浙江省不同地區實驗兔血液蛋白多態性研究
2010-09-19 01:29:50趙偉春徐孝平李莉王德軍喻琦陳民利
中國實驗動物學報 2010年6期
關鍵詞:實驗
趙偉春,徐孝平,李莉,王德軍,喻琦,陳民利
(1.浙江中醫藥大學生物工程學院,杭州 310053;2.浙江中醫藥大學動物實驗研究中心,杭州310053;3.浙江工業大學藥學院,杭州 310014)
浙江省不同地區實驗兔血液蛋白多態性研究
趙偉春1,徐孝平2,李莉1,王德軍2,喻琦3,陳民利2
(1.浙江中醫藥大學生物工程學院,杭州 310053;2.浙江中醫藥大學動物實驗研究中心,杭州310053;3.浙江工業大學藥學院,杭州 310014)
目的 研究浙江省不同產地實驗兔血液蛋白的多態性,了解各地實驗兔的遺傳背景,指導優良品系的選育。方法 選擇蘭溪、農科院、上虞和余姚等地生產的實驗兔的11個血液蛋白位點進行聚丙烯酰胺凝膠電泳,根據電泳圖譜分析基因頻率、Hardy-weinberg平衡狀態、遺傳變異程度和遺傳相似性。結果 Pr、Ptf、Hbα、Hbβ和Alb在所有實驗兔中表現為單態,其余蛋白位點呈多態。各蛋白位點的變異程度Prt1>Sag>Tf1>Po>Tf2>Prt2。各地實驗兔蛋白位點平均雜合度為蘭溪>農科院>上虞>余姚。農科院與余姚實驗兔間的遺傳距離最近,上虞與蘭溪實驗兔的親緣關系最遠,遺傳相似系數分別為0.7938和0.7687。結論 血液蛋白多態性反映了浙江省不同地區生產的實驗兔的遺傳背景,為實驗兔的選育提供了實驗依據。
兔;血液蛋白多態性;基因頻率;雜合度;遺傳距離
實驗兔是生物醫藥產品研發及醫學科學研究領域最常用的生物研究材料。由于兔的繁殖場地較大,實驗室進行大規模的養殖比較困難,目前各實驗室使用的實驗兔大部分來自于農科系統單位和農民個體養殖場等作為經濟動物飼養的兔。由于不同的飼養方式和環境條件導致各種群兔的生物學特性發生改變,質量參差不齊。為規范實驗兔的飼養管理,穩定實驗兔的生物性狀,提高實驗兔的質量,浙江省實驗動物平臺對有一定飼養基礎的浙江省農業科學院畜牧獸醫研究所、上虞市星火兔業養殖場、余姚市泗門建飛實驗兔養殖場等兔場進行引導扶植,按實驗動物的生產設施和管理體系要求,擴大實驗兔的飼養規模,提高實驗兔的遺傳品質。