波紋唇魚消化系統的形態解剖與腸道上皮的掃描電鏡觀察
2010-12-28 04:48:36王永波陳國華駱劍王珺黃宗文尹紹武張本
海洋通報 2010年2期
王永波,陳國華,駱劍,王珺,黃宗文,尹紹武,張本
(1. 海南大學海洋學院 海南大學熱帶生物資源教育部重點實驗室 海南省熱帶水生生物技術重點實驗室,海南 海口 570228;2. 海南省水產研究所,海南 海口 570206)
波紋唇魚消化系統的形態解剖與腸道上皮的掃描電鏡觀察
王永波1,2,陳國華1,駱劍1,王珺1,黃宗文1,尹紹武1,張本1
(1. 海南大學海洋學院 海南大學熱帶生物資源教育部重點實驗室 海南省熱帶水生生物技術重點實驗室,海南 海口 570228;2. 海南省水產研究所,海南 海口 570206)
對波紋唇魚消化系統的解剖構造進行了研究,并利用掃描電鏡對其腸道上皮組織進行觀察,結果如下:波紋唇魚為肉食性無胃魚類,消化道全長極短,頜齒和咽齒都很發達,腸道極粗,比腸長為0.426;肝臟分三葉,呈楓葉狀,胰臟彌散性分布在肝臟組織中,肉眼不易分辨;掃描電鏡觀察發現小腸前部細胞界限不明顯,分泌孔較多,微絨毛密而長短不齊;小腸后部細胞界限也不明顯,但分泌孔明顯減少,微絨毛比小腸前部更密且很平整;直腸細胞為多角形,分泌孔比小腸少,微絨毛較稀疏,且長短不一。
波紋唇魚;消化系統;形態學;掃描電鏡
魚類的消化系統是由消化管以及連附于消化管附近的各種消化腺組成,各器官直接或間接地參與食物的消化和吸收,從外界獲取營養的能力關系到魚類生長、發育乃至繁殖等重要的生命活動。魚類消化系統形態解剖學方面的研究開始得較早,迄今為止,學者們已對多種魚類消化系統的形態解剖學進行過研究[1-8],但有關肉食性海水無胃魚類的報道不多。
波紋唇魚(Cheilinus undulates Rüppell)俗稱蘇眉、拿破侖,隸屬鱸形目、隆頭魚科、唇魚屬。該魚為海水肉食性無胃魚類,主要以軟體動物、魚、海膽、腹足動物、瓣鰓動物、甲殼動物以及其它的無脊椎動物為食[9-12]。它是體型最大的珊瑚礁魚類之一,成體體長可達200 cm,目前發現的最大個體達250 cm,重191 kg。主要分布于非洲東岸,紅海及從印度洋至太平洋中心,我國主要分布在南海與東海的南部海域,在海南、香港、臺灣諸島都有分布。波紋唇魚的魚唇一直是亞洲人盤中的美味佳肴,且其魚肉質地鮮嫩,是目前價格最高的食用魚種之一,具有極高的商品開發價值。近幾年因過渡捕撈等原因導致該魚瀕臨絕種。2004年,世界自然基金會公布了10種瀕臨滅絕的生物名單,波紋唇魚名列第二;同年還被列入IUCN(國際自然保護聯盟)瀕危動物紅皮書;2005年被列入CITES《瀕危野生動植物種國際貿易公約》附錄Ⅱ。
目前對該魚的研究國外做了大量的工作[9-18],而國內較少[19-21],研究工作基本上集中在生態調查和種群保護領域,未見到該魚消化系統形態解剖學方面的研究。在自然條件下,波紋唇魚在達到性成熟之前有一個快速生長的階段,而在人工養殖條件下,發現其生長緩慢,為此我們開展了消化系統解剖學等方面的研究,以期為人工養殖提供基礎理論的指導,也為該魚的生物學積累資料。
1 材料與方法
1.1 實驗材料
