東太平洋表層海水中添加氮、磷的現(xiàn)場(chǎng)培養(yǎng)實(shí)驗(yàn)
2010-09-11 09:50:02唐松
海洋通報(bào) 2010年4期
關(guān)鍵詞:生長(zhǎng)實(shí)驗(yàn)
唐 松
(中國(guó)科學(xué)院廣州地球化學(xué)研究所,廣東 廣州 510640)
東太平洋表層海水中添加氮、磷的現(xiàn)場(chǎng)培養(yǎng)實(shí)驗(yàn)
唐 松
(中國(guó)科學(xué)院廣州地球化學(xué)研究所,廣東 廣州 510640)
借助“中國(guó)首次環(huán)球科學(xué)考察”航次,在東太平洋表層海水進(jìn)行了添加氮、磷的現(xiàn)場(chǎng)培養(yǎng)實(shí)驗(yàn)。現(xiàn)場(chǎng)記錄了實(shí)驗(yàn)水體溫度的變化,用分光光度法對(duì)水體硝酸氮和活性磷酸鹽濃度進(jìn)行了檢測(cè),并用熒光法分析了水體葉綠素a濃度。結(jié)果表明,氮的添加會(huì)引起水體中葉綠素a濃度短期內(nèi)快速增大,同時(shí)伴隨硝酸鹽濃度的顯著降低,而單獨(dú)添加P對(duì)水體中葉綠素a濃度影響并不顯著;水體中N/P比值與葉綠素a濃度、N/P比值與浮游植物生長(zhǎng)速度、溫度與葉綠素a濃度以及溫度與浮游植物生長(zhǎng)速度之間均缺乏相關(guān)性。因此認(rèn)為,在東太平洋實(shí)驗(yàn)海區(qū)表層海水中添加氮會(huì)引起浮游植物快速爆發(fā),而磷的添加并不能刺激浮游植物快速生長(zhǎng),水體 N/P比值和水體溫度都不能單獨(dú)控制浮游植物群落的生長(zhǎng)。
東太平洋;培養(yǎng)實(shí)驗(yàn);營(yíng)養(yǎng)鹽;浮游植物
Abstract:Nutrient enrichment experiment was carried out in the East Pacific Ocean during the Chinese First Around-world Researching Cruise,with the addition of nitrogen or phosphorus to surface seawater.The variation of water temperature was recorded.The nutrient concentration was analyzed with spectrophotometry,and chlorophyll a concentration with fluorescence analysis.The addition of nitrogen resulted in the rapid increase of chlorophyll a concentration with the depletion of nitrogen in the experimental water,but no apparent variation occurred in chlorophyll a concentration when phosphate was added.The correlation was absent between the ratio of nitrogen to phosphorus and chlorophyll a concentration,the ratio of nitrogen to phosphorus and phytoplankton growing rate,water temperature and chlorophyll a concentration,and water temperature and phytoplankton growing rate.So it was considered that addition of nitrogen could stimulate phytoplankton bloom in the surface seawater in the East Pacific Ocean,while addition of phosphate could not.In addition,the ratio of nitrogen to phosphorus and the water temperature cannot control the growth of phytoplankton community.