本實驗通過聚丙烯酰胺凝膠電泳對農科院、上虞和余姚生產的日本大耳白兔的后白蛋白(Po)、前轉鐵蛋白1(Prt1)、前轉鐵蛋白2(Prt2)、轉鐵蛋白1(Tf1)、轉鐵蛋白2(Tf2)、后轉鐵蛋白(Ptf)、慢α球蛋白(Sag)、血紅蛋白 α(Hbα)、血紅蛋白β(Hbβ)和白蛋白(Alb)共11個血液蛋白位點進行了多態性的研究,并以蘭溪農戶生產的日本大耳白兔作為對照,為了解規范化飼養后各養殖場實驗兔的遺傳背景,以及與經濟動物方式飼養的差異提供參考,為實驗兔的生產和應用提供指導。
1 材料與方法
1.1 實驗動物
取日本大耳白兔的同齡母兔同期繁殖生產的后代,共129只(雌64只,雄65只),體質量(2.25±0.25)kg。其中,浙江省農業科學院畜牧獸醫研究所養兔場30只,雌雄各半,生產許可證【SCXK(浙)2006-0025】,以下簡稱“農科院兔”;余姚市泗門建飛實驗兔養殖場 40只,雌雄各半,生產許可證【SCXK(浙)2008-0026】,以下簡稱“余姚兔”;上虞市星火兔業養殖場29只,雌14只,雄15只,生產許可證【SCXK(浙)2008-0036】,以下簡稱“上虞兔”;蘭溪農戶采購30只,雌雄各半,由浙江中醫藥大學動物實驗研究中心采購,以下簡稱“蘭溪兔”。所有實驗兔均預飼養一周后,進行血樣采集。浙江中醫藥大學動物實驗研究中心兔實驗飼養室,使用許可證【SYXK(浙)2008-0116】。
1.2 血樣采集
每只實驗兔采集血樣2 mL,其中1 mL血樣加入至肝素抗凝管中制備紅細胞溶血液(抗凝管的制備:在1.5 mL離心管中加入每滴含30 U的肝素溶液2滴,于60~80℃烘干),反復倒置,使抗凝劑與血液混勻;剩余血樣加至不加抗凝劑的離心管中用于制備血清。
1.3 樣品的制備
取未加抗凝劑的血樣離心(2000 r/min,10 min,4℃),將血清轉移至 0.5 mL離心管中,– 20℃保存。將加抗凝劑的血樣離心(2000 r/min,10 min,4℃)后棄上層血漿,用10倍于紅細胞壓積的生理鹽水洗滌 3 次(2000 r/min,10 min,4℃),取洗滌過的紅細胞40 μL于0.5 mL離心管中,用等體積的蒸餾水稀釋,震蕩使紅細胞破裂溶血,制成紅細胞溶血液,–20℃保存。
1.4 Pr、Ptf、Po、Sag、Prt1、Prt2、Tf1 和 Tf2 的電泳測定
