寒冷刺激誘導大鼠棕色脂肪組織中的心臟干細胞增多
2011-07-31 03:08:20楊俊杰劉志強王海濱劉劍鋒陳韻岱
中國循環雜志 2011年5期
楊俊杰,劉志強,王海濱,劉劍鋒,陳韻岱
干細胞治療缺血性心臟病正引起基礎研究者和臨床工作者的極大興趣。脂肪來源干細胞因其取材方便、易于擴增、多能性好、免疫原性弱等優勢,逐漸成為研究熱點[1,2]。作為脂肪組織的一種重要類型,棕色脂肪在人體內的活性和作用正受到極大關注。棕色脂肪組織有別于常見的白色脂肪組織,可特異性表達解偶聯蛋白(UCP-1)通過解除氧化呼吸鏈中氫離子梯度而抑制能量物質三磷酸腺苷的產生,并最終將生物反應以熱能的形式釋放出來[3]。以往認為棕色脂肪只存在于小型哺乳動物和嬰幼兒體內,但進一步研究發現成人體內仍存在一定數量但活性較低的棕色脂肪[4]。新近的研究還發現乳鼠的棕色脂肪組織內含有一定數量(0.2% ~4%)的心臟前體細胞(又稱心臟干細胞)[5],具有體外自發分化和誘導分化能力[6],較其他干細胞更具運用前景[7]。本研究利用寒冷低溫條件飼養大鼠,檢測其肩胛部棕色脂肪組織活性變化;采用已經報道過細胞提取的方法[8],研究寒冷刺激對于肩胛部棕色脂肪中心臟干細胞含量的影響。
1 材料與方法
實驗分組:2011-03-01從軍事醫學科學院實驗動物中心購買SD大鼠12只(80~120g)。隨機分為2組,每組6只。實驗組在寒冷環境下(4℃)飼養,對照組在常溫條件(24℃)飼養,均飼養28天。兩組飲水量和喂食等處理均相同。
棕色脂肪來源細胞的提取與培養:斷頸處死各組大鼠,隨后取盡肩胛間區的棕色脂肪組織稱重,一部分進行棕色脂肪組織的驗證,另一部分運用酶消化法分離培養獲得棕色脂肪來源細胞。從SD大鼠肩胛部分離脂肪組織1 g進行細胞提取。分離的脂肪組織以無菌液大量沖洗。隨后,用含0.1% Ⅰ型膠原酶、0.1%中性金屬蛋白水解酶和0.05%胰酶的無血清培養基在37℃下消化45分鐘,后使用含10%的無血清培養基終止消化,并通過75 μm的濾網過濾、紅細胞裂解液去除紅細胞。最終將細胞進行離心和重懸,并接種至含10%胎牛血清的培養基中培養,1天后換液棄除未貼壁細胞,每隔兩天觀察并換液。
棕色脂肪組織形態及活性:經冰凍切片包埋劑固定的大鼠肩胛間區的棕色脂肪組織標本經切片后,使用蘇木素伊紅染色法染色并觀察棕色脂肪組織形態。組織蛋白經處理后使用western blot方法檢測UCP-1表達情況。
細胞表面標記物的檢測:提取得到的棕色脂肪來源細胞經過紅細胞裂解液處理后,使用表面標記物CD29、CD34、CD45、CD90 和 CD133 抗體,按照廠家說明加入以上抗體,使用流式細胞儀鑒定。
自發分化心肌樣細胞的檢測:以1×105/cm2、5×104/cm2、1×104/cm2、5×103/cm2、1×103/cm2的細胞密度接種。培養4周后進行免疫熒光檢測。免疫熒光檢測:取出預先置入的細胞爬片固定。經0.1%TrionX-100打孔,分別在肌鈣蛋白-T(cTnT)抗體(1∶200)和α-橫紋肌輔肌動蛋白(α-SCA)抗體(1∶200)中4℃孵育過夜。隨后以二抗標記,并染核,在熒光倒置顯微鏡下觀察物鏡40×視野下綠色熒光范圍中的細胞核數量與總核數量的百分比。
核糖核酸(RNA)提取和逆轉錄多聚酶鏈反應分析:分化后的細胞培養皿中,在倒置位差顯微鏡下選取狹長成簇的細胞進行消化和提取。Trizol法提取各自總RNA,以大鼠心肌組織作為陽性對照。按逆轉錄試劑盒(TIANGEN)方法步驟逆轉錄并進行多聚酶鏈反應擴增,最后進行凝膠成像。在多聚酶鏈反應擴增中使用的引物包括:cTnI(5’-CTCGGAGTATCAGGAAGAGCACA;3’-TAAACTTGCCACGCAGGTCATAG,216 bp),GATA-4(5’-CTGTCATCTCACTATGGGCA;3’-CCAAGTCCGAGCAGGAATT,257bp),Nkx2.5(5’-CAGTGGAGCTGGACAAAGCC;3’-TAGCGACGGTTCTGGAACCA,216 bp),MEF-2C(5’-AGCAAGAATACGATGCCATC;3’-GAAGGGGTGGTGGTACGGTC,347 bp),β-Actin(5’-AGAGGGAAATCGTGCGTGAC;3’-AGGAGCCAGGCAGTAATC,353 bp)。
