健康成人棕色脂肪組織的功能
2011-01-16 05:32:50基爾西
糖尿病天地(臨床) 2011年2期
基爾西
健康成人棕色脂肪組織的功能
基爾西
在冷環境刺激下,我們利用正電子發射計算機斷層掃描(PET)檢測出健康受試者(5名)被激活的棕色脂肪組織,位于下頸部和鎖骨上部之間,還發現此處的葡萄糖攝取量增加。我們獲得其中三名受試者的組織活檢標本,并對信使核糖核酸(mRNA)和棕色脂肪細胞的蛋白質標記物解偶聯蛋白1(UCP1)進行了基因表達水平的檢測;還對預留的樣品進行了形態學評估以及生化分析,結果符合棕色脂肪的胞內特征(分布著很多小脂滴、呈現多室的脂滴)。以上結果表明,在健康成人體內,仍存在大量的有代謝活性的棕色脂肪組織。
以前人們認為,棕色脂肪組織在新生兒時幫助維持人體正常體溫,之后這個組織隨著年齡的增長不斷退化,成年時則完全消失。現在認為,棕色脂肪組織仍存在于成人體內。已證明,在腫瘤自主性兒茶酚胺分泌(如,嗜鉻細胞瘤和副神經節瘤)的患者,棕色脂肪組織可以清楚地顯現出來。所以在診斷腫瘤和腫瘤轉移時,使用葡萄糖代謝示蹤劑F-FDG聯合PET和CT掃描,其結果很可能被這個因素(棕色脂肪組織被清楚地顯現出來)所干擾。鎖骨上組織的葡萄糖攝取量較高,是因為存在在該部位的棕色脂肪組織增加了葡萄糖攝取量。這個觀點已獲得證據支持。研究表明,通過CT圖像(由F-FDG作為示蹤劑)可以清楚顯現,脂肪組織對心得安(一種β受體阻滯劑, 用于治療心律不齊、心絞痛等)和環境溫度(PET掃描前)不同的敏感性;寒冷的冬季往往激活棕色脂肪組織產熱,而不是炎熱的夏季。但是就目前現有的知識,包括最近的研究調查結果,還沒有直接的數據證明,在健康成人受試者,冷環境誘導的F-FDG攝取增加的組織,具有棕色脂肪組織的病理生理特點,其mRNA和相關蛋白(以區分白色脂肪組織)的表達也明確上調。這一點很重要,這證明了健康的成人同樣存在棕色脂肪組織,這種組織在人類嬰兒期后仍可以發揮其正常的生理功能。這些數據很必要獲得證實。
方法
受試者
我們的研究共納入了5名健康受試者,并都簽署了書面知情同意書。研究方案由芬蘭醫院西南區的倫理委員會批準并進行了審查。受試者是通過在當地報紙刊登廣告的方法招募的。對自愿參加的所有人都進行了代謝狀況的檢查。受試者的納入條件是:糖耐量和心血管代謝狀態正常(通過心電圖和血壓進行評估),年齡范圍為20~50歲。
研究設計
研究分為冷暴露PET-CT研究和熱暴露PET-CT研究。共5名受試者,兩次PET-CT掃描都使用的是示蹤劑F-FDG。
冷暴露PET-CT研究設計:①受試者穿著單薄,室內溫度17℃~19℃,呆2小時;②冰水5℃~9℃,受試者的一只腳浸泡其中,冰水內外交替,各5分鐘;③進行PET-CT掃描。
熱暴露PET-CT研究設計:擇日進行,其流程和冷暴露PET-CT研究基本一樣,但不同的是,在掃描前沒有冰水刺激。
這兩部分最后的PET-CT掃描都是在受試者禁食過夜之后進行的(仰臥)。下一步進行組織活檢。活檢部位根據影像圖上顯現的被激活的褐色脂肪組織所在的部位來確定。組織活檢共包括5名受試者中的3名(簽署了脂肪組織活檢知情同意書),都在局麻條件下進行組織切片(包括棕色和白色脂肪組織),進行組織留樣,制成標本(見NEJM. org)。
PET掃描
按照圖爾庫PET中心的標準操作程序(以Hamacher等人先前使用的方法為基礎,經過修訂后的操作程序版本),采用F-FDG標記的正電子示蹤劑進行聯合顯像。
PET試驗開始時,通過導管經一側臂靜脈注入F-FDG;掃描期間獲得對側臂靜脈血液樣品。
活組織切片程序
受試者活組織切片是在局麻(利多卡因聯合腎上腺素)情況下進行的。
棕色脂肪組織切片:根據3名受試者(開放標簽)的PET-CT影像圖顯現的結果,整形外科醫生對冷環境誘導的葡萄糖攝取量增加的部位,分別進行組織切片,獲得受試者對應的組織樣品(簡稱為,標本1,標本2,標本3)。組織切片隨即被分為兩個部分,一部分用福爾馬林固定用于病理生理檢測;另一部分存放于液態氮中冷凍以備用。接著對mRNA和cDNA進行實時定量的PCR分析。最后利用備用的樣品對相關蛋白進行提取。
白色脂肪組織切片:和進行棕色脂肪組織切片同時進行,選擇同一個手術的相同切口下的皮下脂肪,獲得的組織切片采用同樣的操作程序檢測或留樣,以作為對照。
共軛焦顯微鏡觀察
我們進行了組織免疫化學的研究。把細胞培養在玻片上進行固定與染色,使用UCP1抗體,HRP(辣根過氧化物酶)標記抗體(經過化學方法將HRP標記在抗體IgG分子上而制成的HRP-抗體IgG結合物);之后再以共軛焦顯微鏡觀察。共軛焦顯微鏡所看到的影像利用的是其內建的掃描器,通過電腦運算將許多亮點組成線,很多線再組成單一平面影像的光切片,不同平面的切片組合在一起,就形成了三維空間的立體影像,再加入時間因子,就形成了動態的4D影像。
結果
受試者(5名)各自的影像圖比較,結果顯示一致,即,冷環境誘導(和熱環境誘導相比)F-FDG攝取增加,最顯著的部位是鎖骨上區(圖1A,1B和1C)。通過對影像圖的統計分析(圖1D),結果顯示,鎖骨上區的棕色脂肪組織比鄰近部位的白色脂肪組織的葡萄糖攝取量顯著增多,冷環境比熱環境誘導的葡萄糖攝取量,在棕色脂肪組織平均增加約15個單位(P=0.005),在白色脂肪組織增加4個單位(P=0.01)(圖1E)。
對應活組織切片標本No1、2、3的受試者No1、2、3的手術部位,分別是圖1A、1B和1C中用箭頭顯示的部位。采用定量PCR技術對多基因的mRNA表達水平的分析結果見圖2。對活組織切片標本的評估結果表明,作為棕色脂肪組織的標記基因UCP1,其在棕色脂肪組織的表達水平比其在白色脂肪組織的表達水平增加1000個單位還多。
UCP1具有解偶聯作用。電子傳遞鏈產生H+跨線粒體內膜的勢能。在偶聯狀態下,勢能驅使H+通過ATP 合成酶而重新回到線粒體基質中,同時將勢能轉化為ATP 的化學能。由于UCP1在線粒體內膜上有特殊的通道,由電子傳遞鏈泵出的氫離子直接通過這種通道流回線粒體內,使跨線粒體內膜的質子電化學梯度消失,ADP不能進行磷酸化形成ATP,物質氧化與ATP 生成脫偶聯,即質子滲漏,而成為產熱過程,能量以熱能形式釋放,減少了ATP合成。這是一種自主性產熱過程,目前認為這一過程對嚙齒類動物(如,動物冬眠,人類新生兒)維持正常體溫很重要。UCP1基因敲除小鼠的研究結果顯示,在急性冷暴露條件下,表現維持正常體溫的能力嚴重衰退,從而證明了UCP1對這一過程的有益作用。

