應用基因芯片方法檢測結核分枝桿菌利福平和異煙肼的耐藥性
2011-09-20 10:02:40張俊仙吳雪瓊陽幼榮梁艷
中國防癆雜志 2011年10期
關鍵詞:檢測
張俊仙 吳雪瓊 陽幼榮 梁艷
眾所周知,耐多藥結核病已經嚴重影響了全球結核病的控制和治療。耐多藥結核病是指結核患者至少對利福平和異煙肼2種最主要的一線抗結核藥物耐受。然而,由于結核分枝桿菌(Mycobacteriu m tuberculosis,Mtb)是慢生長分枝桿菌,常規的藥敏試驗需4~8周的時間,因此結核患者通常不能得到及時、有效的治療,導致耐多藥結核患者結核菌的增殖和傳播。因此,需要建立一種操作簡便、快速的藥物敏感性試驗方法,這對于控制耐多藥結核病的傳播具有重要的意義。
近年來出現很多耐藥性檢測的新方法,如PCR-單鏈構象多態性(PCR-Single Strand Conformation Polymorphism,PCR-SSCP)、PCR-限制性片段長度多態性(PCR-restrition fragment length polymophism,PCR-RFLP)、DNA 測序法、分子燈塔法、噬菌體生物擴增法等,都因為各種各樣的問題難以在臨床上廣泛推廣。筆者應用PCR-基因芯片和基因測序方法對30株利福平和異煙肼敏感株和50株利福平和異煙肼耐藥株進行檢測,以期為耐多藥結核病的檢測提供簡單、快速、有效的方法。
材料與方法
一、菌株來源
80株Mtb臨床分離株由本室保存,已按全國結核病細菌學檢驗標準化規程進行菌種鑒定證實為Mtb,并根據傳統藥敏試驗結果選擇30株利福平和異煙肼敏感株、50株耐利福平和異煙肼分離株。Mtb標準株H37Rv來自中國藥品生物制品檢定所。
二、試劑和儀器
Mtb耐藥檢測試劑盒(芯片法)購自北京百奧瑞生物技術有限公司。PCR擴增儀和 LuxScan 10K-B微陣列芯片掃描儀等均為博奧生物有限公司提供。
三、細菌DNA的制備
采用本室自制的標本前處理試劑盒提取DNA,置-20℃保存備用[1]。
四、PCR擴增
按試劑盒說明書進行。每管PCR反應體系總體積為20 μ l,其中 Mtb分離株 DNA 模板為2 μ l,PCR擴增試劑1、PCR擴增試劑2和PCR擴增試劑3各 18 μ l。PCR產物1為對照產物,與兩種微陣列都進行雜交。PCR產物2為rpoB基因的擴增產物,與利福平微陣列相對應;PCR產物3為katG基因及inhA基因啟動子的擴增產物,與異煙肼微陣列相對應。PCR擴增程序為:37℃600 s,94℃600 s;94℃30 s,60℃30 s,72℃40 s,35個循環;然后94℃30 s,72℃60 s,10個循環;最后置 72℃420 s。
五、基因芯片
檢測探針和對照探針各重復5個點,每行1~5和6~10分別為同一個探針,利福平相關rpoB基因的探針排布示意見表1。異煙肼相關katG基因和inhA基因啟動子的探針排布示意見表2。
六、芯片雜交和結果判讀
根據試劑盒說明書進行操作。簡而言之,每管含雜交緩沖液9 μ l,分別加入同一樣品的PCR產物1和 PCR產物2各 3 μ l,或者加入 PCR產物 1和PCR產物3各3 μ l。置于PCR擴增儀中于95℃變性5 min,然后冰浴 3 min;將13.5μ l雜交反應混合物經蓋片的加樣孔加入,迅速蓋上雜交盒并密封;放入50℃預熱的恒溫水浴鍋中120 min;水平取出雜交盒,拆開取出芯片,洗滌后離心甩干。置LuxScan 10K-B微陣列芯片掃描儀中讀取信號、判讀結果。

表1 利福平相關rpoB基因的探針排布
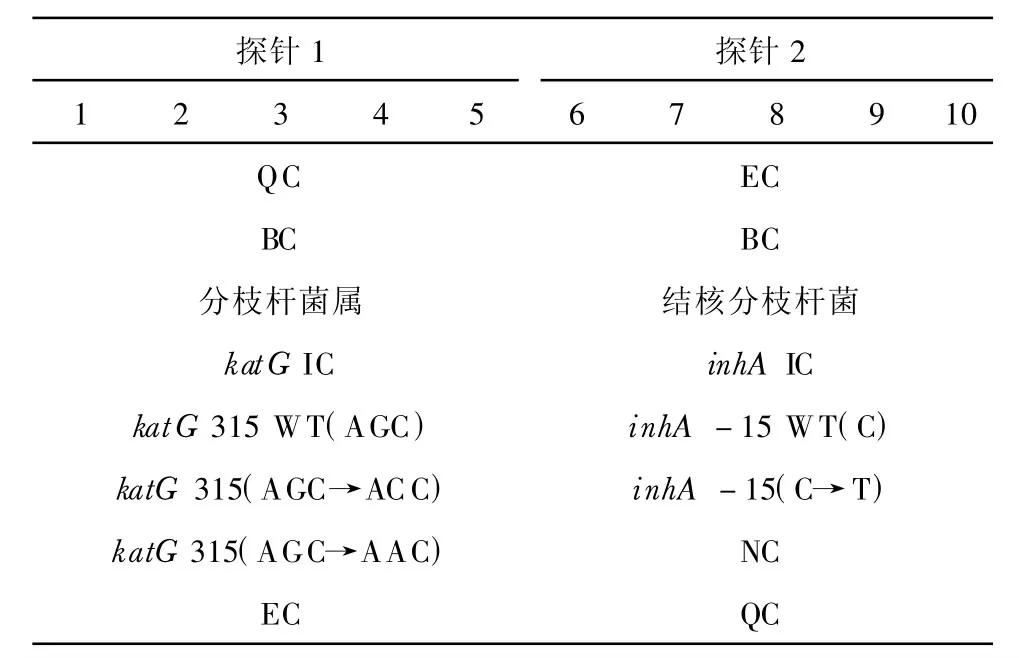
表2 異煙肼相關katG基因和inhA基因啟動子的探針排布
七、PCR-直接測序法