采用聚丙烯酰胺凝膠電泳對各實驗兔的血清樣品進行檢測。電泳系統如下:電泳緩沖液為Tris-甘氨酸(pH 8.3),分離膠緩沖液為 Tris-HCl(pH 8.9),濃縮膠緩沖液為 Tris-HCl(pH 6.7),濃縮膠和分離膠濃度分別為3.5%和12%。各實驗兔的血清樣品先用PBS稀釋5倍后,再與等體積的上樣緩沖液(2×)混勻,取上述混合液15 μL上樣。每一塊凝膠均加了同一個實驗兔的血液樣品作為對照,下同。樣品在濃縮膠和分離膠中的泳動電壓分別為150 V和350 V,電泳時間16 h。整個過程在4℃下進行。采用考馬斯亮藍法染色1 h,然后用甲醇:冰乙酸脫色液脫色至背景無色為止。
1.5 Alb的電泳測定
Alb位點電泳時分離膠濃度改為8%,各實驗兔的血清樣品先用PBS稀釋30倍后,再與等體積的上樣緩沖液(2×)混勻,取上述混合液15 μL上樣,電泳4 h。其他實驗條件同前。
1.6 Hb的電泳測定
Hb位點電泳加樣時,將紅細胞溶血液用 PBS作10倍稀釋,再與等體積的上樣緩沖液(2×)混勻,取上述混合液15 μL上樣。Hb屬于有色蛋白,無須染色即可觀察到分離的帶型。其他實驗條件同Alb位點的電泳檢測。
1.7 數據分析
各等位基因及表型識別均參考 Grunder等[1]和Arana等[2]的標準。根據表型頻率,采用基因計數法計算基因頻率。根據基因型頻率實際觀測值計算理論值,并用χ2適合性檢驗比較實際值與理論值的差異,檢驗二者的差異是否顯著,了解群體的基因頻率是否符合Hardy-Weiberg平衡定律。計算平均雜合度、有效等位基因數和奈氏遺傳距離,采用UPGMA法進行聚類分析。
2 結果
2.1 血清蛋白位點基因型頻率及基因頻率的分析
2.1.1 Pr、Po、Prt1、Prt2、Tf1、Tf2、Ptf和 Sag 蛋白位點基因型頻率及基因頻率分析:Pr、Po、Prt1、Prt2、Tf1、Tf2、Ptf和 Sag位點的部分電泳圖譜見圖 1,基因型頻率及基因頻率結果見表1。在被檢測的所有實驗兔個體中,Pr和Ptf的電泳圖譜均只有一個條帶,為單態位點。Po在所有實驗兔個體中均表現出AB一種表型。蘭溪和農科院兔的Prtl均有6種表型,而上虞兔該蛋白位點無 BC表型,余姚兔則無CC表型。Prt2的表型由2個基因所控制,在4個實驗兔群中均無BB表型,只有AA型和 AB型。Tf1位點的表型由3個等位基因所控制,其中余姚的無AA和AC表型,其余3個實驗兔群則無 AA和 CC表型。Tf2的表型由2個等位基因所控制,其中蘭溪和余姚兔無BB表型,農科院和上虞兔則AA、BB和AB這3種表型均有。Sag在4個實驗兔群中均無CC表型,其中余姚兔無C基因,且只呈現AB表型,蘭溪兔無BB和CC表型,農科院兔無CC和BC表型,上虞兔則無AA和CC表型。
