谷氮酰胺轉(zhuǎn)胺酶對谷朊質(zhì)構(gòu)和流變性質(zhì)的影響
2010-11-02 07:37:48晁麗靜孫貴朋
食品工業(yè)科技 2010年3期
晁麗靜,王 淼,孫貴朋
(1.江南大學食品科學與技術(shù)國家重點實驗室,江蘇無錫 214122; 2.江南大學食品學院,江蘇無錫 214122)
谷氮酰胺轉(zhuǎn)胺酶對谷朊質(zhì)構(gòu)和流變性質(zhì)的影響
晁麗靜1,王 淼2,*,孫貴朋2
(1.江南大學食品科學與技術(shù)國家重點實驗室,江蘇無錫 214122; 2.江南大學食品學院,江蘇無錫 214122)
谷朊是淀粉加工的重要副產(chǎn)品。為了提高谷朊產(chǎn)量和改善谷朊性質(zhì),在制備谷朊的過程中,向面漿中加入微生物轉(zhuǎn)谷氨酰胺酶(MTG)。結(jié)果表明,隨著酶量的增加,谷朊的蛋白含量提高了 3.98%。濕面筋的延伸性和回復(fù)性隨酶量的增加逐漸減小。通過谷朊蛋白質(zhì)分子羧基含量的測定和蛋白質(zhì)分子間相互作用力的分析,在MTG作用 100min后蛋白分子的羧基含量上升,蛋白質(zhì)之間疏水相互作用明顯減弱。研究表明,在微生物轉(zhuǎn)谷氨酰胺酶(MTG)作用下谷朊的性質(zhì)產(chǎn)生了明顯的變化。
谷朊,微生物轉(zhuǎn)谷氨酰胺酶(MTG),質(zhì)構(gòu),流變學特性,化學相互作用,羧基含量
谷朊粉是從小麥面粉中分離出的高蛋白聚合物,通常作為重要的食品添加劑、品質(zhì)改良劑應(yīng)用到現(xiàn)代食品當中[6]。流變性質(zhì)是谷朊的基本性質(zhì)之一,該性質(zhì)使得面團能夠制成面包、蛋糕、餅干和面條等。在濕面筋中,蛋白質(zhì)分子形成具有黏著性和彈性的面筋網(wǎng)絡(luò),使得面筋可以延伸和膨脹[16]。谷朊蛋白主要由具有較好延伸性的醇溶蛋白和具有較好彈性的麥谷蛋白構(gòu)成。谷朊蛋白的谷氨酰胺含量非常高,約為 35%,是微生物谷氨酰胺轉(zhuǎn)移酶(MTG)催化反應(yīng)的良好底物[7,10]。谷氨酰胺轉(zhuǎn)胺酶 [EC 2.3.2.13],是一種催化酰基轉(zhuǎn)移反應(yīng)的轉(zhuǎn)移酶[4],目前使用的多為微生物來源的谷氨酰胺轉(zhuǎn)移酶 (簡稱MTG)。MTG以肽鏈中谷氨酰胺殘基的γ-羧酰胺基作為酰基供體,當酰基受體為多肽鏈中賴氨酸的ε-氨基或伯胺基時,催化蛋白質(zhì)發(fā)生分子內(nèi),分子間或蛋白質(zhì)與小分子間的交聯(lián)反應(yīng);酰基受體為水時,谷氨酰胺殘基脫去氨基生成谷氨酸殘基,發(fā)生水解反應(yīng)。MTG常用于改善蛋白的功能性質(zhì),在食品工業(yè)具有廣闊的應(yīng)用前景。本實驗向面漿中添加MTG制取谷朊,考察了MTG對谷朊產(chǎn)量和性質(zhì)的影響,并對影響其性質(zhì)的因素進行了研究。
1 材料與方法
1.1 材料與設(shè)備
魯王牌特精粉 山東魯王集團有限公司;谷朊粉 東海糧油工業(yè) (張家港)有限公司;轉(zhuǎn)谷氨酰胺酶(2000U/g) 江蘇泰興一鳴精細化工有限公司。
