食管癌癌變過程中端粒DNA長度與細胞DNA含量的變化及意義
2011-05-31 03:42:00王大虎張祥宏
中國老年學雜志 2011年10期
關鍵詞:檢測
王大虎 左 靜 張祥宏
(河北醫科大學基礎醫學研究所病理教研室,河北 石家莊 050017)
染色體末端存在一種維持染色體完整和穩定的特殊結構——端粒(telomere),能防止染色體DNA降解、末端融合缺失和非正常重組〔1〕。人端粒序列為(GGATTT)n,端粒長度(terminal repeat fragment,TRF)為5~15 kb,為染色體末端提供了一個保護性“帽”,對染色體的穩定十分重要〔2〕。每次細胞分裂,端粒會縮短55~200 bp,經歷多次分裂短至一定界限,細胞最終凋亡或死亡。近年來研究發現,端粒長度的改變參與腫瘤的發生和發展。但在食管癌癌變過程中是否存在端粒長度的改變及其與DNA含量的關系目前報道甚少。因此,本研究收集河北省磁縣食管癌高發區人群的食管脫落細胞,定量檢測食管上皮癌變過程中細胞端粒DNA長度改變和細胞DNA含量的變化,分析其與食管上皮癌變的關系,為闡述食管上皮的癌變機制提供實驗依據。
1 材料與方法
1.1 標本來源 所有標本均來自河北省磁縣食管癌高發區現場,采用食管拉網法采集食管上皮脫落細胞,將拉網器網球上的脫落細胞部分作細胞涂片,部分收集后70%乙醇固定用于流式細胞術檢測。隨機抽取100例,其中食管上皮正常組18例,輕度增生組16例,重度增生組35例,癌組31例。細胞學涂片經常規巴氏染色,由2名富有經驗的病理醫師進行細胞學診斷〔3〕。
1.2 主要試劑 端粒肽核酸(PNA)探針試劑盒購自DAKO公司(lot 00004334和00009155),碘化丙啶(Sigma公司)。
1.3 DNA染色 制備單細胞懸液,參照左連富的方法〔4〕,采用碘化丙啶一步插入性DNA熒光染色法,以10%雞紅細胞作內參染色。每份樣品中加入DNA染液(每1 000 ml中含碘化丙啶 50 mg/L,RNA 酶 10 mg/L,TritonX100 1.0%)1.0 ml,4℃冰箱中染色30 min,以500目篩網過濾,使樣品成為合格的單細胞懸液,即可上機檢測。
1.4 原位雜交 制備單細胞懸液,取2×106個細胞分為實驗組和背景對照組;實驗組加入300μl含FITC-(CCCTAA)3PNA探針雜交液,對照組加300μl空白雜交液,混勻后87℃避光孵育15 min變性;然后室溫下避光雜交過夜;洗滌后碘化丙啶進行DNA染色(4℃避光孵育2 h),400目銅網過濾后進行流式細胞儀測定〔5〕。
1.5 流式細胞儀檢測方法 采用美國BeckmanCoulter公司Epics2XLⅡ型流式細胞儀,激發光源為15 mW氬離子激光器,激發波長為 488 nm。檢測前以 flow-checkTMFluorpheres(10 μm)熒光微球(REF 6605359.Beckman Coulter,Inc.Fullerton,CA 92835.)作為標準樣品調整儀器CV值在2%以內。
1.6 數據分析 細胞增殖活性的分析:應用DNA細胞周期分析軟件,計算出DNA組方圖各時相分布的百分比,以增殖指數(PI)表示細胞的增殖活性,PI=(S+G2/M)/(G0/G1+S+G2/M)×100%。流式原位雜交分析:FL1通道檢測FITC熒光強度,FL3通道檢測碘化丙啶熒光強度,端粒熒光定量即Q-FISH值由減去背景后的G0/G1期二倍體細胞平均熒光強度決定。Q-FISH值=實驗樣品平均熒光強度-背景對照平均熒光強度。細胞DNA含量分析:依據二倍體參考細胞G0/1期細胞DNA含量定為2C值DNA指數(DNA index,DI)=1.0,基于標準CV值在5%,判定標準即:DNA二倍體=2C±2CV值,DNA異倍體≠2C±2CV。即:①DI=1.0±0.1(0.9~1.1)為二倍體②DI=1.0±0.15(0.85~1.15)為近二倍體③DI=2.0±2CV(1.9~2.1)為四倍體④DI>2.1為多倍體⑤其余DI值均為非整倍體。在計算DNA異倍體時,把近二倍體、四倍體、多倍體、非整倍體劃為異倍體的范疇。