Keywords:East Pacific Ocean; enrichment experiment; nutrient; phytoplankton
長(zhǎng)期以來,營(yíng)養(yǎng)鹽限制作用作為影響浮游植物生長(zhǎng)最為重要的因素之一,一直是海洋生態(tài)學(xué)研究的重要領(lǐng)域。目前已經(jīng)認(rèn)識(shí)到,在多數(shù)開闊大洋,浮游植物的生長(zhǎng)主要受表層海水中可利用的氮和磷濃度限制[1]。但浮游植物生長(zhǎng)到底是氮限制還是磷限制,目前還沒有形成統(tǒng)一觀點(diǎn)。在東熱帶太平洋海區(qū)的貧營(yíng)養(yǎng)區(qū)域[2-5],貧營(yíng)養(yǎng)的南太平洋亞熱帶海區(qū)[6,7],以及南海[8]的培養(yǎng)實(shí)驗(yàn)結(jié)果都表明,氮是這些海區(qū)最重要的限制營(yíng)養(yǎng)鹽。此外,美國(guó)北卡羅來納州河口[9]、夏威夷Kaneohe海灣[10]、菲律賓西北部海域[11]等也都為氮限制。而在地中海[12]、長(zhǎng)江口[13,14]、東海[15],以及萊州灣附近海域[16]等,則表現(xiàn)出磷限制。我國(guó)大亞灣海域[17,18]、臺(tái)灣海域上升流區(qū)[19]以及東、黃海典型海區(qū)[20]的添加營(yíng)養(yǎng)鹽培養(yǎng)實(shí)驗(yàn)表明,氮和磷對(duì)這些海區(qū)浮游植物生長(zhǎng)都有限制作用。
可以看出,在不同海域氮和磷對(duì)浮游植物生長(zhǎng)的限制作用也不同。而且,目前國(guó)內(nèi)關(guān)于開闊大洋氮和磷對(duì)浮游植物生長(zhǎng)的限制作用報(bào)道較少。借助2005年“中國(guó)首次環(huán)球科學(xué)考察”航次,筆者利用東太平洋表層海水進(jìn)行了添加氮、磷的現(xiàn)場(chǎng)培養(yǎng)實(shí)驗(yàn),這為研究該海區(qū)的營(yíng)養(yǎng)鹽限制因子提供了寶貴的機(jī)會(huì)。本文通過對(duì)實(shí)驗(yàn)過程中水體溫度、營(yíng)養(yǎng)鹽濃度、葉綠素a濃度等參數(shù)進(jìn)行分析,探討溫度、營(yíng)養(yǎng)鹽氮和磷的添加,以及N/P比值等對(duì)東太平洋實(shí)驗(yàn)海區(qū)浮游植物生長(zhǎng)的影響。
1 材料與方法
2005年9月,借助“中國(guó)首次環(huán)球科學(xué)考察航次”,利用東太平洋表層海水(102°37.94’W,3°15.10’S)進(jìn)行了現(xiàn)場(chǎng)添加N、P的現(xiàn)場(chǎng)培養(yǎng)實(shí)驗(yàn)。實(shí)驗(yàn)在3個(gè)500 L的丙烯酸培養(yǎng)桶中進(jìn)行,并配備一套冷卻裝置可控制水體溫度,有關(guān)實(shí)驗(yàn)裝置及其操作的詳細(xì)介紹見參考文獻(xiàn)[21]。1號(hào)桶為采樣點(diǎn)的表層海水,2號(hào)和3號(hào)培養(yǎng)桶在采取的表層海水中分別添加了 0.01mol的 N(KNO3)和 0.01mol的 P(KH2PO4),實(shí)驗(yàn)過程持續(xù)了13天。實(shí)驗(yàn)海區(qū)表層海水營(yíng)養(yǎng)鹽背景值見表1。
實(shí)驗(yàn)過程中每間隔12 h或24 h采樣一次,記錄水體溫度。利用7230 G分光光度計(jì)現(xiàn)場(chǎng)分析水體中的硝酸氮、活性磷酸鹽濃度,同時(shí)用熒光法分析水體葉綠素a濃度。具體實(shí)驗(yàn)流程及營(yíng)養(yǎng)鹽和葉綠素a濃度分析方法參見參考文獻(xiàn)[21]。