根據Mtb序列,用軟件Oligo 6.0設計利福平上下游測序引物rpoB3、rpoB4,可擴增rpoB基因產生240 bp片段,其中包括507~533位密碼子之間81 bp熱點區間的序列。設計異煙肼上下游測序引物 katG 1、katG2和 inhA1、inhA2,可分別擴增katG基因和inhA基因產生273 bp和255 bp片段,分別包括常見的katG基因315位密碼子和inhA基因啟動子-15位突變。擴增產物經2%瓊脂糖電泳,均為陽性,說明沒有基因缺失。擴增產物送上海生工生物工程公司進行基因測序。
結 果
一、芯片雜交結果
PCR-芯片雜交結果見表3。30株Mtb利福平敏感株rpoB基因、異煙肼敏感株katG基因和inhA基因芯片雜交結果與Mtb標準株結果一致,均為Mtb野生型。
50株Mtb耐利福平分離株中,7株芯片雜交為野生型;42株有單位點突變,1株有雙位點突變。以80株Mtb臨床分離株藥敏檢測結果為判斷標準,芯片檢測的靈敏度、特異度、陽性預測值、陰性預測值、準確率分別是 86.00%(43/50)、100.00%(30/30)、100.00%(43/43)、81.08%(30/37)、91.25%(73/80)。43株耐藥突變株中,28株(65.12%)為 rpoB基因 531位突變,其中 26株TCG→T TG突變,2株 TCG→TGG突變;6株(13.95%)為526位突變,其中4株CAC→TAC突變,1株CAC→CGC突變,1株CAC→GAC突變;4株(9.30%)516位突變,2株GAC→GTC突變,2株GAC→TAC突變;3株(6.97%)511位突變,均為CTG→CCG突變;1株(2.33%)513位CAA→AAA突變;1株(2.33%)雙位點突變為 511位CTG→CCG和516位GAC→GGC突變。部分芯片雜交結果見圖1。

表3 50株利福平和異煙肼耐藥株PCR-基因芯片檢測與基因測序結果

圖1 Mtb利福平rpoB基因部分芯片雜交圖
50株Mtb異煙肼耐藥株中,14株芯片雜交均為野生型,36株有katG和/或inhA突變,芯片檢測的靈敏度、特異度、陽性預測值、陰性預測值、準確率分別.72.00%(36/50)、100.00%(30/30)、100.00%(36/36)、68.18%(30/44)、82.50%(66/80)。31株芯片檢測有katG突變的分離株中,30株315位AGC→ACC突變和1株315位AGC→AAC突變;7株芯片檢測有inhA突變的分離株均為-15位C→T突變;其中2株既有katG突變又有inhA突變。部分芯片雜交結果見圖2。
二、PCR-直接測序結果
結果見表3。30株Mtb利福平和異煙肼敏感株DNA測序結果與基因芯片檢測結果一致,均為Mtb野生型。

圖2.tb異煙肼katG基因和inhA啟動子芯片雜交圖
50株Mtb耐利福平分離株中,44株基因測序結果與PCR-基因芯片檢測結果一致,其中3株均為rpoB野生型,41株均為突變型。6株基因測序結果與PCR-基因芯片檢測結果不一致,其中4株為單位點突變,芯片上無相應的探針(分別是531位TCG→TAG、515位 ATG→ATA、513位 CAA→CAT和522位TCG→CAG);2株測序結果為雙位點突變,其中各有1個突變位點芯片上無相應的探針,分別是516位GAC→AAC、526位CAC→CAG 突變。
50株Mtb耐異煙肼分離株中,47株katG基因測序結果與 PCR-基因芯片檢測結果一致,其中16株均為野生型,31株均為315位密碼子突變;3株katG基因測序結果與 PCR-基因芯片不一致,芯片上無相應的探針,分別為299位GGC→GTC突變、322位ACG→GCG突變和 291位 GCT→GAT突變。37株inhA基因測序結果與PCR-基因芯片檢測結果一致,其中30株均為野生型,7株均為-15位突變型;13株inhA基因測序結果與PCR-基因芯片檢測結果不一致,芯片上無相應的探針,其中10株為68位G→A突變,3株分別為-8位T→C、-21位G→C和-17位G→T突變。