LNK872型多功能快速消化器 江蘇省宜興市新宇科教儀器研究所;F W100型高速萬能粉碎機天津市泰斯特儀器有限公司;Glutograph-E面筋儀C.W.Brabender Instruments,Inc;AR1000 Rheometer T A Instruments Company;可見分光光度計 V-1800上海美普達儀器有限公司;熒光分光光度計Hitachi 650-60。
1.2 實驗方法
1.2.1 谷朊粉制取 采用面漿離心法 (Raisio法)[9],并改進為:面粉加水混合,靜止 30min,2500r/min離心 10min,沉淀物用 700mL自來水洗滌 5次,得濕谷朊,60℃干燥磨粉得谷朊粉。
1.2.2 谷朊蛋白質(zhì)含量測定 采用微量凱氏定氮法(GB5009-85),蛋白質(zhì)系數(shù)為 5.7。
1.2.3 面漿離心后上清液中蛋白含量測定 采用雙縮脲法[12]。
1.2.4 濕谷朊延伸性的測定 采用 Glutograph-E電子型面筋特性測定儀測定。載荷 150cm·g,濕面筋擠壓于兩平行波狀板之間,下板在在一恒定扭力下轉(zhuǎn)動,在 0~100s內(nèi)下板轉(zhuǎn)動一定的角度 (剪切角),得到面筋的延伸性 (Stretching),以及在 100~110s內(nèi)扭矩消失后,下板回轉(zhuǎn)一定角度,得到面筋的回復(fù)性(Relaxation),二者均以BU為單位表示。
1.2.5 面漿流變性測定 用 AR1000流變儀測定。40mm齒型平行板系統(tǒng),平行板間距 1mm,測量溫度為 35℃,根據(jù)樣品隨振蕩頻率(0.01~10Hz)的變化確定線性粘彈區(qū)。在線性粘彈區(qū)范圍內(nèi),固定振蕩頻率,測定貯能模量 G′和損耗模量 G″隨時間 (1h)掃描的變化。
測定前 20min和好面漿 (25g面粉加入 50mL水中),提前3min加入MTG酶液,調(diào)勻后加約1.5mL于兩平行板之間,測量時間為 1h。
1.2.6 蛋白質(zhì)分子間相互作用力測定 參照M.C.Go′mez-Guille′n[18]的方法。樣品處理過程為: 30.3g的谷朊粉分散于 500mL的 0.01mol/mL,pH=7的 Tris緩沖液中,添加 0.02496g的MTG,在反應(yīng)溫度為 25℃下反應(yīng) 4h。分別在 0、15、30、45、60、90、120、150、180、240min處取樣,100℃下 4min滅酶活。2500r/min離心 10min,棄上清,得樣品。
1.2.7 蛋白質(zhì)分子上羧基含量測定 參照 Kingsley K[14]等的方法。樣品處理過程為:用胰蛋白酶對谷朊進行水解,水解條件是:底物濃度為 5%,pH=8.5, 40℃水解 5h。取 50mL的水解物,添加 0.05g的MTG,反應(yīng)溫度為 25℃,調(diào) pH=7,進行反應(yīng)。分別在 0、15、30、45、60、90、120、180、240min處取樣, 100℃下 4min滅酶活,冷卻,得樣品。
2 結(jié)果與討論
2.1 MTG加入量對制得谷朊得率和蛋白含量的影響
按 0、6、25、100mg MTG/kg面粉的比例添加MTG,制面漿,2600r/min離心 10min。上清液用雙縮脲法測蛋白質(zhì)含量,沉淀制取谷朊粉,并分別取約0.5g進行微量凱氏定氮,結(jié)果如表 1。