1.7 統計學分析 計量資料用x±s表示,應用 SPSS13.0統計軟件包進行t檢驗、單因素方差分析和χ2檢驗,相關性分析采用Spearman相關分析。
2 結果
2.1 食管上皮癌變過程中細胞周期分布變化 正常組、輕度增生組、重度增生組及癌組 PI值分別為(11.51±4.08)、(14.08±5.00)、(19.04±5.39)和(29.46±5.50),隨著細胞學分級的升高,PI升高,癌組顯著高于重度增生組(P<0.01),重度增生組高于輕度增生組(P<0.05);輕度增生組與正常組相比雖有升高的趨勢,但無顯著差別(P>0.05)。相關性分析顯示,PI值與細胞學分級呈顯著正相關關系(r=0.79,P<0.01)。2.2 食管上皮癌變過程中細胞DNA含量的變化及其與細胞學分級的關系 癌組的DI值明顯高于重度增生組、輕度增生組和正常組(P<0.01);而正常組、輕度增生組和重度增生組的DI值雖呈上升趨勢,但無統計學意義(P>0.05);四組間異倍體率比較顯示,癌組異倍體率高于重度增生組(P<0.01)和輕度增生組,且重度增生組高于輕度增生組(P<0.05),但輕度增生組與正常組相比無明顯差別(P>0.05)。經Spearman等級相關分析顯示,DI值與細胞學分級呈正相關關系(r=0.60,P<0.01),提示隨著細胞學分級的升高,DI值增大,二倍體所占百分比明顯降低,異倍體百分比明顯升高。見表1,圖1,圖2。
2.3 食管上皮癌變過程中端粒DNA長度的變化及其與細胞學分級的關系 流式原位雜交顯示,正常組、輕度增生、重度增生和癌組中Q-FISH值分別為(50.83±8.86)、(49.51±3.16)、(36.96±8.02)和(27.81±6.59),癌組端粒長度明顯短于重度增生組和輕度增生組(P<0.01),重度增生組短于輕度增生組(P<0.01),但輕度增生組與正常組比較無顯著性差別(P>0.05)。經Spearman等級相關分析顯示,端粒長度與細胞學分級呈負相關(r=-0.79,P<0.01),隨著細胞學分級增高,端粒長度Q-FISH值逐漸降低。見圖1,圖3。
按照端粒長度的劃分標準〔6〕,即Q-FISH值<正常組端粒長度80%(<0.76)為縮短,>正常組織120%(>1.14)為延長。100例標本中輕度增生組9例,重度增生組22例和癌組26例端粒縮短,其端粒縮短的陽性率分別為56.25%(9/16)、62.86%(22/35)和83.87%(26/31),呈上升趨勢(P>0.05),但差異無統計學意義。見表2,圖1。

表1 流式細胞術檢測不同組別的DNA含量和異倍體變化
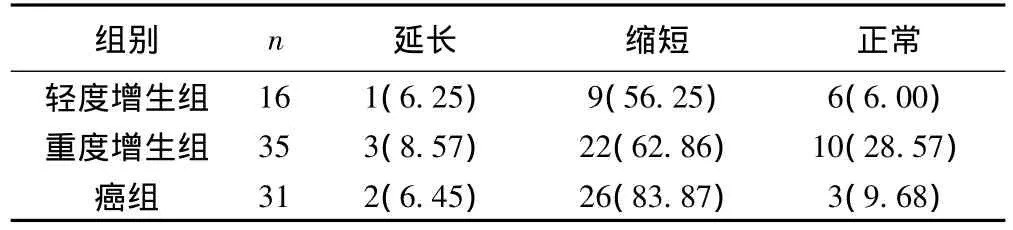
表2 食管上皮不同組別中端粒縮短情況〔n(%)〕

圖1 流式細胞術檢測端粒長度和DNA含量在各組中的變化

圖2 在正常食管上皮和食管癌細胞DNA倍體的不同

圖3 流式細胞術定量檢測端粒長度在各組中的變化
2.4 食管上皮癌變過程中細胞端粒DNA長度與細胞DNA含量的相關性分析 FCM檢測的100例食管上皮脫落細胞標本中,隨著細胞分級逐步升高,端粒長度Q-FISH值逐步下降的同時,DI值升高,DI值與端粒長度Q-FISH值呈顯著負相關關系(r= -0.46,P <0.01)。
3 討論
原癌基因的激活、抑癌基因的突變是腫瘤形成的重要分子基礎。正常細胞在致癌因素作用下轉變為惡性細胞并不意味著一定能形成腫瘤,哺乳動物體內有一套精細的調節機制控制著細胞的分裂次數,使細胞分裂達到一定程度后死亡,可見腫瘤細胞獲得無限增殖的特殊能力——永生化,在腫瘤的發生中起著至關重要的作用。