表1 東太平洋實(shí)驗(yàn)海水背景值Tab.1 Background value of the experimental water in the East Pacific Ocean
2 結(jié)果與討論
2.1 營(yíng)養(yǎng)鹽的添加對(duì)浮游植物生長(zhǎng)的影響
葉綠素a濃度是反應(yīng)浮游植物生長(zhǎng)最直接、準(zhǔn)確的參數(shù),同時(shí),其變化速度反應(yīng)了浮游植物的生長(zhǎng)速度。
實(shí)驗(yàn)過程中水體葉綠素a濃度變化見圖1。整個(gè)實(shí)驗(yàn)過程中1、2、3號(hào)培養(yǎng)桶中葉綠素a濃度平均值分別為0.68 mg/m3,1.97 mg/m3和0.83 mg/m3。由圖1可以看出,各培養(yǎng)桶水體中葉綠素a濃度的相對(duì)快速增大發(fā)生在實(shí)驗(yàn)的第2-5天。其中,2號(hào)加N桶中葉綠素a濃度增大最為顯著,在實(shí)驗(yàn)第5天達(dá)到最高值6.69 mg/m3,高出初始值(0.16 mg/m3)40多倍。1號(hào)空白桶和3號(hào)加P培養(yǎng)桶水體中葉綠素a濃度的最高值分別出現(xiàn)在實(shí)驗(yàn)第5天和第4天,值分別為1.51 mg/m3和1.92 mg/m3,比水體初始值高出8倍和9倍。培養(yǎng)實(shí)驗(yàn)第5天后,三個(gè)培養(yǎng)桶中的葉綠素a濃度均下降,至實(shí)驗(yàn)結(jié)束時(shí),分別降至0.65 mg/m3,0.65 mg/m3和0.45 mg/m3。不難看出,在東太平洋表層海水中添加N對(duì)浮游植物生長(zhǎng)有明顯促進(jìn)作用。雖然添加P的3號(hào)桶中葉綠素a濃度也發(fā)生較明顯增大,但與1號(hào)空白桶相比,無論其變化趨勢(shì),還是平均值或者最高值,都非常接近。因此,筆者認(rèn)為在該海區(qū)單獨(dú)添加P對(duì)刺激浮游植物爆發(fā)并沒有明顯作用。
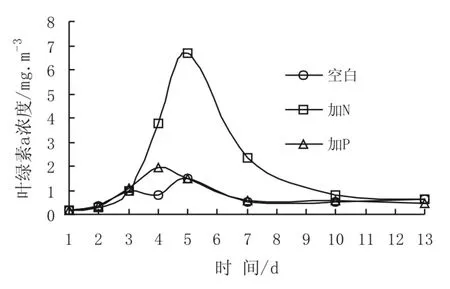
圖1 實(shí)驗(yàn)水體中葉綠素a濃度變化Fig.1 Variation of Chl-a concentration in the experimental water
除了浮游植物生長(zhǎng)量,其生長(zhǎng)速率的變化也是判斷浮游植物生長(zhǎng)狀態(tài)的重要參數(shù)。本文根據(jù)下面的公式計(jì)算浮游植物生長(zhǎng) 速率(R):
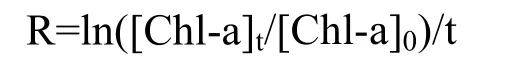
式中:t代表實(shí)驗(yàn)時(shí)間,[Chl-a]t和[Chl-a]0分別代表了在實(shí)驗(yàn)時(shí)間t時(shí)和初始時(shí)的葉綠素a濃度。
1-3號(hào)培養(yǎng)桶水體中平均R值分別為0.44/d、0.58/d和0.46/d。圖2顯示了東太平洋海水培養(yǎng)實(shí)驗(yàn)中浮游植物生長(zhǎng)速度的變化。可以看出,三個(gè)培養(yǎng)桶中R值變化趨勢(shì)基本一致,在實(shí)驗(yàn)過程前期的增大后,都表現(xiàn)出降低的趨勢(shì)。1號(hào)和3號(hào)桶中浮游植物生長(zhǎng)速度在實(shí)驗(yàn)第3天分別增至最大值0.90/d和0.87 /d。2號(hào)桶中R值的最大值出現(xiàn)在實(shí)驗(yàn)的第4天,為1.03 /d。各培養(yǎng)桶中R值在出現(xiàn)最高值后,均保持下降趨勢(shì)。實(shí)驗(yàn)結(jié)束時(shí),1-3號(hào)培養(yǎng)桶中的R值分別為0.12 /d 、0.11 /d 和0.07 /d。可見,浮游植物生長(zhǎng)速度的變化也表明,2號(hào)桶中N的添加對(duì)實(shí)驗(yàn)水體中浮游植物生長(zhǎng)影響最明顯。3號(hào)桶中R值的變化趨勢(shì)、平均值和絕對(duì)值均接近于1號(hào)桶,因此認(rèn)為,P的添加對(duì)浮游植物的生長(zhǎng)并沒有顯著影響。
