains most relevant to the purpose under investigation.
It has been previously demonstrated that host genetic background can strongly influence the response to pathogens.Comparing seven inbred laboratory strains of mice to influenza A(H1N1&H7N7)virus infection,Srivastava showed that two strains of mice in particular,DBA/2J and A/J,had a very high susceptibility to their viral infections relative to the other strains[11].Their findings indicated that the genetic background of the host contributes significantly to influenza A virus infection.However,little was known about the influence of the genetic background to avian influenza A H5N1 virus in particular.Our current study showed that the mortality rate was diverse in infected BALB/c(70%),ICR(50%),NIH Swiss(40%),C57BL/6(25%),and KM Swiss (10%)mice.These results reiterate the importance of host genetic background and provide an increased understanding of this notion specifically for avian influenza A(H5N1)virus.Furthermore,both BALB/ c and C57BL/6 inbred mice strains produce interferon (IFN)at strain characteristic levels:BALB/c mice with a lower level and C57BL/6 m ice with a higher level[13].It is well known that IFN is a broad-spectrum antiviral protein.Our mortality results match with what would be expected based on IFN levels:BALB/c (70%)has far higher IFN than C57BL/6(25%). Among the three outbred stocks(ICR,NIH Swiss,and KM Swiss),the mortality rate of infected KM Swiss was the lowest(only 10%).The low mortality rate might be connected with the strain characteristics of KM Swiss,which has higher reproductivity,adaptability,and ability to resist disease[13]. Therefore,our study provides a foundation for further research to elucidate how the genetic sensitivity and host immune-response of mice relate to avian influenza A H5N1 virus infection.
Acknow ledgm ents
This study was supported by grants 2005B20201019 and 2004B20201028 from the Agricultural Science&Technology Project of Guangdong Province,and grants 06023011 from the Natural Science Project of Guangdong Province.The authors thank Dr.Ming Liao for generously providing the A/Goose/Guangdong/NH/2003(H5N1)virus. Dr.Peigen Huang is a visiting professor at Guangdong Laboratory Animals Monitoring Institute.
(Fig.4,F(xiàn)ig.5 on colur page 1).
Reference
[1]Thomas JK,Noppenberger J.Avian influenza:A review.Am J Health-Syst Pharm.2007,64:149-165.
[2]李志東,劉忠華,劉香梅,等.禽流感H5N1亞型病毒感染ICR小鼠的動物模型[J].中國實驗動物學報,2008,16(2): 91-94.
[3]Bright RA,Cho DS,Rowe T,et al.Mechanisms of pathogenicity of influenza A(H5N1)viruses in mice.Avian Dis.2003,47: 1131-1134.
[4]Dybing JK,Schultz-Cherry S,Swayne DE,et al.Distinct pathogenesis of Hong Kong-origin H5N1 viruses in mice compared to that of other highly pathogenic H5 avian influenza viruses.J Virol.2000,74:1443-1450.
[5]Lu X,Tumpey TM,Morken T,et al.A mouse model for the evaluation of pathgenesis and immunity to influenza A(H5N1) viruses isolated from humans.J Virol.1999,73:5903-5911.
[6]O’Neill E,Krauss SL,Riberdy JM,et al.Heterologous protection against lethal A/HongKong/156/97(H5N1)influenza virus infection in C57BL/6 mice.J Gen Virol.2000,81:2689-2696.
[7]Xu T,Qiao J,Zhao L,et al.Acute respiratory distress syndrome induced by avian influenza A(H5N1)virus in mice.Am J Respir Crit Care Med.2006,174:1011-1017.
[8]閔凡貴,張鈺,趙維波,等.H5N1流感病毒不同途徑接種BALB/c小鼠臨床病理學比較[J].廣東醫(yī)學,2007,28(2): 192-194.
[9]Sidwell RW.The mouse model of influenza virus infection.In: Zak O,M.A.Sande MA(edited):Handbook of animal models of infection.New York,NY Academic Press,Inc.1999,p981-987.
[10]李志東,劉忠華,陳美才,等.小鼠、大鼠、豚鼠和沙鼠對禽流感病毒致病敏感性的比較初報[J].中國實驗動物學報,2004,12(4):212-214.
[11]Srivastava B,Blazejewska P,Hessmann M,et al.Host genetic background strongly influences the response to influenza A virus infections.PLoS One.2009,4(3):e4857.
[12]Huang R,Deng SC,Wu YE,et al.A nonhuman primate model for evaluation of pathogenesis and immunity to avian influenza A (H5N1)virus.J AALA.2007,46(4):97.
[13]孫靖.實驗動物學基礎[M].北京:科學技術出版社,2005,50.
Com parison of the Pathogenicity of Avian Influenza A(H 5N1) Virus in Five Strains of M ice
HUANG Ren1,WANG Ying-yan1,DENG Shao-chang1,HUANG Pei-gen3,ZHANG Yu1,LIU Zhong-h(huán)ua1,WU Yu-e1,LI Wen-de2,M IN Fan-gui1,LIU Xiang-mei1
(1.Key Animal Experiment Laboratory for Guangdong Province,Guangdong Laboratory Animal Monitoring Institute,Guangzhou,510260,China;2.Guangdong Medical College,Zhanjiang,524023;3.Department of Radiation Oncology,Massachusetts General Hospital,Harvard Medical School,Boston,MA 02114,USA)
ObjectiveTo test our hypothesis that sensitivity to avian influenza A(H5N1)virus varies amongmouse strain backgrounds,we compared the pathogenicity of H5N1 viral infection in 5 mouse strains.M ethods Onehundred-fifty mice from 2 inbred strains(BALB/c and C57BL/6),and 3 outbred stocks(ICR,NIH Swiss,and KM Swiss)were used.Thirty mice of each strain were subjected to an infected group(20 m ice),in which mice were inoculated with 0.1 m L(104.875TCID50)of A/Goose/Guangdong/NH/2003(H5N1)virus intra-nasally;ten control m ice received noninfectious allantoic fluid.Clinical signs were assessed daily for 14 days post-infection.Necropsy was performed on m ice that died during the experiment and those euthanized at end of study.Tissue samp les were collected for viral isolation and pathological analysis.Results H5N1 virus infection can cause respiratory illness in all 5 strains with severe or m inor acute respiratory distress symptoms,but with different mortality rates:70%in BALB/c;50%in ICR;40%in NIH Swiss;25%in C57BL/6;and 10%in KM Swiss mice.Necrotizing interstitial pneumonia was found in all cases of death.The virus was isolated from the lungs of all infected dead mice.ConlusionA/Goose/Guangdong/NH/2003 (H5N1)virus can infect all mouse strains used in this study,and can cause clinical symptoms and pathological changes similar to those found in humans infected with H5N1 viruses.However,the pathogenicity of H5N1 viral infection varies significantly between the different mouse strains.Thus,in future study of H5N1 virus infections the mouse strain most relevant to their particular research purpose should be selected as animal model.
H5N1 avian influenza virus;Animal models;M ice
R373.13
A
1005-4847(2010)04-0271-06
2010-01-06
黃韌,廣州市海珠區(qū)新港西路105號,電話:020-34063369,E-mail:labking@sohu.com。黃培根,哈佛醫(yī)學院麻省總醫(yī)院放射腫瘤系,E-mail:peigen@steele.mgh.harvard.edu
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
學苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
云南中醫(yī)學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