目前研究發現,端粒長度的改變參與了腫瘤的發生和發展。Maruyama等〔7〕檢測了不同病變期胃黏膜的端粒長度,發現從正常胃黏膜到腸化、再到腺瘤的發展過程中,端粒長度逐漸縮短。Meeker應用熒光原位雜交對114例浸潤性乳腺癌、29例原位癌和10例良性增生乳腺組織檢測證實,80.0%的浸潤性乳腺癌和78%的導管原位癌與自身正常對照相比,平均端粒長度有明顯縮短〔8〕。為了證實端粒與食管癌發生、發展的關系,本研究采用現場實驗和實驗室相結合的方法,應用熒光原位雜交流式定量法〔5〕(Q-Flow FISH)對食管癌高發區隨機抽取的100例食管上皮脫落細胞進行端粒長度的檢測。結果顯示,食管癌及癌前病變細胞的平均端粒長度均顯著短于正常食管上皮細胞,其平均端粒長度隨細胞增生的程度和癌變發展階段呈漸進性縮短,并且半數以上(56.25%輕度增生、62.86%重度增生)癌前病變端粒長度與正常組相比有顯著縮短,提示端粒縮短參與了食管上皮癌變的發生。同時本研究還發現,無論是食管癌細胞還是癌前病變細胞,端粒變化呈非均一性,既有縮短又有延長,8.57%重度增生和6.45%癌端粒長于正常平均長度,但是端粒總體變化趨勢與食管癌及癌前病變的輕重程度相一致。考慮可能源于幾個不同原因:①除端粒酶之外還有其他機制(如ALT)參與端粒穩定的調節;②在端粒酶或ALT起作用維持端粒長度之前,細胞增殖程度影響著總體端粒縮短水平;③細胞分裂過程中端粒斷裂-融合-橋聯(breakage-fusion-bridge,BFB)造成其長短不一〔9〕;④證據顯示未經修復的DNA氧化損傷也促進了端粒的丟失〔10〕。
在一些腫瘤中已觀察到,染色體末端的端端融合在腫瘤形成相關基因的不穩定性中起著重要作用,而這種融合可能起因于染色體末端長片段的丟失,如端粒的丟失〔11,12〕。基于這種觀點,端粒的縮短可能引起DNA的不穩定性,并在腫瘤的形成中起著重要作用。Ma〔13〕報道,在伴有DNA含量增高的腫瘤細胞中,存在染色體融合和(或)雙微染色體。為了證實食管上皮癌變過程中端粒長度的變化與DNA含量的關系,本研究應用流式細胞術分析不同食管黏膜上皮細胞內DNA含量。結果顯示,隨著食管上皮從正常、輕度增生、重度增生到癌的進展,DI值明顯增高,即DNA含量明顯增加,同時伴有異倍體數目的不斷增高,癌細胞組異倍體率達到80.65%;同時,隨著癌前病變的進展,PI值逐步增高,處于增殖期的細胞明顯增多。對31例癌的端粒長度和DNA指數的分析顯示,總體上,在異倍體中的端粒長度明顯短于二倍體,端粒長度和DNA指數呈負相關,也就是端粒越短,DNA指數越高。
本研究采用Q-Folw FISH對不同病變的食管上皮細胞進行端粒長度檢測,此方法簡單、快速,適合體外研究和進行大規模高發區現場患者的臨床研究,與傳統的Southern印跡測量的TRF值有良好的相關性。試驗標本采用的是食管拉網采集的脫落上皮細胞,相對減少了非上皮細胞成分的混雜,實驗數據更加精確。
綜上所述,本研究采用流式細胞術的方法對河北省磁縣食管癌高發區現場食管拉網獲得的食管上皮細胞進行了端粒長度和DNA含量等的檢測和分析,結果提示在食管上皮癌變過程中存在著端粒長度及DNA含量的改變,且在癌變早期就出現端粒長度的縮短。考慮獲得惡性表型的食管上皮細胞端粒、DNA長度不斷縮短,同時伴有染色體DNA活躍擴增,最終端粒維持在短而相對穩定的狀態下,導致了惡性細胞永生化,引起食管癌的發生。對食管上皮細胞進行端粒長度檢測將有助于監測食管癌的發生,同時也為抗腫瘤治療開辟了新方向。
1 Bellon M,Nica C.Regulation of telomerase and telomeres human tumor viruses take contro〔lJ〕.JNCI,2008;100(2):98-108.