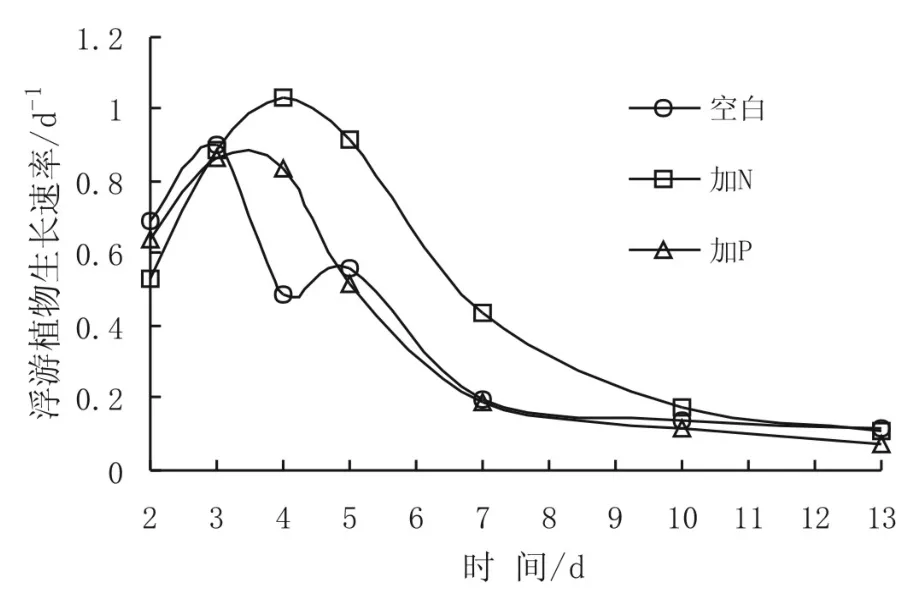
圖2 實(shí)驗(yàn)水體中浮游植物生長(zhǎng)速度變化Fig.2 Variation of phytoplankton growth rate in the experimental water
2.2 溫度對(duì)浮游植物生長(zhǎng)的影響
東太平洋海水培養(yǎng)實(shí)驗(yàn)過程中溫度變化見圖3。整個(gè)實(shí)驗(yàn)過程平均溫度為22.8 ℃,變化范圍為21~26 ℃。在實(shí)驗(yàn)的前10天,溫度曲線表現(xiàn)出鋸齒狀的變化特征,此后溫度一直升高,最高值出現(xiàn)在實(shí)驗(yàn)結(jié)束時(shí),達(dá)到26 ℃。雖然已有研究表明,水體溫度對(duì)硅藻的生長(zhǎng)有顯著影響[22-24],但由于不同浮游植物種類適宜的生存溫度有所差異,水體溫度對(duì)整個(gè)浮游植物群落的影響要遠(yuǎn)遠(yuǎn)小于對(duì)具體種的影響[25]。在東太平洋培養(yǎng)實(shí)驗(yàn)過程中,各培養(yǎng)桶浮游植物相對(duì)快速生長(zhǎng)時(shí),水體溫度并沒有表現(xiàn)出明顯的規(guī)律性變化與之對(duì)應(yīng),而且水體溫度與葉綠素a濃度以及R值之間都缺乏明顯相關(guān)性(表2)。因此認(rèn)為,至少在該實(shí)驗(yàn)過程中,水體溫度并不能單獨(dú)主控整個(gè)浮游植物群落的生長(zhǎng)量,關(guān)于其對(duì)具體浮游植物種類的影響還有待深入研究。
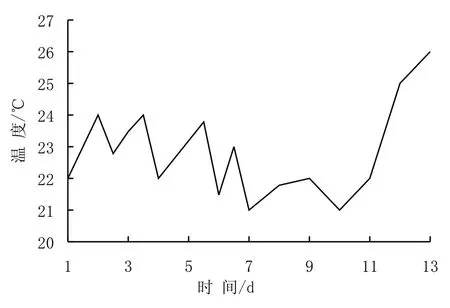
圖3 實(shí)驗(yàn)水體中溫度的變化Fig.3 Variation of temperature in experimental water

表2 溫度與Chl-a濃度以及R值的相關(guān)性分析結(jié)果Tab.2 Result of correlation analysis between temperature,Chl-a and the value of R
2.3 實(shí)驗(yàn)水體中的營(yíng)養(yǎng)鹽濃度的變化
圖4顯示了實(shí)驗(yàn)過程中水體硝酸鹽和磷酸鹽濃度的變化。與各培養(yǎng)桶水體中葉綠素a濃度在實(shí)驗(yàn)第2-5天的明顯增長(zhǎng)相對(duì)應(yīng),1號(hào)和3號(hào)培養(yǎng)桶中水體硝酸鹽濃度分別由第 2天的 6.27 μM 和3.82 μM 降低至第 4天的 0.16 μM 和 0.13 μM,2號(hào)加 N桶中的硝酸鹽濃度則在實(shí)驗(yàn)第 2-6天快速下降,由32.65 μM降低至3.95 μM。很明顯,浮游植物的生長(zhǎng)與水體中硝酸鹽的消耗存在密切聯(lián)系,而且,N的添加可促進(jìn)浮游植物快速爆發(fā)。同時(shí),伴隨著水體中硝酸鹽濃度的降低,1號(hào)和2號(hào)培養(yǎng)桶中磷酸鹽濃度分別由初始的1.00 μM和0.93 μM降低至實(shí)驗(yàn)第4天的0.07 μM和0.26 μM,3號(hào)培養(yǎng)桶中磷酸鹽濃度初始值為18.61 μM,至實(shí)驗(yàn)第5天降為17.82 μM。這說明在浮游植物生長(zhǎng)消耗水體硝酸鹽的同時(shí),也利用了其中的磷酸鹽。此外,雖然 3號(hào)培養(yǎng)桶中的平均磷酸鹽濃度高出1號(hào)和2號(hào)培養(yǎng)桶50多倍,但該桶中浮游植物的生長(zhǎng)量?jī)H接近于1號(hào)空白桶(見圖),而且其中磷酸鹽濃度的消耗也與其它兩個(gè)培養(yǎng)桶中相近。這說明,磷酸鹽并不是實(shí)驗(yàn)海區(qū)表層海水中浮游植物生長(zhǎng)的主要限制營(yíng)養(yǎng)鹽。
