線粒體加工肽酶干擾 RNA真核表達載體的構建
2010-09-20 08:03:52張艷梅呼格吉樂馬飛煜馬翔凌
中風與神經疾病雜志 2010年1期
關鍵詞:研究
張艷梅, 呼格吉樂, 馬飛煜, 鄔 偉, 馬翔凌
帕金森病(Parkinson's Disease,PD)是一種常見的神經系統變性病,其發生、發展機制涉及多基因表達改變[1]。盡管 PD的發病機制目前仍不完全清楚,但一些誘發因素在不同階段作用于不同基因,引起相關基因結構及表達水平改變而致病。MPPs的主要功能是特異識別一大類線粒體前體蛋白,并在單一和特定位點剪切這類蛋白,很可能與 PD有關[2]。在本研究的前期工作中利用蛋白組學技術對MPP+誘導的 PC12細胞 PD模型中發現線粒體加工肽酶(m itochondrial processing peptidases,MPPs)表達量變化顯著[3],因此本研究采用干擾 RNA技術特異性剔除或關閉 MPP+基因的表達,使培養細胞中的靶定基因沉默。因此本實驗針對 Pmpca基因,設計并構建 siRNA(small interference RNA,siRNA)表達載體,為進一步在體內外研究 MPPs的功能奠定基礎。
1 材料與方法
1.1 材料
PC12細胞購自中科院上海細胞庫;AXYGEN質粒小量提取試劑盒購于杭州愛思進生物技術有限公司;BamH I、Bbs I購自 sigma公司;干擾 RNA的合成購自上海吉瑪公司;測序工作在上海 Invitrogen公司完成。
1.2 方法
1.2.1 寡核苷酸的設計與合成
因國內外對 Pmpca-siRNA表達載體的構建,未見報道。本研究根據 Pmpca的所有變異體的共同序列設計出 shRNA,經 blast軟件進行同源搜索,確定其特異性。以下序列為 Pmpca部分 cDNA序列,黑體部分為靶序列即干擾部位。


shRNA模板中的 loop結構選用了 TTCAAGAGA以避免形成終止信號,shRNA的轉錄終止序列采用T6結構。正義鏈模板的 5'端添加了 CACC,與 BbsI酶切后形成的粘端互補;反義鏈模板的 5'端添加了GATC,與 BamHI酶切后形成的粘端互補;如果 siRNA的第一個堿基不是 G,則在 CACC后補加一個G,以下是以 SR-Pmpca-1425為例具體說明模板的設計。

1.2.2 shRNA模板的退火
將 DNA oligo分別用 TE(pH 8.0)溶解,濃度為100μmol/L。取相應的正義鏈和反義鏈 oligo溶液。退火條件:95℃ 5min;85℃ 5min;75℃ 5m in;70℃5min;4℃保存。退火處理后得到濃度為 10μM的shRNA模板。將所得模板溶液稀釋 500倍,終濃度為 20nmol/L,用于連接反應。
1.2.3 PGPU6/GFP/Neo載體的線性化
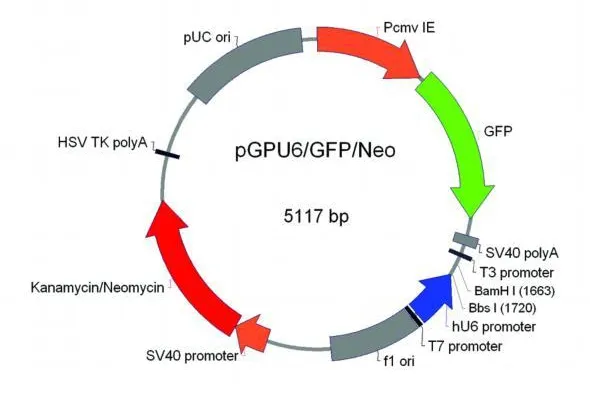
圖1 PGPU6/GFP/Neo載體圖
取 2μg PGPU6/GFP/Neo載體進行酶切處理:37℃酶切1h,瓊脂糖電泳,使用 Agarose Gel DNA Purification Kit Ver2.0(TaKaRa)回收,電泳檢測估算濃度,稀釋濃度至 50ng/ul。
1.2.4 PGPU6/GFP/Neo-shRNA載體的構建
(1)載體的連接反應:22℃ 1h,轉到 JM109感受態細胞中。
(2)每個連接反應挑取 5個菌落,接種到含50μg/ml Kanamycin的 LB培養基中,37℃孵育過夜。挑取過夜培養平板上單個菌落置于 5ml含卡那霉素(30μg/m L)的 LB培養基中,37℃,250r/min振蕩培養12h。