表 1 MTG加入量對制得谷朊得率和蛋白質(zhì)含量的影響
由表 1可以看出,隨著酶量的增加,谷朊中粗蛋白含量顯著增加,MTG加入量為 100mg/kg面粉時蛋白質(zhì)含量增加了 3.98%。上清液中剩余可溶性蛋白的量隨著酶量的增加而逐漸減少,這表明在MTG的作用下,有部分可溶性的蛋白交聯(lián)到了面筋蛋白質(zhì)上,從而引起上清液中測得的蛋白質(zhì)含量減少[19],谷朊中蛋白質(zhì)含量增加,并且在較高酶量的作用下,交聯(lián)反應(yīng)更加明顯。由表 1還可以看出,谷朊得率隨著酶量的增加而下降,這是由于在MTG作用下,谷朊的粘性有所下降,淀粉易于分離,但影響谷朊全部回收,導(dǎo)致粗蛋白含量的增加,得率有所下降。
2.2 MTG加入量對谷朊質(zhì)構(gòu)的影響
按 0、1、3、6、12、25、100mg/kg面粉的比例添加MTG,制取濕面筋,用 Grutograph-E電子型面筋特性測定儀測定面筋的延伸性和回復(fù)性。
由圖 1可以看出,濕面筋的延伸性和回復(fù)性隨酶量的增加逐漸減小。當MTG加入量為 100mg/kg面粉時,濕面筋的延伸性由原來的 591BU急劇下降到98BU,回復(fù)性也相應(yīng)的由91BU下降到27.5BU,這同時反映出濕面筋的抗延伸性阻力隨MTG添加量的增多是逐漸增大的。并且在濕面筋流變性質(zhì)測量過程中,發(fā)現(xiàn)濕面筋的黏性隨MTG加入量的增加逐漸下降。這些數(shù)據(jù)表明,在反應(yīng)發(fā)生的過程中面筋蛋白中的醇溶蛋白部分交聯(lián)到了麥谷蛋白上,導(dǎo)致濕面筋的延伸和恢復(fù)性下降,抗延伸阻力增加和粘性的降低[19]。

圖 1 MTG加入量對谷朊延伸性和回復(fù)性的影響
2.3 MTG加入量對面漿流變性質(zhì)的影響
測定前20min和好面漿,提前3min加入酶液,三組面漿加酶量分別為:0、25、100mg/kg面粉,調(diào)勻后加約 1.5mL面漿于兩平行板之間,測量時間為 1h。
G′是儲能模量,代表樣品的彈性部分,G′越大代表樣品的彈性越好;G″是損耗模量,代表樣品的粘性部分,G″越大代表樣品的粘性越好;tgδ=G″/G′,δ越大則粘性成分占優(yōu)勢,體系表現(xiàn)為流體的特性,δ越大則體系表現(xiàn)為固體的特性。由圖 2和圖 3可以看出,3種MTG添加量的面漿動態(tài)粘彈曲線 G″(損耗模量)始終大于 G′(貯能模量),體系以黏性為主。隨時間的變化,MTG用量為 100mg/kg面粉的面漿G″和 G′升高都是最大,說明該用量下MTG對于面漿中蛋白的作用程度最大,面漿的黏性和彈性均增大。MTG用量為 25mg/kg面粉時,G′、G″變化在高酶量和無酶的曲線之間,但是圖 4所顯示出的δ值卻是最小。分析其原因:此種現(xiàn)象與MTG催化蛋白質(zhì)所發(fā)生兩種反應(yīng)密切相關(guān)。這兩種反應(yīng)分別為:一是交聯(lián)蛋白質(zhì),使面筋網(wǎng)絡(luò)增強,使得 G′增大;二是催化蛋白質(zhì)谷氨酰胺殘基脫去氨基生成谷氨酸殘基,該反應(yīng)使得蛋白質(zhì)的帶電性發(fā)生改變,導(dǎo)致整個面漿體系不穩(wěn)定,使得 G″增大[12]。