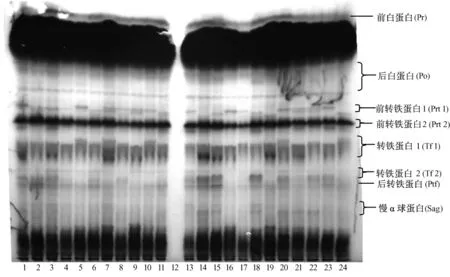
注:第12泳道為空白對照;第1~8泳道為蘭溪樣品;第9~11泳道為農科院樣品;第13~17泳道為上虞樣品;第18~24泳道為余姚樣品。圖 1 Pr、Po、Prt1、Prt2、Tf1、Tf2、Ptf和 Sag 的電泳圖譜Note:Lane 12 was blank controls.Lanes 1 to 8 were samples from rabbits bred in Lanxi.Lanes 9 to 11 were samples from the Agricultural Academy.Lanes 13 to 17 were samples from Shangyu.Lanes 18 to 24 were samples from Yuyao.Fig.1 Electrophoresis patterns of Pr,Po,Prt1,Prt2,Tf1,Tf2,Ptf and Sag
2.1.2 血紅蛋白(Hb)和白蛋白(Alb)蛋白位點基因型頻率及基因頻率分析:在所有被檢測的樣品中,Hb只分離出 Hbα和 Hbβ兩種成分,泳動較快的Hbα比泳動較慢的Hbβ成分少(圖2)。Alb呈均一的球狀帶(圖3)。
2.2 蛋白位點的Hardy-Weinberg平衡狀態分析
由于 Pr、Ptf、Hbα、Hbβ 和 Alb 位點為單態位點,不進行Hardy-Weinberg平衡狀態分析,其余位點的統計結果見表2。經χ2適合性檢驗,在蘭溪生產的實驗兔中Prt1、Prt2和Tf1為平衡位點,Po為高度不平衡位點(P<0.01),Tf2和 Sag為不平衡位點(0.01<P<0.05)。在農科院生產的實驗兔中 Po和Tf1是高度不平衡位點(P<0.01),其余位點均為平衡位點(P>0.05)。在上虞生產的實驗兔中Po和Prtl為高度不平衡位點(P<0.01),其余位點均為平衡位點(P>0.05)。余姚生產的實驗兔則以Po和Sag為高度不平衡位點(P<0.01)。
2.3 蛋白位點變異程度的分析