(3)質粒提取:取 5m l在 LB培養基中培養過夜的菌液于離心管中,室溫 11200r/min離心 1min,棄盡上清;用 250μl已加入 RNase A的 Buffer S1懸浮細菌沉淀,加入 250μl Buffer S2,翻轉混合 4~6次,混合均勻,使菌體充分裂解,形成透亮的溶液。加入350μl Buffer S3,翻轉混合 6~8次,11200r/min離心10min;吸取離心上清并轉移到 DNA制備管中,11200r/min離心 1m in,棄濾液;將制備管置回離心管 ,加 500μl BufferW1,11200r/min離心 1min,棄濾液;將制備管置回離心管,加 700μl Buffer W2,11200r/min離心 1m in,棄濾液;以同樣的方法再用700μl BufferW 2洗滌 1次。確認在 Buffer W2 concentrate中已按試劑瓶上的指定體積加入無水乙醇;將制備管置回 2m l離心管中,11200r/min離心1min;將制備管移入新的 1.5ml離心管中,在 DNA制備管膜中央加 60~80μl Eluent或去離子水,室溫靜置 1min。 11200r/m in離心 1min。將 Eluent或去離子水加熱至 65℃將提高洗脫效率;使用堿裂解法抽提質粒,所得質粒用 Bam H I,Pst I分別酶切鑒定(見圖2、圖3),測序鑒定。
2 結 果
2.1 質粒酶切
使用堿裂解法抽提質粒,所得質粒用 Bam H I,Pst I分別酶切鑒定,陽性重組載體應該可以被BamH I切開,而不能被 Pst I切開。酶切結果表明,所有質粒均為陽性重組載體(見圖2、圖3)。
2.2 測序鑒定
每組選擇兩個克隆進行測序鑒定。構建的 siRNAs序列與基因庫中序列完全相同,并且未發現有突變、缺失、插入等異常存在。
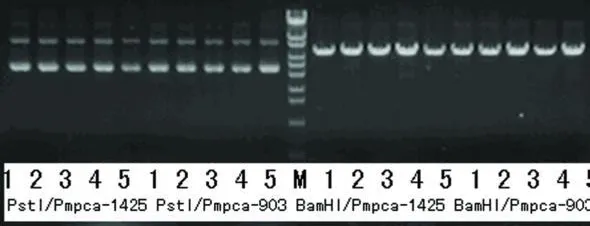
圖2 重組載體的酶切鑒定
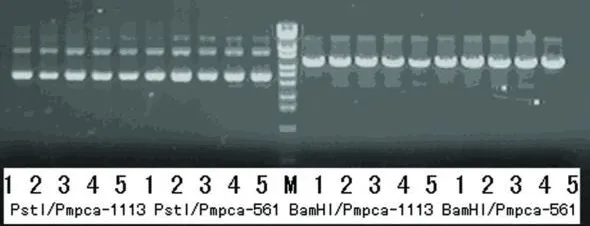
圖3 重組載體的酶切鑒定
3 討 論
RNAi技術的基本原理是將雙鏈 RNA(doublestranded RNA,dsRNA)裂解為 21~25個核苷酸組成的 siRNA作為介導子,引起同源序列特異性的 mRNA降解[4]。siRNA是 RNA干擾過程中的重要中間分子。體外制備的方法包括直接化學合成或利用Dicer或 RNA酶Ⅲ消化長的 dsRNA,也可利用質粒和病毒載體構建載體在細胞內表達[5]。RNAi技術近年來發展迅速,已成為分子生物學研究的主要技術手段之一,如 siRNA脂質體介導法[6],抑制艾滋病病毒[7,8]癌癥的早期發現[9]等,已逐漸被應用于人類基因功能研究和基因治療方面。當然,開發這類RNAi治療需要花費幾年甚至更長的時間,但隨著研究的不斷深入將會有更多以 RNAi機制為基礎的治療手段被用于臨床[10]。
線粒體加工肽酶(mitochond rial processing peptidases,MPPs)是一種金屬內切蛋白酶[11],它也是由 α和 β兩個亞單位組成的可溶性的異源二聚體,分子量是 100~110kD。MPPs的主要功能是特異識別一大類線粒體前體蛋白,并在單一和特定位點剪切這類蛋白。目前關于 MPPs與神經系統變性病的研究開展很少,但從上述研究中可以看出,MPPs與神經變性病的發生有著一定的聯系,但在國內外 PD研究的相關文獻中未見該蛋白的報道。張艷梅等[3]研究首次發現 MPPs蛋白在 PD的發病中可能起一定的作用,值得深入研究以闡明其詳盡的作用機制。
本研究通過構建 shRNA的真核表達載體,旨在為帕金森病的基因治療提供一種有效手段。MPPs真核表達載體的構建未見報道。本研究根據該基因的所有變異體的共同序列設計出 shRNA,經 blast軟件進行同源搜索,確定其特異性,模板為含有靶點序列回文結構,這樣細胞內合成的 RNA為發夾樣雙鏈結構,可產生分子內莖-環結構,被內源性 Dicer酶處理成 21nt的雙鏈 siRNA,發揮 siRNA的效應,達到特異性抑制靶基因表達的目的。再用 PAGE方式純化,確定其準確性。通過與真核表達載體PGPU6/GFP/Neo連接,酶切鑒定和基因測序證明 MPPs特異性 siRNA真核表達載體 PGPU6/GFP/Neo-Pmpca-SR-1113、PGPU6/GFP/Neo-Pmpca-SR-1425、PGPU6/GFP/Neo-Pmpca-SR-561、PGPU6/GFP/Neo-Pmpca-SR-903構建成功。