試驗魚取自海南省三亞市紅沙港的海水養殖網箱,該批魚是當地漁民從南沙群島、西沙群島等南海海域捕獲的幼魚經人工養殖的亞成體,3齡,主要喂以魷魚、對蝦、蟹、小雜魚等。選擇體表無傷、健康的個體13尾(分兩次取樣,第一次10尾,第二次3尾)。樣本的平均體重322.86 g,平均體長228.9 mm。
1.2 實驗方法
解剖觀察前,測量魚體重,體長。打開腹腔,原位觀察各器官的位置。測定腸道全長,腸道的直徑等指標,精確到1 mm,計算消化道指數(比腸長 = 腸道長/體長;比腸重 = 腸重/體重;比肝胰臟重 = 肝胰臟重/體重 × 100%),然后將整個腸道翻轉,沖洗腸粘膜表面,在解剖鏡下觀察腸道粘膜皺褶的表面形態,排列和分布。
取3尾波紋唇魚的腸道粘膜作掃描電鏡觀察。取粘膜塊,大小約1 cm × 1 cm,經PBS磷酸緩沖液清洗,4% 戊二醛固定,日立S-3000N型掃描電鏡觀察并照相。從小腸前部(腸道第一個自然折點之前部分),小腸后部(腸道第一個自然折點和鮑氏瓣之間部分),直腸(鮑氏瓣到肛門)等三個部位取材。
2 結 果
2.1 消化道指數
測定了10尾波紋唇魚的消化道指數,結果見表1。波紋唇魚為肉食性海水無胃魚類,消化道結構簡單,食道之后即為小腸,小腸在腹腔內有一個彎曲折疊,呈“ ”型。體長245 ~ 295 mm的波紋唇魚,比腸長平均為0.426。大部分的魚類比腸長都在1.2 ~ 6.0之間[23],已有報道的海水肉食性魚類的比腸長都比波紋唇魚大,如大菱鲆 (Scophthalmus maximus)[22]的比腸長為0.75,鱈魚 (Gadus morhua) 為0.966,大銀魚(Protosalanx hyalocranius) 為0.69,可見在肉食性魚類中波紋唇魚的比腸長是較小的。但波紋唇魚的小腸很粗,在正常攝食條件下和饑餓狀態下的腸道直徑,見表2。從表2可知,波紋唇魚腸道的直徑是無胃魚泰山璃霖魚 (Varicorhinus macrolepi) 和鯽魚[8]的數倍。波紋唇魚的平均比腸重為0.019,與肉食性魚類大菱鲆的比腸重0.015相差不大,但是大菱鲆是有胃魚,總的比消化道重比波紋唇魚的大。波紋唇魚的胰臟呈彌散性分布在肝臟之中,肉眼很難分辨出胰臟組織,所以稱為“肝胰臟”。波紋唇魚的比肝胰臟重為1.5%,符合隆頭魚科魚類比肝重的特征[23]。
2.2 消化道的形態
波紋唇魚的口端位,下頜長于上頜,屬上位口(圖版I-1)。前上頜骨不固著于上頜骨,口可向前伸出。口咽腔較大,口的最外側為唇,唇較厚、發達,稍掀起唇可見上下頜各具1行尖銳而鋒利的頜齒,呈錐形,上下頜最前面分別具1對大的犬齒,在13尾標本上都發現這對犬齒在上頜左小右大,在下頜左大右小,合攏時正好互補(圖版I-2,3)。波紋唇魚的咽齒十分發達,左右上咽齒分開,各由多行顆粒狀齒愈合成骨板狀的三角形,下咽齒也由多行顆粒狀齒愈合成骨板狀,整體呈“T”型(圖版I-4)。
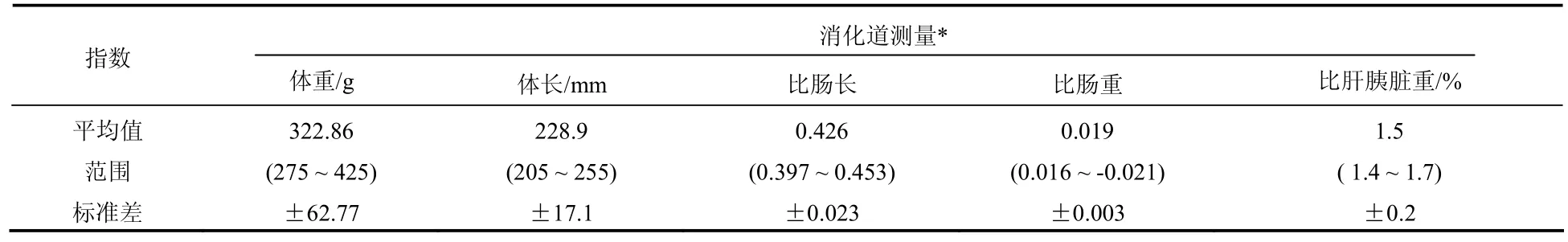
表1 波紋唇魚消化道指數Tab. 1 Alimentary tract exponent of Cheilinus undulatus
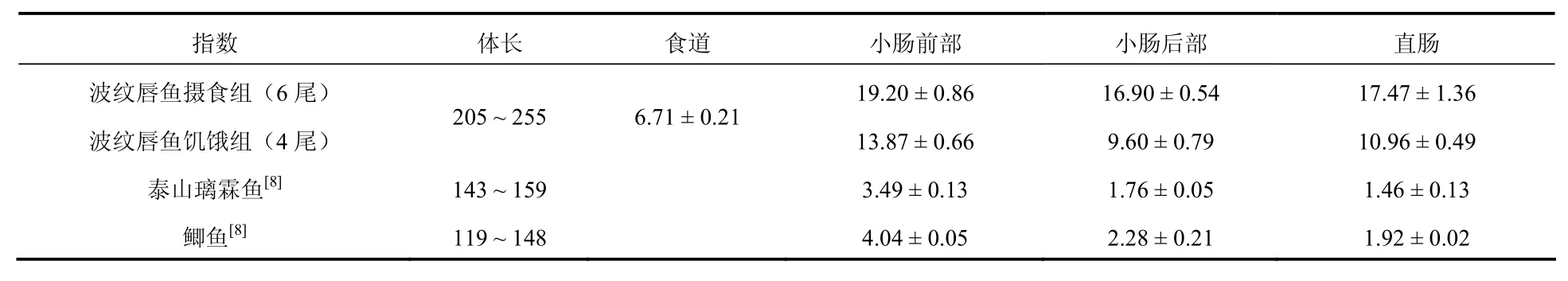