此次培養(yǎng)實(shí)驗(yàn)中,三個(gè)培養(yǎng)桶中浮游植物生長(zhǎng)速度變化趨勢(shì)基本一致。在實(shí)驗(yàn)第3天前,1號(hào)和3號(hào)培養(yǎng)桶中R值持續(xù)增大,而2號(hào)桶中浮游植物生長(zhǎng)速度的升高發(fā)生在實(shí)驗(yàn)前4天。與R值的增大對(duì)應(yīng),各培養(yǎng)桶水體中的硝酸鹽濃度都發(fā)生降低,尤其2號(hào)加N桶中表現(xiàn)最明顯,硝酸鹽濃度在實(shí)驗(yàn)第5天比初始值降低了18.22 μM(見圖4)。這說明N的消耗對(duì)于浮游植物生長(zhǎng)有明顯的促進(jìn)作用。在R值增大期,雖然各培養(yǎng)桶水體中磷酸鹽濃度在硝酸鹽濃度降低的同時(shí)也有所減小,但在3號(hào)加P培養(yǎng)桶中,R值快速增大至峰值時(shí),水體磷酸鹽濃度僅減小了0.79 μM,與2號(hào)桶和3號(hào)桶中磷酸鹽濃度的降幅接近。因此認(rèn)為,實(shí)驗(yàn)海區(qū)海水中浮游植物不能單獨(dú)利用P促進(jìn)自身快速生長(zhǎng)。
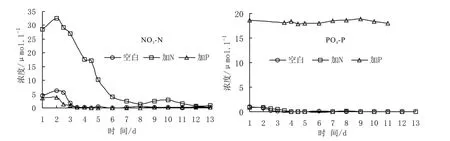
圖4 實(shí)驗(yàn)水體中營(yíng)養(yǎng)鹽濃度變化Fig.4 Variation of nutrient concentration in experimental water
2.4 N/P比值與浮游植物生長(zhǎng)的關(guān)系
圖5顯示了此次培養(yǎng)實(shí)驗(yàn)過程中水體N/P值的變化。考慮到1號(hào)培養(yǎng)桶中水體營(yíng)養(yǎng)鹽濃度偏低,實(shí)驗(yàn)誤差對(duì)N/P比值的影響較大,在此不做討論。2號(hào)、3號(hào)培養(yǎng)桶中水體平均N/P值分別為94.98和0.14。在實(shí)驗(yàn)第 6天之前,2號(hào)培養(yǎng)桶水體以高氮為特征(見圖4),平均硝酸鹽和磷酸鹽濃度分別為21.61 μM和0.34 μM。在浮游植物爆發(fā)時(shí),N/P比值由初始值 32.29突增至實(shí)驗(yàn)第 4天的 385.60。3號(hào)培養(yǎng)桶水體始終以高磷為特征(見圖4),硝酸鹽和磷酸鹽的平均濃度分別為0.69 μM和17.42 μM。在浮游植物相對(duì)快速生長(zhǎng)期內(nèi),水體N/P比值由實(shí)驗(yàn)第2天的0.44下降至第4天的0.07。可見,在2號(hào)和3號(hào)培養(yǎng)桶水體中浮游植物相對(duì)快速生長(zhǎng)時(shí),N/P比值的變化規(guī)律并不統(tǒng)一。已有研究表明,水體中N/P比值與浮游植物生長(zhǎng)特性有很大關(guān)系[26],而且不同種類浮游植物生長(zhǎng)的最佳營(yíng)養(yǎng)鹽條件對(duì)N/P比值的要求也不同[27-29]。由于添加營(yíng)養(yǎng)鹽不同,2號(hào)和3號(hào)培養(yǎng)桶中浮游植物組成肯定不同。正是由于滿足這些浮游植物有利生長(zhǎng)的N/P比值要求不同,造成在2號(hào)和3號(hào)培養(yǎng)桶中葉綠素a濃度相對(duì)快速增大時(shí),水體N/P比值變化表現(xiàn)出不同特征。另外,對(duì)實(shí)驗(yàn)過程中2號(hào)和3號(hào)培養(yǎng)桶水體N/P比值和Chl-a濃度以及R值進(jìn)行相關(guān)性分析的結(jié)果(表3)表明,水體中的 N/P比值同浮游植物生長(zhǎng)量和生長(zhǎng)速度之間無明顯相關(guān)性。這說明東太平洋實(shí)驗(yàn)海區(qū)水體中的N/P比值不能單獨(dú)控制浮游植物群落的整體生長(zhǎng)特性。