圖1-1 PET-CT掃描影像

圖A、B、C分別顯示的是受試者1、2、3的頸胸部影像。PET-CT使用的是示蹤劑F-FDG。每一行左邊顯示的是經軸切片,中間是冠狀切片,右邊是矢狀切片。箭頭顯示的是葡萄糖攝取增加的鎖骨上部激活的棕色脂肪組織。影像圖左邊顯示了F-FDG攝取水平的顏色比對,紅色端表示最高水平。
圖2顯示了3名受試者棕色脂肪組織中的幾個基因表達水平。棕色脂肪細胞DIO2的mRNA表達明顯上調(和白色脂肪細胞相比)。這個結果很有意義,棕色脂肪細胞表達Dio2的目的,可能是為了使T3(三碘甲狀腺氨酸)能夠繼續保持自身(棕色脂肪組織)高水平的新陳代謝。此外,圖2還顯示,棕色脂肪組織中的PGC1α的mRNA表達也明顯上調。這并不令人驚訝,因為,冷環境誘導在很大程度上是首先通過激活PGC1α來使UCP1基因表達上調的。激活PGC1α是誘導產生UCP1的關鍵所在;在PGC1α敲除小鼠,cAMP誘導和冷誘導UCP1的活化水平顯著降低。PRDM16是調節棕色脂肪細胞形成的關鍵轉錄因子(圖2)。在成熟嚙齒動物的棕色脂肪組織,ADRB3(β3-腎上腺素受體)是β-腎上腺素受體三個亞型中最重要的。我們發現,如圖2所示,3名受試者的棕色脂肪組織中這種受體亞型表達也顯著上調(和白色脂肪組織進行比較)。
圖2顯示的mRNA表達數據,是根據PETCT影像圖確定的脂肪組織樣本進行的檢測結果,顯示了棕色脂肪組織預期的基因表達譜。為了明確冷誘導F-FDG攝取增加的部位的組織樣本表達UCP1,我們進行了Western blot(蛋白質印跡分析),結果顯示,3名受試者所對應的棕色脂肪組織樣本都檢測到UCP1蛋白,而對應的白色脂肪組織對照樣本中沒有檢測到任何UCP1,和預期的一樣(圖3A)。我們還檢測了線粒體標記物細胞色素C,結果發現,棕色脂肪組織比白色脂肪組織有更多更充足的細胞色素C(圖3B)。
對3名受試者的活檢樣品進行組織學分析,結果清楚地表明,鎖骨上部棕色脂肪組織細胞內含有多室的脂滴,而鄰近的白色脂肪組織細胞中卻沒有(圖3C)。UCP1免疫熒光分析結果顯示,棕色脂肪組織的UCP1水平很高,而白色脂肪組織幾乎沒有;這又一次說明基因表達和組織學特征間的關系(圖3D)。
共軛焦顯微鏡觀察結果顯示,在棕色脂肪組織,UCP1蛋白信號和線粒體標記物COI(細胞色素氧化酶亞基I)的信號重疊,從覆蓋面看,可以說兩者共區域化(圖3E)。在白色脂肪組織切片上沒有檢測到或觀察到UCP1蛋白(圖3E)。