綜上所述,本實驗為進一步研究 MPPs基因功能缺陷對 PC12細胞的侵襲、增值的影響,以及雙向電泳探究基因功能缺陷后蛋白的表達奠定了基礎。
[1]李淑娟,胡國華,過紅明,等.白介素-1β和小膠質細胞在帕金森病中的作用[J].中國老年學雜志,2009,29(5):1189-1191.
[2]Taylor AB,Smith BS,Kitada SJ,etal.Crystal structuresofmitochondrial processing peptidase reveal the mode for specific cleavage of import signal sequences[J].Structure Cambridge,2001,9(7):615-625.
[3]張艷梅,馬飛煜,胡林森,等.蛋白質組學技術鑒定帕金森病模型中的 3種新蛋白質[J].國際神經病學神經外科學雜志,2008,35(4):291-294.
[4]呼格吉樂,王震宇,王文禮,等.RNAi技術在腫瘤研究中的應用進展[J].疾病監測與控制雜志,2009,25(1):61-63.
[5]Stockinger A,Eger A,Wolf J,etal.E-cadherin regulates cell growth bymodulating proliferation-dependent beta-catenin transcriptional activity[J].Cell Biol,2001,154(6):1185-1196.
[6]Durcan N,Murphy C,Cryan SA.Inhalable siRNA:potentialas a therapeutic agent in the lungs[J].Mol Pharm,2008,5(4):559-566.
[7]Kumar P,Ban HS,Kim SS,etal.T cell-specific siRNA delivery suppresses HIV-1 infection in humanized mice[J].Cell,2008,134(4):577-586.
[8]Liu YP,Gruber J,Haasnoot J,etal.RNAi-mediated inhibition of HIV-1 by targeting partially complementary viral sequences[J].Nucleic Acids Res,2009,37(18):6194-6204.
[9]Quon K,Kassner PD.RNA interference screening for the discovery of oncology targets[J].Expert Opin Ther Targets,2009,13(9):1027-1035.
[10]Grosshans H,Filipowicz W.Molecular biology:the expanding world of small RNAs[J].Nature,2008,451(7177):414-416.
[11]Oleksandr G,Patrizia C,Grazia I.Mitochondrial processing peptidases[J].Biochim Biophy Acta,2002,1592(1):63-77.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