圖5 實(shí)驗(yàn)水體中N/P值變化Fig.5 Variation of the ratio of N to P in experimental water
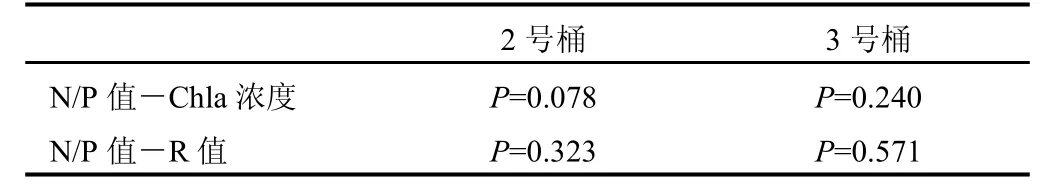
表3 N/P比值與Chl-a濃度以及R值的相關(guān)性分析結(jié)果Table.3 Result of correlation analysis between the ratio of N to P ,Chl-a and the value of R
此外,在高氮的2號(hào)培養(yǎng)桶中,無論是整個(gè)實(shí)驗(yàn)過程的平均N/P值,還是浮游植物快速生長(zhǎng)時(shí)的N/P平均值都遠(yuǎn)大于16。而在高磷的3號(hào)桶中,水體N/P比值則一直遠(yuǎn)小于16。根據(jù)Redfield定律,2號(hào)培養(yǎng)桶中浮游植物生長(zhǎng)應(yīng)受P限制,3號(hào)桶中則為N限制。而前面的分析已表明,在2號(hào)桶中,浮游植物生長(zhǎng)與水體硝酸鹽濃度的降低有明顯的對(duì)應(yīng)關(guān)系;而在3號(hào)桶中,水體中硝酸鹽濃度在實(shí)驗(yàn)第3天就降低至接近表層海水的水平,浮游植物生長(zhǎng)也沒有表現(xiàn)出受N限制的影響。可見,Redfield定律有其一定的適用范圍,水體N/P比值并不能單獨(dú)決定浮游植物群落生長(zhǎng)的營(yíng)養(yǎng)鹽限制因子。
3 結(jié) 論
利用東太平洋表層海水進(jìn)行了添加氮、磷的現(xiàn)場(chǎng)培養(yǎng)實(shí)驗(yàn),通過對(duì)水體溫度、營(yíng)養(yǎng)鹽濃度和葉綠素a濃度等參數(shù)分析認(rèn)為:
a)至少在該實(shí)驗(yàn)過程中,水體溫度不能單獨(dú)控制浮游植物群落生長(zhǎng);
b)實(shí)驗(yàn)海區(qū)水體中浮游植物生長(zhǎng)表現(xiàn)出明顯的N限制作用,單獨(dú)添加P并不能刺激浮游植物短期內(nèi)快速爆發(fā);
c)水體中的N/P比值既不能控制游植物群落的整體生長(zhǎng)特性,也不能決定其限制因子。
致謝:感謝彭曉彤老師、林榮澄老師、蔣磊和吳自軍博士,以及“大洋一號(hào)”船員在現(xiàn)場(chǎng)實(shí)驗(yàn)過程中給予的大力支持和幫助!
[1]Gruber N.The dynamics of the marine nitrogen cycle and its influence on atmospheric CO2 variations [A].The ocean carbon and climate [C].Dordrecht: Kluwer Academic,2005: 97.
[2]Thomas W H.Surface nitrogenous and phytoplankton in the northeastern tropical Pacific Ocean [J].Limnol Oceanogr,1966,11:393-400.
[3]Thomas W H.The nitrogen nutrition of phytoplankton in the northeastern tropical Pacific Ocean [J].Stud Trop Oceanogr Miami,1967,5: 280-289.