反應條件為:各反應體系先94℃預變性3 min,接著35個循環擴增(94℃變性30 s,55℃退火30 s,72℃延伸1 min),最后延伸5 min至22℃。
統計學方法:數據以均數±標準差(x±s)表示,應用SPSS 13統計軟件進行統計分析,組間比較采用t檢驗或方差分析,以P<0.05為差異有統計學意義。
2 結果
2.1 寒冷刺激后的棕色脂肪形態與活性表達
寒冷刺激后,肩胛部棕色脂肪組織的形態特征較對照組發生相應變化,具體表現為脂肪細胞線粒體彌漫增多,脂肪滴變小,脂肪組織毛細血管基質增生。進一步檢測發現實驗組棕色脂肪組織中特異性功能蛋白UCP-1的表達量明顯增加。(圖1)
2.2 提取的棕色脂肪來源細胞中的表面標志物表達
細胞流式檢測發現,實驗組表面標記物CD29、CD133和CD29/CD133表達分別為(22.93±1.88)%、(15.12±1.47)% 和(9.13±1.23)%,均明顯高于對照組(16.31±1.52)%、(7.83±0.90)%和(4.22±0.62)%,差異均有統計學意義(P<0.01)。而間充質細胞特異性表面抗原以及造血干細胞表面特異性抗原的表達,在兩組中差異均未見統計學意義(P>0.05)。
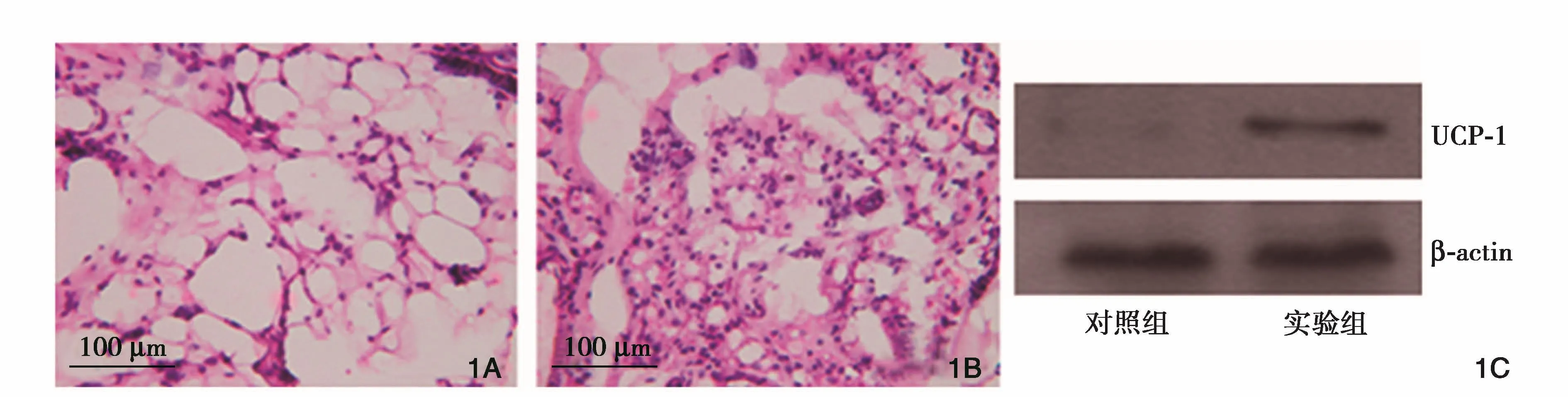
圖1 棕色脂肪形態與活性表達。1A:常溫飼養后提取的棕色脂肪組織形態(HE染色)。1B:低溫飼養后提取的棕色脂肪組織形態(HE染色)。1C:特異性蛋白UCP-1在兩組棕色脂肪組織中的表達 HE:蘇木素伊紅 UCP-1:解偶聯蛋白-1 β-actin:β-肌動蛋白
2.3 自發分化后的心肌樣標志物表達
定期顯微鏡下觀察,棕色脂肪來源細胞經體外培養逐漸出現心肌細胞的形態學和功能變化:細胞在1周時多數細胞貼壁鋪展生長,細胞分化數量較少,但有一定數量圓狀成簇樣細胞存在;2~4周后,狹長的細胞明顯增多,并表現為自發搏動的狀態。進一步免疫熒光檢測發現,該狹長型、具有自發搏動能力的細胞簇,可表達心肌樣特異性標志物cTnT和α-SCA。(圖2)
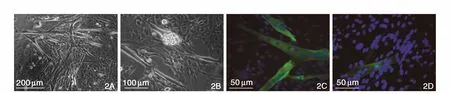
圖2 自發分化后的心肌樣標志物表達。2A:光鏡下細胞形態(×10)。2B:光鏡下細胞形態(×20)。2C:表達肌鈣蛋白-T。2D:表達α-橫紋肌輔肌動蛋白
不同接種密度下棕色脂肪來源細胞自發分化cTnT陽性細胞的比例:實驗組的自發分化cTnT陽性細胞比例顯著高于對照組,差異均有顯著統計學意義(P<0.01)。實驗組棕色脂肪來源細胞自發分化cTnT陽性細胞比例最高為(27.3±2.8)%,而對照組最高只有(4.1±0.7)%。(表1)

表1 不同接種密度下兩組棕色脂肪來源細胞自發分化肌鈣蛋白-T陽性細胞的比例(x±s)
2.4 心臟特異因子的表達
逆轉錄多聚酶鏈反應結果顯示,心肌細胞特異因子(肌鈣蛋白I、肌細胞增強因子-2C)和轉錄因子(轉錄因子GATA結合蛋白-4、同源框蛋白Nkx2.5轉錄因子)能夠在棕色脂肪來源細胞自發分化后的類心肌細胞中檢測出,但是較正常心肌組織表達量相比仍較為微弱。圖3

圖3 棕色脂肪來源細胞自發分化后的心臟特異因子和轉錄因子逆轉錄多聚酶鏈反應分析。1:棕色脂肪來源的類心肌細胞;2:大鼠心肌細胞(陽性對照);3:陰性對照。n=6。cTnI:肌鈣蛋白-I;MEF-2C:肌細胞增強因子-2C;GATA-4:轉錄因子GATA結合蛋白-4;Nkx2.5:同源框蛋白Nkx2.5轉錄因子;β-actin:β-肌動蛋白
3 討論