如表3所示,6個多態蛋白位點的平均雜合度之和在4個產地實驗兔間存在差異,排列次序為蘭溪>農科院>上虞>余姚,各蛋白位點的變異程度Prt1>Sag>Tf1>Po>Tf2>Prt2。Prt1在各實驗兔
群中的變異程度均最大,而 Prt2的變異程度均最小,其余蛋白位點的變異程度在各實驗兔群間存在差異,其中蘭溪兔蛋白位點的雜合度為Sag>Tf1>Po>Tf2;農科院兔為 Tf1>Sag>Po>Tf2;上虞兔為Sag>Tf1>Po>Tf2;余姚兔則 Po=Sag>Tf1>Tf2。
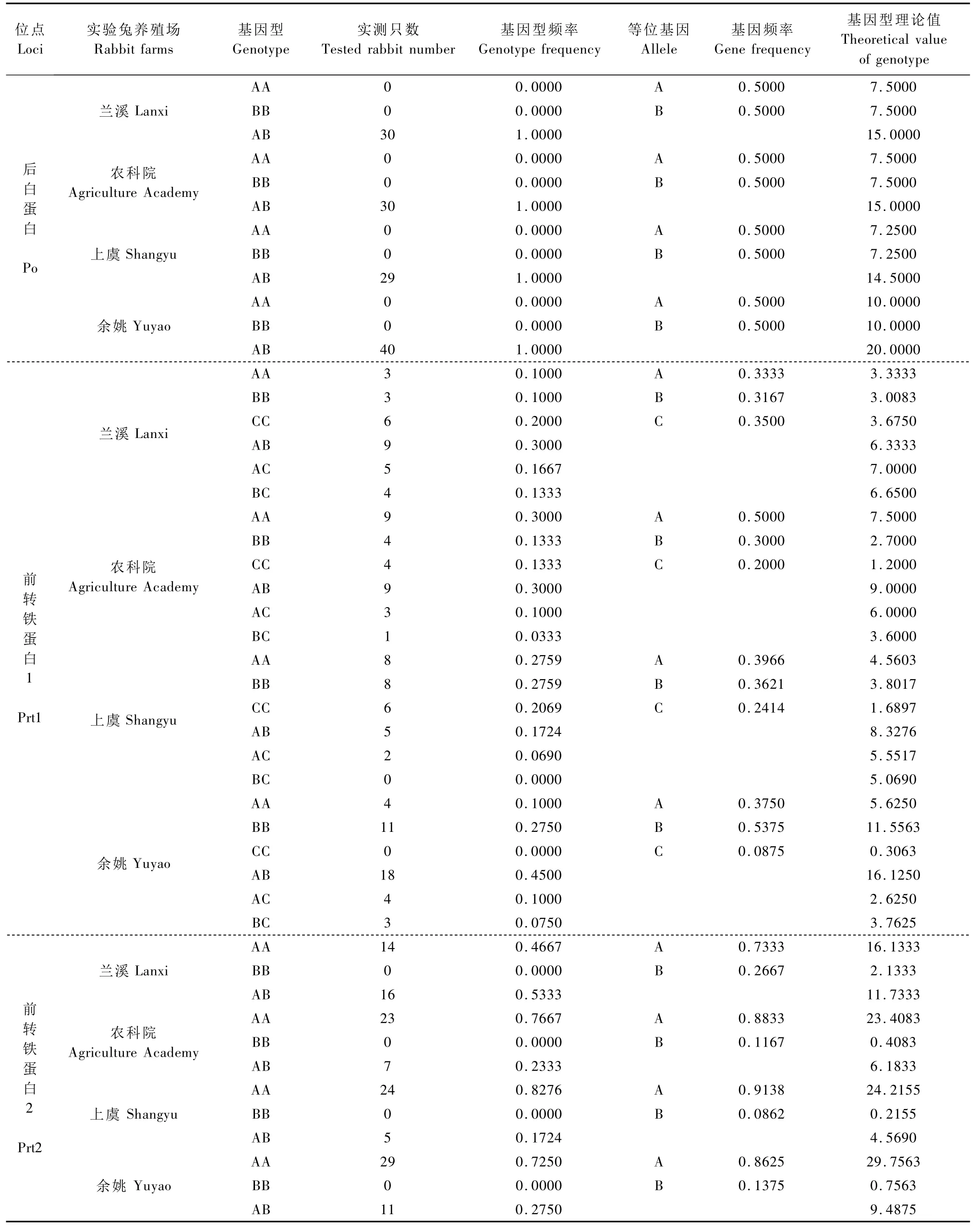
表1 Po、Prt1、Prt2、Tf1、Tf2和 Sag位點的基因型頻率和基因頻率Tab.1 Genotype frequency and gene frequency of Po,Prt1,Prt2,Tf1,Tf2 and Sag
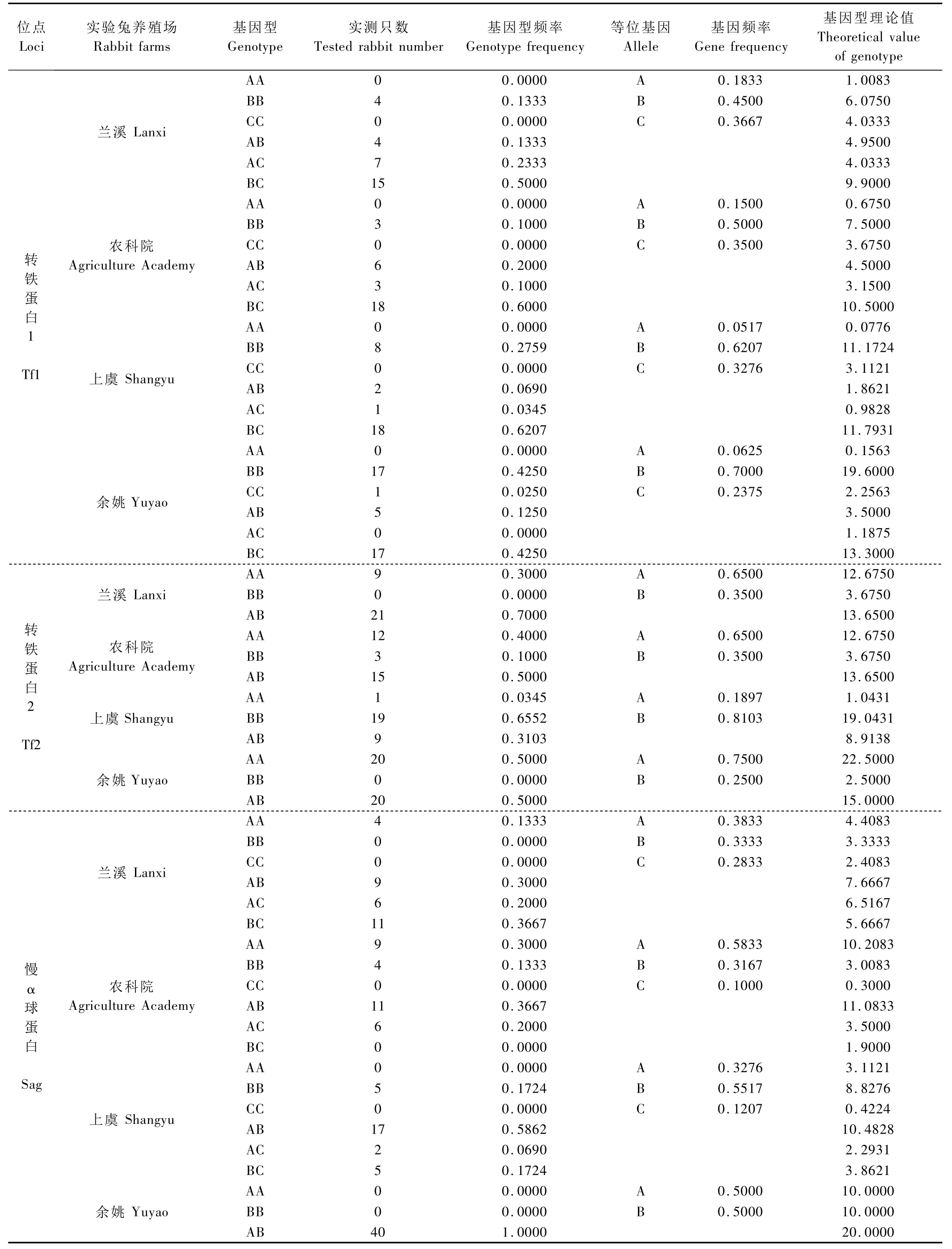
續表
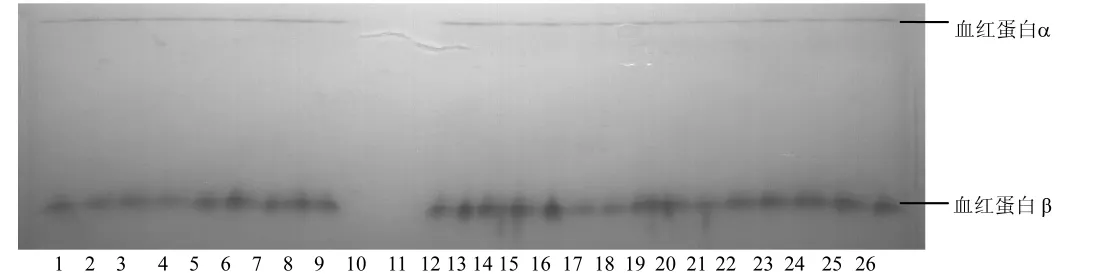
注:第10和11泳道為空白對照,1~9泳道為蘭溪兔樣品,12~26為農科院兔樣品。圖2 血紅蛋白的電泳圖譜Note:Lanes 10 and 11 were blank controls.Lanes 1 to 9 were samples of rabbits bred in Lanxi.Lanes 12 to 26 were samples of Agriculture Academy.Fig.2 Electrophoretic patterns of hemoglobin
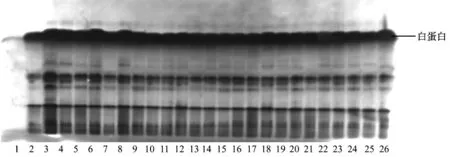
注:第1泳道為空白對照;第2~19泳道為農科院兔的血清樣品;第20~26泳道為蘭溪兔的血清樣品。圖3 白蛋白的電泳圖譜Note:Lane 1 was blank control.Lanes 2 to19 were samples of rabbits from the Agriculture Academy.Lanes 20 to 26 were samples of rabbits from Lanxi.Fig.3 Electrophoretic patterns of serum albumin
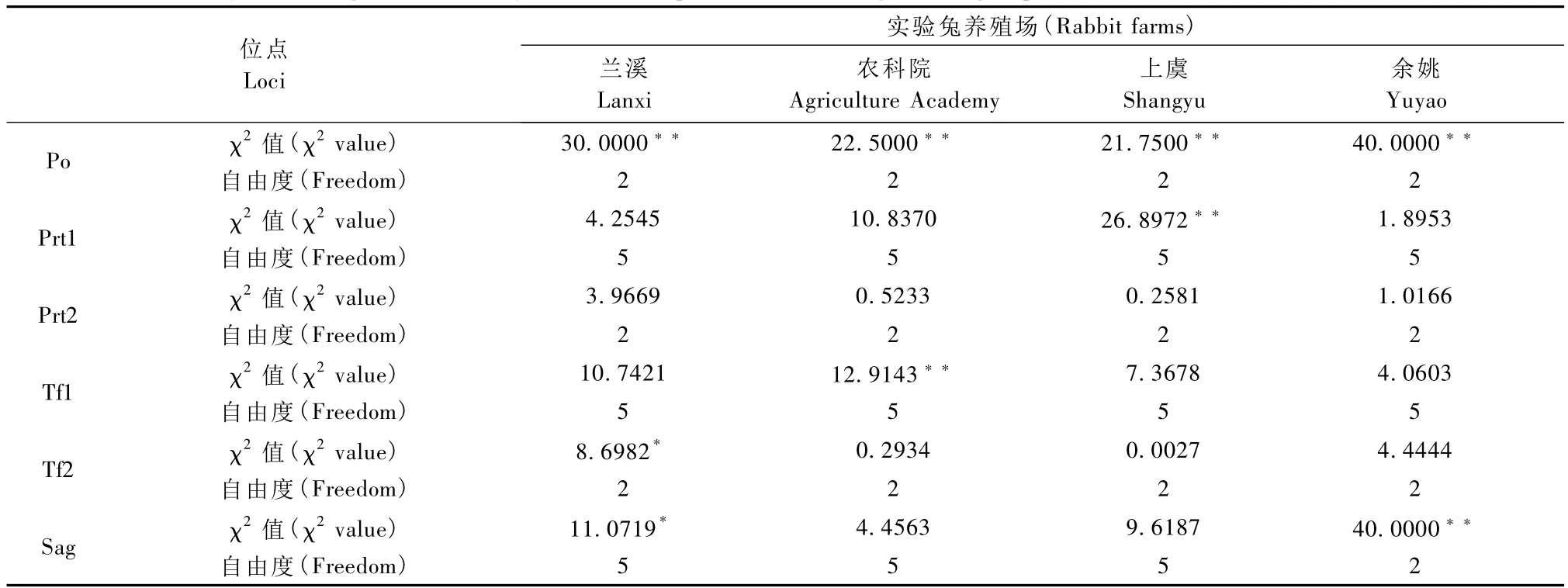