[4]Thomas W H.Phytoplankton nutrient enrichment esperiments off Baja California and in the eastern equatorial Pacific Ocean [J].J Fish Res Bd Can,1969,26: 1133-1145.
[5]Thomas E.Effect of ammonium and nitrate concentration on chlorophyll increases in natural tropical pacific phytoplankton populations [J].Limnol Oceanogr,1970,15: 386-394.
[6]Dufour P,Charpy L,Bonnet S,et al.Phytoplankton nutrient control in the oligotrophic South Pacific subtropical gyre [J].Mar Ecol Prog Ser,1999,179: 285-290.
[7]Dufour P,Berland B.Nutrient control of phytoplankton biomass in atollagoons and Pacific Ocean waters: Studies with factorial enrichment bioassays [J].J Exp Mar Bio Ecol,1999,234: 147-166.
[8]Chen Y L,Chen H C,Karl D M,et al.Nitrogen modulates phytoplankton growth in spring in the South China Sea [J].Continental Shelf Research,2004,24: 527-541.
[9]Mallin M.Phytoplankton ecology of North Carolina estuaries [J].Estuaries,1994,17: 561-574.
[10]Larned S T.Nitrogen-versus phosphorus-limited growth and sources of nutrients for coral reef macroalgae [J].Marine Biology,1998,132 (3): 409-421.
[11]Terrados J.nutrient limitation of the tropical sea grass Enhalus acoroides (L.)Royle in Cape Bolinao,NW Philippines [J].Aquatic Botany,1999,65 (14): 123-139.
[12]Thingstad T,Frede U L Z,Fereidoun R P.1imitation of heterotrophic bacteria and phytoplankton in the northwest Mediterranean [J].Limnology and Oceanogrophy,1998,43 (1):88-94.
[13]Ning X,Vaulot D,Liu Z,et a1.Standing stock and production of phytoplankton in the estuary of the Changjiang (Yangtze River)and the adjacent East China Sea [J].Marine Ecology Progress Series,1988,49 (10): l4l-150.