表2 波紋唇魚消化道的直徑 (單位:/mm)Tab. 2 Diameter on digestive tract of Cheilinus undulates
用剪刀沿口角的兩側剪至鰓蓋骨后方,打開口咽腔,可見口咽腔上、下部均為銀白色,光滑,前部生長有發達的口腔瓣。舌為長條形,前端圓,游離,銀白色(圖版I-2,3)。
波紋唇魚第一至第三對鰓弓上具全鰓,第四對鰓弓上具半鰓,與鰓相對,前三對鰓弓上各有二列鰓耙,第四對鰓弓上僅一列鰓耙,且鰓耙長度僅約為前三對的一半,屬多列不對稱型鰓耙,鰓耙為白色的錐形狀,十分堅硬,中間長,兩邊短(圖版I-5)。
波紋唇魚的食道短,體長270 mm 左右的個體食道長約10 mm,直徑約10 mm(圖版I-7)。食道壁厚,前部緊接著口咽腔,后端與小腸前部相連(波紋唇魚無胃)。食道直徑約為小腸前部直徑的1/2 ~ 1/3。
波紋唇魚的腸較短,腸道長與體長的比為0.426,腸在腹腔中向后行至腹腔后部,折向前,再折向后,形成一個盤曲,整個消化道呈“ ”型;可分為小腸前部、小腸后部和直腸,食道末端至小腸回繞的彎曲之間為前部,呈紫紅色,是最粗的一段,體長270 mm左右的個體,直徑約20 mm。自小腸回繞段至直腸為小腸后部,較細,呈粉紅色。小腸最后有一個狹窄處,即小腸和直腸的分界,此處有一瓣膜(鮑氏瓣),鮑氏瓣附近的小腸略帶黃色。直腸很短,直徑較粗,呈淺紅色,到肛門處明顯變細(圖版I-7)。
2.3 消化腺的形態
打開波紋唇魚的左側腹腔,能看到肝臟(圖版I-6)。肝臟分三葉,呈楓葉狀(圖版I-8),左、右兩葉較小,腹側葉較大。肝臟呈黃褐色,背面清楚可見紅色的呈葉脈狀的放射型血管;腹面被很多深溝分割成塊狀,深溝內有白色的膽管,膽管像植物的根系一樣向肝總管集中,整個肝臟呈放射狀。波紋唇魚的左葉,腹側覆蓋了小腸前部,背側緊貼鰾;右葉在波紋唇魚身體的右側,腹側也覆蓋小腸前部,正好填充了魚鰾和小腸前部之間的間隙;腹葉的大部分位于腹腔左側,從腹側包裹著小腸前部,并覆蓋了整個小腸后部。
波紋唇魚的膽囊較大,橢球狀,藍綠色,位于肝腹側葉和右葉間,大部被肝腹側葉所覆蓋。肝總管和總膽管經乳頭從腹側通入小腸前部(圖版I-9)。
波紋唇魚沒有單獨的胰臟,胰組織呈彌散性的分布在肝組織中,肉眼很難分辨。
2.4 消化道粘膜的的掃描電鏡觀察
消化道粘膜褶皺。在低倍鏡下,食道褶皺的走向和身體平行,呈整齊的縱向排列;小腸前部(圖版II-10)、小腸后部和直腸(圖版II-11)的粘膜褶走向多樣,有“Z”型,“V”型、“S”型等走向的粘膜褶皺,各段無明顯差別。
消化道各部的細胞界限。小腸前部和后部的細胞界限較模糊,基本分辨不出來(圖版II-12、13);直腸部分的細胞界限清楚,呈多角形(圖版II-14)。
消化道各部的分泌孔形態和密度。腸道各部均有較多的分泌孔,但數量和形狀上有差異,小腸前部的分泌孔密度最大(圖版II-12),小腸后部和直腸較小腸前部相比,分泌孔的密度明顯變小(圖版II-13、14)。分泌孔呈圓型或橢圓型,但小腸前部和后部分泌孔的四周比較平滑,有些可見含有顆粒狀的分泌物,直腸的分泌孔四周不平滑,有很多縫隙。
腸道各部的微絨毛的形態和數量。小腸前部的微絨毛較密,但不整齊,多數長短不一(圖版II-15);小腸后部的微絨毛比前腸密,較平整,長短均一(圖版II-16);直腸的微絨毛較稀疏,長短不一,且相鄰處有較深的溝(圖版II-17)。