表2 4個養殖場實驗兔血液蛋白位點的Hardy-weinberg平衡狀態分析Tab.2 Hardy-Weinberg balance analysis of serum protein loci among rabbit groups bred in different farms
2.4 不同產地實驗兔群的遺傳相似性分析
4個產地的實驗兔群中(圖4),農科院、余姚和蘭溪兔間的遺傳相似系數均大于0.78,其中農科院與余姚兔間的遺傳距離最近,遺傳相似系數為0.7938。上虞兔與其他3個產地實驗兔的遺傳相似系數均較小,其中與蘭溪兔的遺傳距離最遠,遺傳相似系數為0.7687。
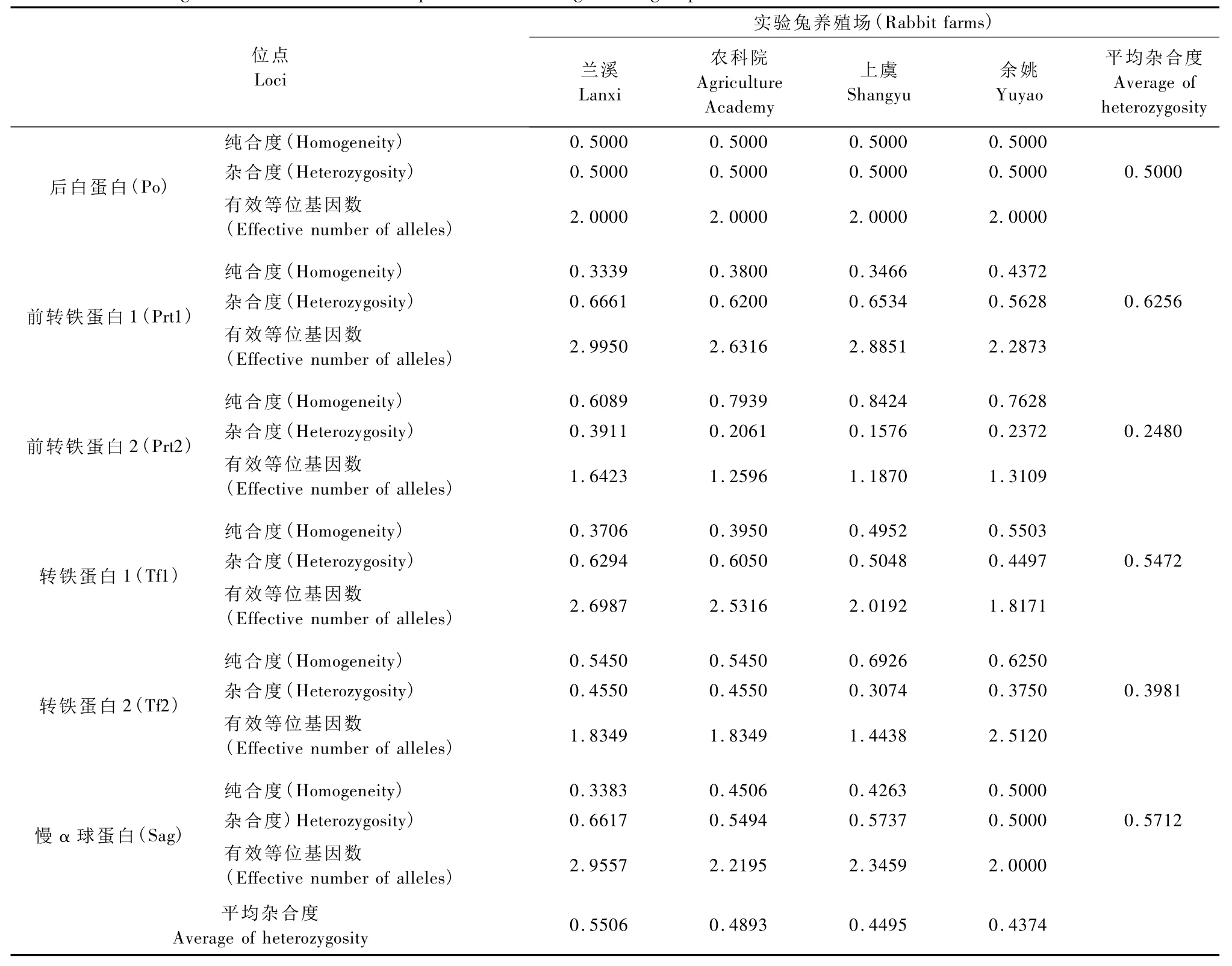
表3 4個養殖場實驗兔血液蛋白位點變異程度Tab.3 The genetic variation of serum protein loci among rabbit groups bred in different farms

圖4 4個養殖場實驗兔間的UPGMA聚類分析Fig.4 Cluster analysis on the similarity values of the four rabbit groups bred in different farms by UPGMA
3 討論
3.1 基因型頻率和基因頻率分析
等位基因頻率的明顯差別從理論上可用于標記品種特征。在本實驗的4個產地實驗兔群中,Pr、Ptf、Hbα、Hbβ 和 Alb 都表現單態,在長期的進化過程中保持了一種遺傳的穩定性。這與其他家兔的研究結果一致[3-5]。已有研究表明,Po受兩個共顯性等位基因控制,如中國白兔、丹麥兔、比利時兔和加利福尼亞兔[6,7]。本研究發現 Po在蘭溪、農科院、上虞和余姚生產的實驗兔中均表現為AB表型。Prt在中國白兔、新西蘭兔、丹麥兔、比利時兔和加利福尼亞兔中均存在多態性[8,9]。本研究中僅蘭溪和農科院生產的實驗兔Prtl出現6種表型,其余產地兔群均只出現其中某幾種表型。Tf在青紫藍兔、新西蘭兔、家利福尼亞兔、丹麥兔等多數家兔品種中表現出多態性[10,4],僅在少數幾個品種中表現單態,如塞北兔[11]。本研究的 Tfl表現出 5種表型,無 AA表型。Sag由A、B和 C這3個等位基因控制,A、B為優勢基因,基因型的表現不一樣。