[14]胡明輝,徐春林.長(zhǎng)江口浮游植物生長(zhǎng)的磷酸鹽限制 [J].海洋學(xué)報(bào),1989,11 (4): 439-443.
[15]Wong G T F,Gong G C,Liu K K,et a1.Excess Nitrate’in the East China Sea [J].Estuarine,Coastal and Shelf Science,1998,46:411-4l8.
[16]鄒立,張經(jīng).渤海春季營(yíng)養(yǎng)鹽限制的現(xiàn)場(chǎng)實(shí)驗(yàn) [J].海洋與湖沼,2001,32(6): 672-678.
[17]王曉偉,李純厚,李占東,等.大亞灣海域春季浮游植物生長(zhǎng)的限制性營(yíng)養(yǎng)元素 [J].中國(guó)水產(chǎn)科學(xué),2007,14(5): 836-842.
[18]朱艾嘉,黃良民,許戰(zhàn)洲.氮、磷對(duì)大亞灣大鵬澳海區(qū)浮游植物群落的影響 [J].熱帶海洋學(xué)報(bào),2008,27(1): 38-45.
[19]王玉玨,洪華生,王大志,等.臺(tái)灣海峽上升流區(qū)浮游植物對(duì)營(yíng)養(yǎng)鹽添加的響應(yīng) [J].生態(tài)學(xué)報(bào),2008,28(3): 1 321-1 327.
[20]劉媛,曹振銳,黃邦欽,等.東、黃海典型海區(qū)浮游植物對(duì)營(yíng)養(yǎng)鹽添加的響應(yīng) [J].廈門大學(xué)學(xué)報(bào)(自然科學(xué)版),2004,43(增刊):147-152.
[21]唐松.中大西洋營(yíng)養(yǎng)鹽富集實(shí)驗(yàn)中浮游植物生長(zhǎng)的營(yíng)養(yǎng)鹽限制作用 [J].礦物巖石地球化學(xué)通報(bào).2008,27(2): 112-118.
[22]Eppley R W.Temperature and phytoplankton growth in the sea [J].Fishery Bulletin,1972,70: 1 063-1 085.
[23]Goldman J C,Carpenter E J.A kinetic approach to the effect of temperature on algal growth [J].Limnology and Oceanography,1974,19: 756-766.
[24]Yoder J A.Effect of temperature on light-limited growth and chemical composition of Skeletonema costatum (Bacillariophyceae)[J].Journal of Phycology,1979,15: 362-370.
[25]Sarmiento j L,Gruber N.Ocean biogeochemical dynamics [M].Princeton University Press,2006.
[26]John E H,Flynn K J.Growth dynamics and toxicity of Alexandrium fundyense (Dinophyceae): The effect of changing N:P supply ratos on internal toxin and nutrient levels [J].Eru J Phycol,2000,35:11-23.
[27]候繼靈,張傳松,石曉勇,等.磷酸鹽對(duì)兩種東海典型赤潮藻影響的圍隔實(shí)驗(yàn) [J].中國(guó)海洋大學(xué)學(xué)報(bào),2006,36: 163-169.
[28]康燕玉,梁君容,高亞輝,等.氮、磷比對(duì)兩種赤潮藻生長(zhǎng)特性的影響及藻間競(jìng)爭(zhēng)作用 [J].海洋學(xué)報(bào),2003,28(5): 117-122.
[29]劉東艷,孫軍,陳宗濤,等.不同氮磷比對(duì)中肋骨條藻生長(zhǎng)特性的影響 [J].海洋湖沼通報(bào),2002,2: 39-44.
In situ nutrient enrichment experiment with addition of nitrogen or phosphorus to surface seawater in the East Pacific Ocean
Tang Song
(Guangzhou Institute of Geochemistry,Chinese Academy of Sciences,Guangzhou 510640,China)
Q178.1+12; Q179.1
A
1001-6932(2010)04-0402-05
2009-11-09;
2009-11-25
國(guó)家自然科學(xué)基金資助項(xiàng)目(40903033,40403004)
唐松(1979-),男,陜西渭南人,博士,副研究員,主要從事海洋地球化學(xué)研究。電子郵箱:tangsong@gig.ac.cn
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學(xué)生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