討 論
目前的研究表明,大約95%以上的Mtb耐利福平分離株在RNA聚合酶β亞單位編碼基因rpoB的507-533位密碼子的81bp熱點區域有突變[2],90%的點突變發生在該區域11個氨基酸密碼子之一的部位,其中最常見的突變位點是531位、526位和516位,并且531>526>516[3-5]。而Mtb耐異煙肼主要與katG基因和inhA啟動子突變有關,katG基因最常見的突變是315位密碼子AGC→ACC的突變,該突變引起過氧化氫酶活性降低,導致異煙肼耐藥[6]。InhA基因編碼脂肪酸轉運蛋白(NADH依賴的enoyl-ACP還原酶),是分枝菌酸細胞壁合成的必需成分,活化的異煙肼結合到NADH上抑制了依賴NADH酶的活性,抑制了分枝菌酸的合成而導致了細胞的死亡。inhA基因的突變修改了該酶,使它失去了和NADH的親和力,導致異煙肼耐藥[7-8]。因此,這些區域是利福平和異煙肼耐藥株分子檢測的主要靶位[9-11]。
基因芯片技術是目前分子診斷的先進方法。國內外很多專家分別報道用基因芯片檢測耐利福平Mtb標本,其檢測結果與測序結果大多數相符[12-13]。因此,應用基因芯片快速檢出rpoB基因、katG基因和inhA基因突變可以作為耐多藥結核病的診斷指標。
本研究應用已商品化的基因芯片分析Mtb rpoB基因的6個野生型和13個突變型位點,分析katG基因315位密碼子的1個野生型和2個突變型及inhA基因啟動子區-15位的野生型和突變型,因此,可檢出耐多藥結核病常見的基因突變位點。
本實驗中30株Mtb利福平和異煙肼敏感株中,DNA芯片雜交結果也均為野生型,特異度為100.00%。50株耐利福平臨床分離株中43株可檢測到rpoB基因突變,靈敏度為86.00%,與郭乃洲等[14]報道的一致。這是因為該基因陣列探針只包括最常見的13種突變,基因測序結果證明4株突變株芯片上無相應的突變探針,因此,基因芯片檢測的靈敏度低于基因測序。此外,3株在分析的部位未發現基因突變,有可能在本檢測范圍外的其他位點存在突變,或者是存在其他耐藥機制,或者傳統藥敏實驗誤差所致。本試驗中,檢測到的最常見的突變位點也是531位、526位和516位密碼子,與報道一致。
50株Mtb耐異煙肼分離株中,36株有katG和(或)inhA突變,靈敏度為 72.00%,其中katG 基因突變率為62%,與袁瑛等[15]、菅記涌等[16]報道的一致;inhA-15位基因突變率僅為 14%,與Morris等[17]報道的一致。由于芯片只能檢測到常見的野生型和突變型,因此,katG基因測序顯示的3株少見的突變基因型芯片未能檢出。此外,16株在分析的部位未發現基因突變,筆者將進一步分析,查找原因。
在耐異煙肼分離株中,通過PCR-基因芯片只檢出7株inhA-15位C→T突變,但基因測序發現10株芯片檢測敏感的菌株存在inhA 68位G→A突變,該突變為首次報道,它在異煙肼敏感菌株中不存在,但在測序證實的20株inhA基因突變株中,該突變基因型占了50%,高于inhA-15位突變,該突變是否與異煙肼耐受相關尚需進一步研究證實。
總之,用PCR-基因芯片可快速、特異地檢測出大多數Mtb耐多藥分離株,用于臨床耐藥性檢測,可指導臨床治療。
[1] 張敦熔.現代結核病學.北京:人民軍醫出版社,2000:96-98.