高酶量作用下,蛋白質(zhì)間的交聯(lián)反應(yīng)很明顯。在交聯(lián)反應(yīng)的后期,水代替伯胺成為酰基受體,發(fā)生脫酰胺反應(yīng),脫酰胺反應(yīng)也很明顯,所以 G″和 G′升高都是最大。在MTG用量為25mg/kg面粉時,催化蛋白質(zhì)間的交聯(lián)反應(yīng)占主導(dǎo)地位,脫酰胺反應(yīng)較弱,導(dǎo)致 G′、G″變化在高酶量和無酶的曲線之間,但是δ值卻是最小,此時面漿的凝膠性最好。
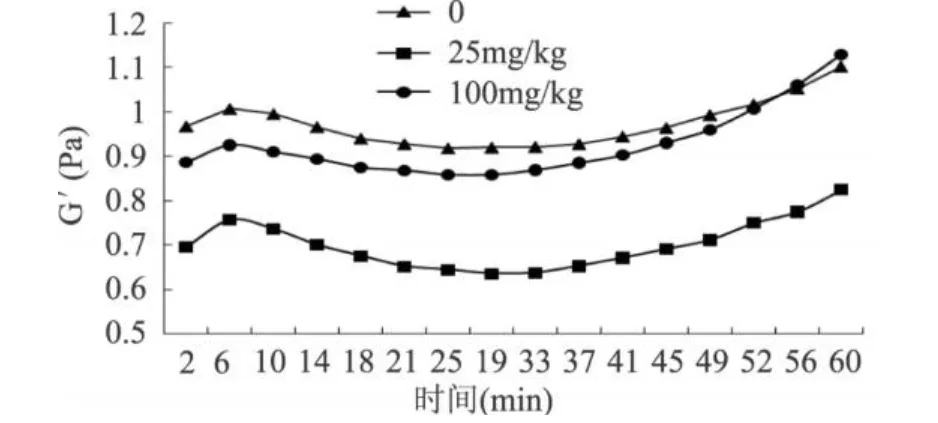
圖 2 MTG加入量對面漿貯能模量(G′)的影響
2.4 MTG作用過程中蛋白質(zhì)間相互作用力的變化
MTG對谷朊作用的過程中,會對谷朊蛋白分子間的作用力產(chǎn)生影響,由圖 5可以看出,隨著酶反應(yīng)的進行,谷朊蛋白質(zhì)間的氫鍵和離子鍵沒有發(fā)生明顯的變化。在酶作用 100min后,蛋白質(zhì)間疏水的相互作用明顯下降。從MTG作用機理來看,隨著交聯(lián)反應(yīng)的進行,體系中伯胺的濃度會逐漸降低,交聯(lián)反應(yīng)的后期,水會代替伯胺成為酰基受體,導(dǎo)致水解反應(yīng)發(fā)生,谷氨酰胺殘基脫去氨基生成谷氨酸殘基,蛋白質(zhì)分子上的羧基含量增加。由圖 6可看出,在以谷朊水解物為底物,MTG催化反應(yīng)發(fā)生 100min后,體系中的羧基含量明顯上升。所以MTG催化谷朊反應(yīng)時,在 100min后,有水解反應(yīng)發(fā)生,蛋白質(zhì)分子上的羧基含量上升,導(dǎo)致蛋白質(zhì)間的靜電排斥作用加大,疏水相互作用降低,從而對谷朊的性質(zhì)產(chǎn)生影響。

圖 3 MTG加入量對面漿損耗模量(G″)的影響
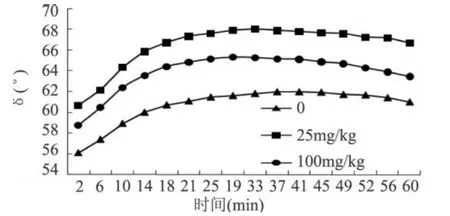
圖 4 MTG加入量對面漿凝膠性(δ)的影響
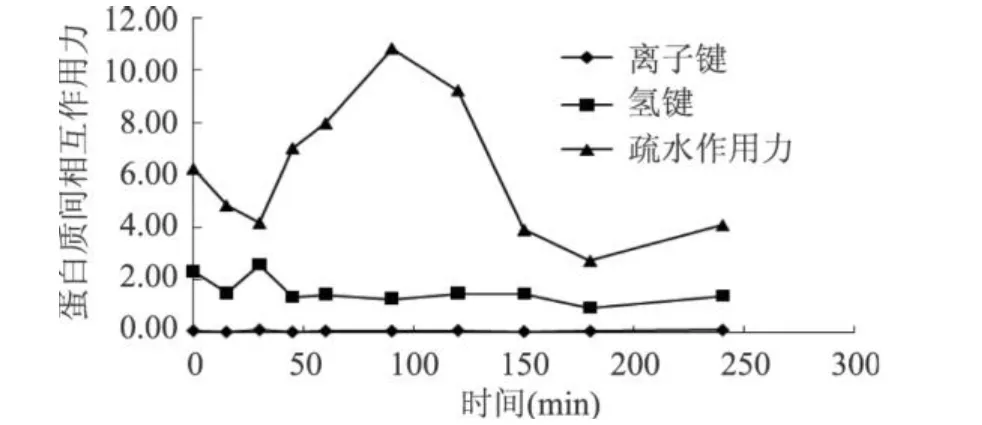
圖 5 MTG作用時間對谷朊蛋白質(zhì)間相互作用力的影響
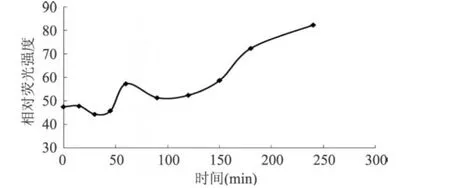
圖6 羧基含量隨MTG作用時間的變化
3 結(jié)論
微生物轉(zhuǎn)谷氨酰胺酶作用下會發(fā)生兩種反應(yīng),交聯(lián)反應(yīng)和脫酰胺反應(yīng)。這兩種反應(yīng)共同對制得的谷朊的蛋白含量、質(zhì)構(gòu)和流變性質(zhì)產(chǎn)生影響,使得隨著酶濃度增加,制得的面筋粗蛋白含量增加,黏性和延展性顯著下降,抗延伸性阻力呈增強趨勢。在交聯(lián)反應(yīng)的后期,水代替伯胺成為酰基受體,發(fā)生水解反應(yīng),導(dǎo)致體系中蛋白分子上的羧基含量增加,蛋白質(zhì)間疏水相互作用力降低。
[1]周大捷,馬凡賽 .小麥活性谷朊粉和淀粉的生產(chǎn)技術(shù)-小麥深度加工的開端[J].糧食與飼料工業(yè),1996(2):34-37.