3 討 論
3.1 波紋唇魚腸道的分段
有關無胃魚類腸道的分段問題在鯉科魚類中研究較多,林浩然[2]認為食道之后便是腸,沒有分段的必要;而其他學者大都把腸道分為前、中、后腸三部分[4,5]。波紋唇魚屬隆頭魚科,雖然也無胃,但消化道的形態和鯉科的魚類還是有很大的不同。作者通過對波紋唇魚消化道解剖和掃描電鏡觀察,發現消化道各段有很大差異,這些形態上的差別,暗示其機能上的不同,故將波紋唇魚的消化道分為三部分,即小腸前部、小腸后部、直腸。小腸前部為腸道第一個自然折點之前的部分,這個部分的顏色呈紫紅色,明顯比小腸后部顏色深,直徑比小腸后部大,分泌孔密度也比小腸后部大;小腸后部,即腸道第一個自然折點和鮑氏瓣之間的部分;直腸部分為鮑氏瓣到肛門的部分。
3.2 消化系統構造與機能
波紋唇魚屬于肉食性無胃魚類,主要以軟體動物(腹足類、瓣鰓類、頭足類),魚類、海膽、甲殼類動物和其它的無脊椎動物為食[9-12],當食物充足時更喜好攝食甲殼類。從已報道的波紋唇魚食性資料來看,其經常攝食一些較堅硬的動物(如螃蟹等),這和它的消化系統的結構是相適應的。波紋唇魚的頜齒非常發達,在上下頜的前端還各具有2個犬牙狀的齒,有利于咬住比較堅硬動物的外殼;口咽腔較大,有利于攝取比較大的食物;該魚的咽齒為臼齒狀,有利于磨碎貝類、甲殼類等堅硬外殼。觀察飼養在網箱的波紋唇魚,該魚喜好打斗,常利用上下頜前端犬牙攻擊對手,使對方受傷。
波紋唇魚的食道壁很厚,管腔內具有多列較高的縱行粘膜褶,有利于吞咽大而較堅硬的食物。波紋唇魚無胃,食物從食道直接進入小腸。波紋唇魚的腸道極短,比腸長為0.426,與目前所見報道比腸長最小的飛魚 (0.41 ~ 0.55) 相當。比腸長小、腸道結構簡單(只有一個盤曲)的特點與波紋唇魚的食性相關,該魚的食糜團內含有很多所攝食動物的堅硬骨骼碎片,食糜團在消化道內不需要作長距離或復雜的移動是有利的。波紋唇魚的前腸很粗,正常攝食情況下直徑可達18 mm(體長270 mm的個體),是無胃魚泰山璃霖魚(體長300 mm的個體)前腸直徑的5倍,有效地增加了腸道的容量。在小腸和直腸的交接處的鮑氏瓣可以防止食糜團的倒流,還可以保證食糜團在小腸內有充足的消化和吸收時間。
腸道的掃描電鏡觀察發現小腸前部的分泌孔最多,小腸后部和直腸較少,說明小腸前部分泌粘液的功能較強,當食糜團由食道進入腸道后和較多的腸道分泌物相混合,有助于食物的消化和吸收,對腸壁也有潤滑的作用,起到保護腸壁的作用。腸道粘膜的微絨毛主要作用是增加物質消化和吸收的面積,波紋唇魚腸道的微絨毛總體來看小腸內的長而密,直腸的較稀疏,這與齊口裂鰒魚 (Schizothorax prenanti)[24]、烏鱧(Ophicephalus argus)[25]等腸道內微絨毛的分布情況相一致,可以推斷,波紋唇魚的小腸是食物主要的消化和吸收的場所。
波紋唇魚的人工養殖開展極少,根據該魚的消化道短的特點,建議飼養波紋唇魚采取少量多次的投喂方式,可能有利于生長。
[1] 秉志. 鯉魚的解剖 [M]. 北京: 科學出版社, 1960.
[2] 林浩然. 五種不同食性鯉科魚的消化道 [J]. 中山大學學報, 1962(3): 65-78.