發現寒冷刺激后大鼠肩胛部棕色脂肪來源細胞的體外心肌自發分化現象顯著增加。同時,其肩胛部棕色脂肪組織形態結構發生相應的顯著變化,特異性蛋白明顯過表達,提示寒冷刺激下棕色脂肪組織活性顯著增強可能與上述現象有關。
相關文獻指出,棕色脂肪來源細胞中的CD29陽性和CD133陽性的細胞,具有高度自發分化為心肌分化的能力[9]。本實驗發現,寒冷刺激下棕色脂肪組織來源細胞表面標志物中CD29和CD133所占比例明顯升高,而且CD133表達陽性的細胞中有更多的表達CD29。雖然我們沒有做其它心臟干細胞特異抗原的檢測,但是在后續的體外培養中發現,經過4周無分化誘導培養基培養,寒冷刺激組中的心肌分化效率明顯增高。運用逆轉錄多聚酶鏈反應也鑒定了狹長細胞中的心肌特異性基因表達情況。同時細胞接種密度也影響著心肌自發分化的效率:通過設立不同接種細胞數目的分組,發現寒冷刺激組比常溫飼養組中的心肌特異性標記肌鈣蛋白表達效率明顯增高,這些都說明寒冷刺激組的棕色脂肪來源細胞體外心肌自發分化現象更為明顯,因而可以推測該組中的棕色脂肪中含有更多的心臟干細胞。盡管如此,完整的心肌細胞還需要成熟的膜電位和縫隙連接證實,這也是本實驗的不足之處。
從組織起源角度,脂肪組織和肌纖維組織都來源于胚胎中胚層。研究發現,白色脂肪和棕色脂肪的起源不盡相同,相比之下,棕色脂肪細胞與肌細胞更傾向于具有相同的前體干細胞來源。因此在棕色脂肪組織中發現肌細胞的前體細胞,甚至心臟干細胞及其自發分化的現象,確實具有一定的理論和前期研究基礎。雖然也有報道指出,白色脂肪組織中提取的細胞也具有體外分化心肌的能力,但效率非常低[10]。
在其他混雜因素被嚴格控制的前提下,以上的實驗結果說明寒冷刺激下,棕色脂肪活性與其中心臟干細胞含量有一定聯系。近年來,多項人體影像學研究顯示,成人體內仍然存在棕色脂肪,只是在寒冷、饑餓等狀態下,棕色脂肪組織的功能進一步活化以至可以被PET等技術檢測[11]。人體心臟內甚至也含有存在活性的棕色脂肪組織,在寒冷誘導下同樣可以被檢測到[12],更說明心臟修復與棕色脂肪組織之間存在一定關聯。我們有理由推測,棕色脂肪組織中的心臟干細胞,極有可能參與心臟的修復,并在寒冷等因素影響下,心臟干細胞的動員或增殖能力加強,使得體內心臟干細胞活性或絕對數量相應增加,可以更有效的修復受損心肌,改善心臟功能,降低心臟病預后。本研究對于拓展心臟干細胞的理念以及深入研究心臟干細胞治療都具有重大意義。
另一方面,棕色脂肪在成人體內確實較少,但是當棕色脂肪的轉化和擴增技術不再成為難題的時候,棕色脂肪組織或許可以在心臟病干細胞治療領域發揮巨大作用。如何進一步純化并分選棕色脂肪組織中的心臟干細胞,是下一步研究需要弄清的問題。同時,棕色脂肪在寒冷中的活性變化由諸多因素參與介導[13,14],心臟干細胞含量增多的具體機制與哪些因素有關還不得而知[15,16],也將成為進一步的研究方向。
[1]張衛澤,陳躍武,陳永清,等.5-氮胞苷體外誘導成人脂肪間充質干細胞分化為心肌樣細胞的初步研究.中國循環雜志,2006,3:228-231.
[2]Palpant NJ,Metzger JM.Aesthetic cardiology:adipose-derived stem cells for myocardial repair.Curr Stem Cell Res Ther,2010,5:145-152.
[3]Richard D,Picard F.Brown fat biology and thermogenesis.Front Biosci,2011,16:1233-1260.
[4]Biljana B,Aleksandra K,Vesna P,et al.Adaptive changes in interscapular brown adipose tissue during reacclimation after cold:The role of redox regulation.J Therm Biol,2007,32:261-269.