[2]吳紅奎 .漫談谷朊粉 [J].西部糧油科技,1999(24): 34-35.
[3]史新慧,王蘭,鄒風羽,等 .小麥面筋蛋白改性的研究[J].鄭州糧食學院學報,2000,3(21):27-31.
[4]周楠迪,陳堅,鄭美英,等.谷氨酰胺轉(zhuǎn)移酶的功能性質(zhì)及其在食品中的應(yīng)用方法[J].中國食品添加劑,2000(1):55-59.
[5]張洪淵,萬海清 .生物化學 [M].北京:化學工業(yè)出版社,2001.
[6]張紅印,王蘭,席與芳,等 .小麥面筋蛋白的乙酰化改性[J].無錫輕工大學學報,2002,5(21):239-243.
[7]周惠明 .谷物科學原理 [M].北京:中國輕工業(yè)出版社,2003.
[8]趙冬艷,王金水,劉宇宸 .濕熱處理提高谷朊粉乳化性的研究[J].糧食與飼料工業(yè),2003(4):45-47.
[9]趙秋紅,莫重文 .小麥谷朊與淀粉制取研究[J].西部糧油科技,2003(1):37-40.
[10]王璋,許時嬰,湯堅 .食品化學[M].北京:中國輕工業(yè)出版社,2006.
[11]陳海英 .谷氨酰胺轉(zhuǎn)胺酶改性谷朊粉及其機理的研究[D].沈陽:沈陽農(nóng)業(yè)大學,2006.
[12]孔祥珍,周惠明,王洪燕 .堿性蛋白酶水解小麥面筋蛋白的研究[J].食品工業(yè)科技,2006(9):104-116.
[13]劉鳳華,席慧,高音 .蛋白質(zhì)交聯(lián)的研究進展[J].生物技術(shù)通報,2004(4):10-13.
[14]Kingsley K Agyarea, Youling L Xiong, Kwaku Addo.Influence of salt and pH on the solubility and structural characteristicsof transglutaminase- treated wheat gluten hydrolysate.[J]Food Chemistry,2008,107:1131-1137.
[15]Kong Xiangzhen,Zhou Huiming,Qian Haifeng.Enzymatic preparation and functional properties of wheat gluten hydrolysates [J].Food Chemistry,2006,5386:1-6.
[16]L Day,M A Augustin,I L Batey,et al.Wheat-gluten uses and industry needs[J].Food Science&Technology,2006(17): 82-90.
[17]M Motoki,K Seguro.Transglutaminase and its use for food processing[J].Food Science&Technology,1998(9):204-210.
[18]M C Go′mez-Guille’n,A JBorder1’as,P Montero.Chemical Interactions of Nonmuscle Proteinsin the Network of Sardine(Sardina pilchardus)Muscle Gels[J]. Lebensm-W iss u-Technol.
[19]Nicole Bauer,Peter Koehler,HerbertW ieser,et al.Studies on Effects of Microbial Transglutaminase on Gluten Proteins of Wheat.Ⅰ.Biochemical Analysis[J].Cereal Chem,80(6): 781-786,787-790.
Effect of m icrobial transglutam inase on texture and rheologicalproperties of wheat gluten
CHAO L i-jing1,WANGM iao2,*,SUN Gui-peng2
(1.State KeyLaboratory of Food Science and Technology,Jiangnan University,Wuxi 214122,China; 2.School of Food Science and Technology,Jiangnan University,Wuxi 214122,China)
W hea t g luten is an i mp ortant by-p roduc t of s ta rch m anufac turing p rocedure.M ic rob ia l transg lutam inase (M TG)was added into flour susp ens ion to ge t g luten.The inc reas ing concentra tions of the M TG led to a inc rease of p rote in content in g luten and a dec rease of extens ion and recove ry of we t g luten.By m easuring the ca rboxyl g roup on p rote in and ana lyzing inte rac tions be tween p rote in m olecules,it can be found tha t the content of the ca rboxyl g roup inc reased and the hyd rop hob ic inte rac tion be t ween p rote ins dec reased afte r100m in due to hyd rolys is ca ta lyzed by M TG.The refore both c ross-link and hyd rolys is ca ta lyzed by M TG influenced the p rote in content and qua lity of g luten.
whea t g luten;M TG;texture;rheology p rop e rties;chem ica l inte rac tions;ca rboxyl g roup content
TS201.2
A
1002-0306(2010)03-0327-04
2009-06-01 *通訊聯(lián)系人
晁麗靜(1985-),女,碩士研究生,從事酶反應(yīng)以及生物技術(shù)在食品中應(yīng)用方面的研究。
國家自然科學基金面上項目(20776063)。