[3] 孟慶聞, 蘇錦祥. 白鰱的系統解剖 [M]. 北京: 科學出版社, 1960: 77-82.
[4] 喻子牛, 孫曉瑜, 孫世春. 真鯛消化道的組織學和形態學研究 [J]. 水產學報, 1997, 21(2): 113-119.
[5] 潘黔生, 郭廣生, 方之平, 等. 6種有胃真骨魚消化系統比較解剖的研究 [J]. 華中農業大學學報, 1996, 15(5): 463-468.
[6] 孟慶聞. 魚類比較解剖學 [M]. 北京: 科學出版社, 1987: 80-131.
[7] 孟慶聞. 魚類學 [M]. 上海: 科學技術出版社, 1989: 201-297.
[8] 張金花, 王樹迎. 泰山蠕霖魚腸道的解剖研究 [J]. 山東農業大學學報(自然科學版), 2003, 34(4): 579-581.
[9] Randall J E, Head S M, Sanders A P L. Food habits of the giant humphead wrasse (Labridae) [J]. Environ. Biol. Fishes, 1978, 3: 235-238.
[10] Myers R F. Micronesian Reef Fishes [M]. 2nd ed. Coral Graphics, Guam, 1991, 298.
[11] Pogonoski J J, Pollard D A, Paxton, J R. Conservation Overview and action plan for Australian threatened and potentially threatened marine and estuarine fishes [M]. Environment Australia, Canberra, 2002: 373.
[12] Choat J H, Davis C R, Ackerman J L. Age structure and growth in a large teleost, Cheilinus undulatus, with a review of size distribution in labrid fishes[J]. Marine Ecology Progress Series. 2006. 318: 237-246.
[13] Sadovy Y, Kulbicki M, Labrosse P, et al. The humphead wrasse, Cheilinus undulatus: synopsis of a threatened and poorly known giant coral reef fish[J]. Reviews in Fish Biology and Fisheries 2004,13: 327-364.
[14] Sluka R D. Humphead wrasse (Cheilinus undulates) abundance and size structure among coral reef habitats in maldies [J]. Atoll Res. Bull, 2005. 538:189-198.