[5]Yamada Y,Wang XD,Yokoyama S,et al.Cardiac progenitor cells in brown adipose tissue repaired damaged myocardium.Biochem Biophys Res Commun,2006,342:662-670.
[6]Yamada Y,Yokoyama S,Fukuda N,et al.A novel approach for myocardial regeneration with educated cord blood cells cocultured with cells from brown adipose tissue.Biochem Biophys Res Commun,2007,353:182-188.
[7]Cypess AM,Lehman S,Williams G,et al.Identification and importance of brown adipose tissue in adult humans.N Engl J Med,2009,360:1509-1517.
[8]Liu Z,Wang H,Zhang Y,et al.Efficient isolation of cardiac stem cells from brown adipose.J Biomed Biotechnol,2010,2010:104-296.
[9]Yamada Y,Yokoyama S,Wang XD,et al.Cardiac stem cells in brown adipose tissue express CD133 and induce bone marrow nonhematopoietic cells to differentiate into cardiomyocytes.Stem Cells,2007,25:1326-1333.
[10]Jumabay M,Zhang R,Yao Y,et al.Spontaneously beating cardiomyocytes derived from white mature adipocytes.Cardiovasc Res,2010,85:17-27.
[11]Nedergaard J,Bengtsson T,Cannon B.Unexpected evidence for active brown adipose tissue in adult humans.Am J Physiol Endocrinol Metab,2007,293:E444-452.
[12]Virtanen KA,Lidell ME,Orava J,et al.Functional brown adipose tissue in healthy adults.N Engl J Med,2009,360:1518-1525.
[13]Petrovic V,Buzadzic B,Korac A,et al.NO modulates the molecular basis of rat interscapular brown adipose tissue thermogenesis.Comp Biochem Physiol C Toxicol Pharmacol,2010,152:147-159.
[14]Seebacher F,Glanville EJ.Low levels of physical activity increase metabolic responsiveness to cold in a rat(Rattus fuscipes).PLoS One,2010,5:e13022.
[15]王蕾,張世杰,王海鵬,等.阿托伐他汀對兔舒張性心力衰竭模型心室重構的影響.中國循環雜志,2010,25:223-226.
[16]張愛東,郭軍,李自成,等.胰島素樣生長因子-1預處理對大鼠骨髓間充質干細胞移植效率及心肌修復的影響.中國循環雜志,2011,26:57-60.