2 Svenson U,Roos G.Telomere length as a biological marker in malignancy〔J〕.Biochim Biophys Acta,2009;1792:317-23.
3 沈 瓊.食管細胞學診斷〔M〕.北京:人民衛生出版社,1983:134-9.
4 左連富,林培中,齊鳳英,等.食管癌前細胞DNA、端粒酶含量及多基因表達產物的定量檢測〔J〕.生物化學與生物物理進展,2002;29(2):273-7.
5 黃 馨,石桂英,陳顯達,等.利用流式熒光原位雜交法測定細胞端粒長度〔J〕.中國比較醫學雜志,2010;20(7):67-71.
6 Hiyama E,Yokoyama T,Tatsumoto N,et al.Telomerase activity in gastric cancer〔J〕.Cancer Res,1995;55:3258-326.
7 MaruyamaY,Hanai H,Kaneko E.Telomere length and telomerase activity in intestinal metaplasia adenoma and well-differentiated adenocarcinoma of the stomach〔J〕.Nippon Rinsho,1998;56(5):1186-9.
8 Salhab M,Jiang WG,Newbold RF,et al.The expression of gene transcripts of telomere associated genes in human breast cancer:correlation with clinicopathological parameters and clinical outcome〔J〕.Breast Cancer Res Treat,2008;109(1):35-46.
9 Cesare AJ,Reddel RR.Telomere uncapping and alternative lengthening of telomeres〔J〕.Mech Ageing Dev,2008;129(1):99-108.
10 Tatsumoto N,Hiyama E,Murakami Y,et al.High telomerase activity is an independent prognostic indicator of poor outcome in colorectal cancer〔J〕.Clin Cancer Res,2000;6:2696-701.
11 de Lange T,Shelterin C.The protein complex that shapes and safeguards human telomeres〔J〕.Genes Dev,2005;19:2100-10.
12 Palm W,de Lange T.How shelterin protects mammalian telomeres〔J〕.Annu Rev Genet,2008;42:301-34.
13 Ma C,Martins S,Trask B,et al.Sister chromatid fusion initiates a amplification of the dihydrofolate reductase gene in Chinese hamster cells〔J〕.Genes Dev,1993;7:605-20.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