[15] Choat J H, Bellwood D R. Wrasses and parrotfishes. In: Paxton J R, Eschmeyer W N (eds.), Encyclopedia of Fishes. University of New South Wales Press, Sydney, 1994: 211–215.
[16] Dorenbosch M., Grol M.G.G., Nagelkerken I., et al. Seagrass beds and mangroves as potential nurseries for the threatened Indo-Pacific humphead wrasse, Cheilinus undulatus and Caribbean rainbow parrotfish, Scarus guacamaia [J].Biological conservation 2006,129:277-282.
[17] Olivier Chateau, Laurent Wantiez. Site fidelity and activity patterns of a humphead wrasse, Cheilinus undulatus (Labridae), as determined by acoustic telemetry [J]. Environ Biol Fish, 2006: 187-196.
[18] Murdjani, M. Marine fish culture, development and status in Indonesia. In: Proceedings of the Workshop on Aquaculture of Coral Reef Fishes and Sustainable Reef Fisheries, Kota Kinabalu, Sabah, Malaysia. Network of Aquaculture Centers in Asia, Bangkok, December 1996: 53–59.
[19] 沈世杰, 臺灣魚類志 [M]. 臺北:臺灣大學動物學系, 1993: 444-456.
[20] 張賀真, 王愛麗, 劉郁. 蘇眉魚中毒的急救護理 [J]. 罕少疾病雜志, 2005, 12(4): 28-29.
[21] 李桂英, 馬曉華. 32例蘇眉魚中毒患者的急救與護理 [J]. 現代護理, 2005, 11(10): 781-782.
[22] 王海英. 大菱鲆主要消化酶—蛋白酶.脂肪酶.淀粉酶的研究 [D]. 中國海洋大學優秀博士論文. 2006
[23] 尾崎久雄. 魚類消化生理(下冊) [M]. 上海: 上海科學技術出版社, 1985: 361-378.
[24] 方靜, 謝林, 李遜, 等. 齊口裂腹魚消化道粘膜上皮的掃描電鏡觀察 [J]. 水生生物學報,1995, 19(2): 188-189
[25] 李玉和, 郭淑華. 烏鱧消化道粘膜上皮的掃描電鏡研究 [J]. 解剖學報, 1992, 98-100.
Morphology and anatomy on digestive system of Cheilinus undulates Rüppell with scanning electron microscopy of intestinal tract
WANG Yong-bo1,2, CHEN Guo-hua1, LUO Jian1, WANG Jun1,HUANG Zong-wen1, YIN Shao-wu1, ZHANG Ben1
(1. College of Ocean, Hainan University; Key Laboratory of Tropic Biological Resources of Ministry of Education; Hainan Province Key Lab for Tropical Hydrobiology and Biotechnology; Haikou 570228, China; 2. Hainan Provincial Fisheries Research Institute, Haikou 570206, China)
In this study, the digestive system of Cheilinus undulates Rüppell was dissected and observed. The epithelial tissue of intestinal tract was observed using scanning electron microscope (SEM). The results are as follows:Cheilinus undulates Rüppell are predacity agastria fish; They have extremely short alimentary tracts and strong intestinal tracts; The ratio of intestine size to intestine length is 0.426; Jaw teeth and pharynx teeth are extreme upgrowth; Liver is divided into three parts, shaped like maple leaf; Pancreas is dispersed in liver tissue, hardly recognized by eyes. Cell boundary of anterior small intestine is unclear observed by SEM, many secretion pits are found, microvilli are dense but the length is uneven; Cell boundary of posterior small intestine is not distinctive either,but the number of secretion pits reduces obviously, and microvilli are denser and even; Recturn cells are polygonal cells, secretion pit number is smaller than small intestine, and microvilli are sparse and uneven.