圖1-2 冷熱暴露條件下脂肪組織消耗葡萄糖的量
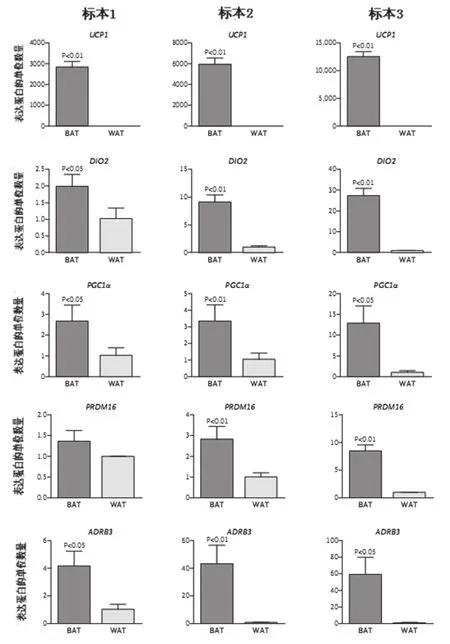
圖2 棕色和白色脂肪組織中的基因表達

圖3-1 棕色和白色脂肪組織的免疫組織學分析
討論
PET-CT研究和組織活體切片標本分析都表明,健康成人體內存在棕色脂肪組織。激活的棕色脂肪組織和白色脂肪組織相比,UCP1蛋白和細胞色素C的表達水平顯著增強。人類棕色脂肪組織細胞內的線粒體非常致密,相比之下,白色脂肪組織細胞內的線粒體稀疏。
從本實驗獲得的數據和之前對棕色脂肪組織代謝研究的調查結果,我們推測,冷環境誘導激活棕色脂肪組織,可能對人類的能量平衡方面很重要。例如,從我們實驗數據之一,PET-CT掃描得出的數據看,兩側鎖骨上部棕色脂肪組織的重量為63g,葡萄糖的吸收率為12.2μmol/100g/min,那么可以得出整個組織的吸收率為7.7μmol/ min。如果激活的持續時間為24小時,那么由棕色脂肪攝取的葡萄糖一共為11 mmol。考慮到游離脂肪酸可促進質子經棕色脂肪組織細胞的解偶聯蛋白返流至線粒體,這一發現可能是重要的。事實上,來源于所攝取的葡萄糖熱量,只占整體(激活的棕色脂肪組織所產生)熱量的10%左右。因此,如果該研究中的棕色脂肪組織得到充分激活的話,它所能消耗的能量相當于4.1kg脂肪組織一年所能消耗的能量總和。我們認為這是一個溫和的假設,因為棕色脂肪組織的活化程度,很可能沒有達到最大(我們估計是50%)。此外,嚙齒動物模型的研究表明,完全激活棕色脂肪組織時所產生的總熱量,來源于葡萄糖的“貢獻”估計是2%。在此實驗結果的基礎上,我們推測,激活棕色脂肪組織有可能對人類的能量消耗有重要作用。

圖3-2 棕色和白色脂肪組織的免疫熒光分析
總之,我們的研究表明,在冷環境背景下,鎖骨上部棕色脂肪組織的葡萄糖攝取率比白色脂肪組織增加15個單位。還發現該組織表達多個基因(UCP1、DIO2、PGC1α、PRDM16和ADRB3)的mRNA;表達UCP1蛋白和細胞色素C的水平也顯著上調(由免疫印跡分析評估);此外,該組織細胞內含有多室的脂滴。最后,還顯示了該組織所產生的UCP1蛋白的線粒體定位。我們認為,這些研究結果直接鑒定了人類棕色脂肪組織的功能。依據生化、分子和形態學標準,棕色脂肪組織仍存在于健康成人。因此,我們認為健康成人棕色脂肪組織值得進一步研究;鑒于目前肥胖流行,該組織將可能作為藥物的靶向目標。
10.3969/j.issn.1672-7851.2011.02.010
瑞典生物醫藥研究所臨床遺傳學醫學院
(莊稼英 編譯)