各實驗兔群不同位點的等位基因頻率不同,這是由于不同實驗兔其遺傳背景、選育方法和選育程度不同造成的。我國實驗兔除中科院上海實驗動物中心1979年從日本引進新西蘭兔和2003、2004年相繼從美國引進SPF級新西蘭兔和日本大耳白兔進行少量飼養外,全國各地極大部分的實驗兔來自以特種經濟動物引入我國飼養的新西蘭兔和日本大耳白兔。這些種群的飼養歷史悠久,由于長期在特殊的飼養環境中飼養,以及在繁殖過程中為了獲得高繁殖率,相互間有雜交現象,并未保持動物本身的遺傳系,造成了血緣雜交和品系間的遺傳污染,出現遺傳學上的基因突變或漂變。
3.2 多態蛋白位點的Hardy-Weinberg平衡狀態分析
χ2適合性檢驗結果表明,在所檢測的實驗兔中Po均為高度不平衡位點(P<0.01),Tf1、Prtl位點上也有多個實驗兔群為高度不平衡位點,而 Prt2、Tf2和Sag以平衡位點為主。這些不平衡位點的基因頻率偏離 Hardy-Weinberg平衡狀態,說明這些位點受到了選擇、突變等因素的影響。
3.3 群體遺傳變異分析
4個產地實驗兔群11個蛋白位點的平均雜合度存在差異。其中蘭溪兔個體間變異程度最大,其次為農科院兔,上虞和余姚兔個體間變異較小。這表明蘭溪和農科院兔的遺傳多樣性較豐富,個體間差異較大。這是由于蘭溪和農科院兔來自以特種經濟動物引入,飼養歷史悠久,在長期飼養過程中,日本大耳白兔有與新西蘭兔種群和中國實驗兔的雜交,具有復雜的血緣關系。而上虞和余姚兔種群的飼養時間短,規模比較小,實驗兔以封閉繁殖為主,未引進外來血緣,因此個體間的遺傳品質相對較純。Prt1在4個實驗兔群中的變異程度均較大,表明該位點的選擇潛力大。
3.4 群體遺傳相似性
在4個產地的實驗兔群中,農科院兔、余姚兔與蘭溪兔群間的遺傳相似系數均較大,而上虞兔與其他3個產地實驗兔的遺傳距離均較遠,這是由于上虞兔引入的日本大耳白兔,引入時間短,沒有外來血緣,品種較純正。其余3個產地的日本大耳白兔,飼養時間長,已帶有新西蘭兔種群和中國本兔的血緣,故與其血緣關系較遠。因此,血液蛋白位點的多態性較好的反映了各群體的遺傳背景。
[1] Grunder AA,Sartore G,Stormont C.Genetic variation in red cell esterase of rabbits[J].Genetics.1965,52(6):1345-1353.