Cheilinus undulates Rüppell; Digestive system; Morphology; SEM
S917
A
1001-6932(2010)02-0199-07
2009-04-13;
2009-09-28
國家“十一五”863計劃現代農業技術領域“海水養殖種子工程”重大項目——名貴海水魚類苗種規模化繁育技術(2006AA10A414);“十一五”國家科技支撐計劃重點項目-南海區主要海水養殖種類種質保存與評價技術(2007BAD29B03)
作者介紹:王永波(1983-),男,碩士,主要從事魚類人工繁殖與育種方面的研究。電子郵箱:wangyongbo211@163.com
陳國華,教授,電子郵箱:chguh@hainu.edu.cn
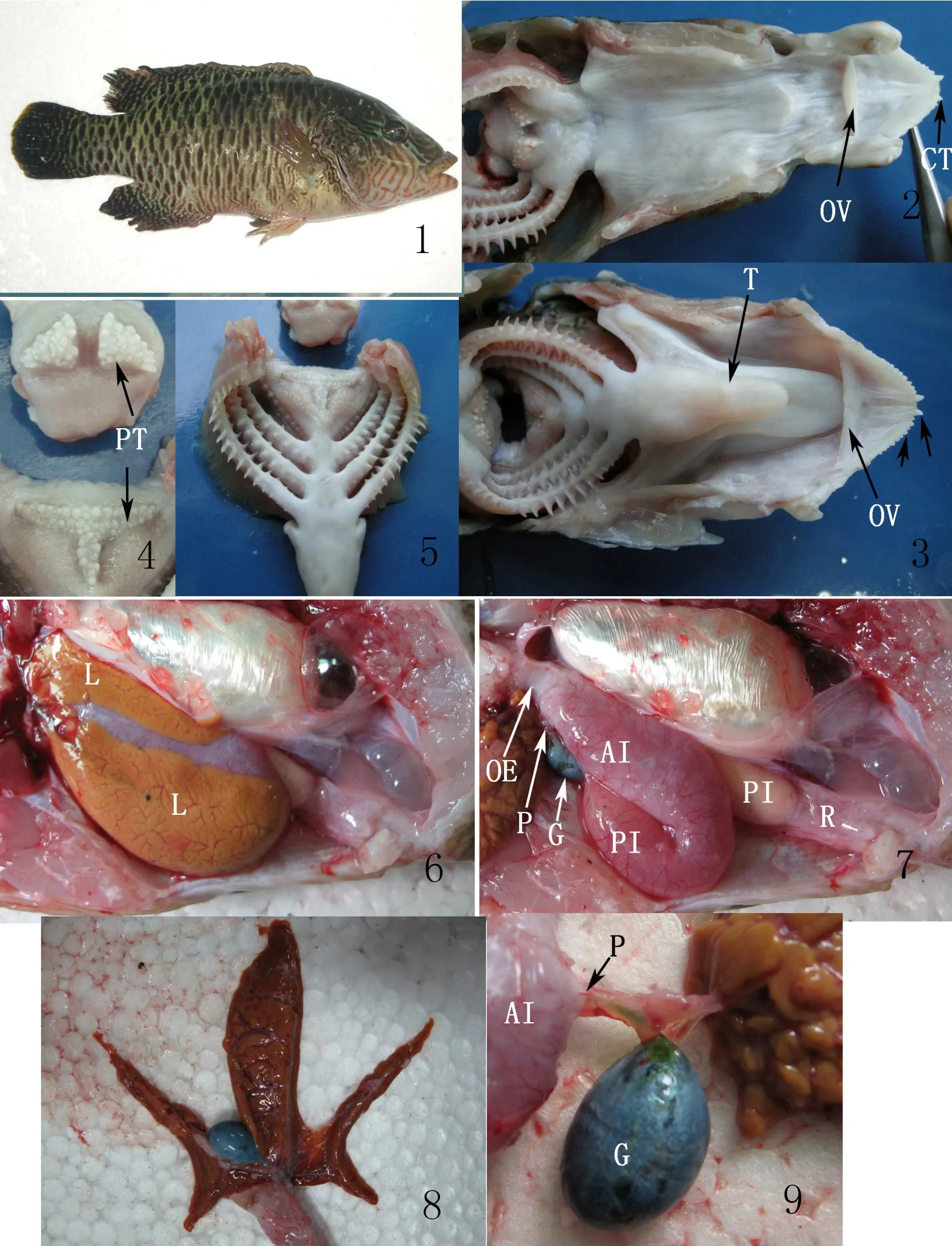
圖版I

圖版II