[2] Arana AP,Zaraoza P,Rodeuar C,et al.Evidence for transferrin polymorphism in Spanish wild rabbits[J].Animal Genetics.1987,18(2):125-132.
[3] 周立波,任文陟,張嘉保,等.Vc-獺兔、日本大耳白兔與新西蘭白兔血液蛋白多態性研究[J].動物醫學進展.2004,25(3):102-105.
[4] 孫杰,趙宗盛,李大全,等.日本大耳白兔、新西蘭兔血液蛋白多態性的研究[J].中國畜牧雜志.2002,38(1):7-8.
[5] Ferrand N. Biochemical and genetic studies on rabbit haemoglobin II Eeectrophoretic polymorphism of the α chains[J].Biochem Genetics.1990,21(2):217-219.
[6] 龐有志.家兔血液蛋白多態性研究進展[J].獸醫大學學報.1992,12(4):71-37.
[7] 劉若余.家兔血清蛋白遺傳多態性[J].貴州農學院學報.1997,16(1):22-24.
[8] Usher DC,Cogbum B,Richard RF.Rabbit linkage group VIII:the alleles of Prt gene[J].Biocheml Genetics.1983,21(4):511-526.
[9] Vieira J, Ferrand N. Fournew allelesatthe mannose-6-phosphate isomerase locus in rabbit[J].Animal Genetics.1995,26(1):37-38.
[10] Markovich LG. Transferrin polymorphism in different rabbits breeds[J].Animal Breed Abst.1977,45(1):70.
[11] 吳淑琴,孫全文,穆秀明,等.塞北兔血液蛋白多態性研究[J].中國養兔雜志.2002,(4):17-21.
Serum Protein Loci Polymorphism in Laboratory Rabbits Bred in Different Regions of Zhejiang Province
ZHAO Wei-Chun1,XU Xiao-Ping2,LI Li1,WANG De-Jun2,YU Qi3,CHEN Min-Li2
(1.College of Bioengineering,Zhejiang Chinese Medical University,Hangzhou 310053,China;2.Laboratory Animal Research Center,Zhejiang Chinese Medical University,Hangzhou 310053,China;3.College of Pharmaceutical Science,Zhejiang University of Technology,Hangzhou 310014,China)
ObjectiveTo study the serum protein loci polymorphism in rabbits bred in different regions of Zhejiang province.Method Eleven blood protein loci of rabbits bred in four regions of Zhejiang province were analyzed by vertical polyacrylamide gel electrophoresis.The rabbits were bred in the Agriculture Academy,in Lanxi,Shangyu and Yuyao,respectively.The proteins included the prealbumin(Pr),postalbumin(Po),pretransferrin 1 (Prt1),pretransferrin 2(Prt2),transferrin 1(Tf1),transferrin 2(Tf2),posttransferrin(Ptf)and slow α-globulin(Sag),respectively.The gene frequency,Hardy-Weinberg balance,genetic variation and similarity coefficient of the four groups of rabbits were analyzed.ResultsPr,Ptf,Hbα,Hbβ and Alb in all rabbits had a monomorphism,while the others were polymorphic.The genetic variation of serum protein loci were Prt1>Sag>Tf1>Po>Tf2>Prt2.The heterozygosity of serum protein loci were Lanxi>Agriculture Academy>Shangyu>Yuyao.The genetic distance between the rabbits bred in Agricultural Academy and Yuyao was the nearest among those four regions.The similarity coefficient was 0.7938.The relationship between the rabbits bred in Shangyu and Lanxi was the farthest,with a similarity coefficient of 0.7687.ConclusionsThe serum protein polymorphism reflects the genetic background of the rabbits bred in different regions of Zhejiang province.It provides an experimental basis for breeding of rabbits.
Rabbits;Serum protein polymorphism;Gene frequency;Heterozygosity;Genetic distance
Q346
A
1005-4847(2010)06-0506-08
10.3969/j.issn.1005-4847.2010.06.013
2010-06-24
浙江省科技廳實驗動物公共服務平臺項目(No.2008F80017,No.Dwpt-bz-12),浙江省衛生高層次創新人才培養工程項目(2008年)。
趙偉春(1973-),女,副研究員,博士,研究方向:生物工程。E-mail:weichunzhao@hotmail.com
陳民利。E-mail:minlichen01@yahoo.com.cn
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