[2] Musser JM.Antimicrobial agent resistance in mycobacteria:molecular genetic insights(Review). Clin Microbiol Rev,1995,8(4):496-514.
[3] Nachamkin I,Kang C,Weinstein MP.Detection of resistance to isoniazid,rifampin,and streptomycin in clinical isolates of Mycobacterium tuberculosis by molecular methods.Clin Infect Dis,1997,24(5):894-900.
[4] Jou R,Chen HY,Chiang CY,et al.Genetic diversity of multidrug-resistant Mycobacterium tuberculosis isolates and identification of 11 novel rpoB alleles in Taiwan.J Clin Microbiol,2005,43(3):1390-1394.
[5] Bolotin S,Alexander DC,Chedore P,et al.M olecular characterization of drug-resistant Mycobacterium tuberculosis isolates from Ontario,Canada.J Antimicrob Chemother,2009,64(2):263-266.
[6].usser JM,Kapur V,Williams DL,et al.Characterization of the catalase-peroxidase gene(katG)and inhA locus in isoniazid-resistant and-susceptible strains of Mycobacterium tuberculosis by automated DNA sequencing:restricted array of mutations associated with drug resistance.J Infect Dis,1996,173(1):196-202.
[7] Luckner SR,Liu N,am Ende CW,et al.A slow,tight binding inhibitor of InhA,the enoyl-acyl carrier protein reductase from Mycobacterium tuberculosis.J Biol Chem,2010,285(19):14330-14337.
[8] Dessen A,Quémard A,Blanchard JS,et al.Crystal structure and function of the isoniazid target of Mycobacterium tuberculosis.Science,1995,267(5204):1638-1641.
[9] Drobniewski FA,Watterson SA,Wilson SM,et al.A clinical,microbiological and economic analysis of a national service for the rapid molecular diagnosis of tuberculosis and rifampicin resistance in Mycobacterium tuberculosis.J Med Microbiol,2000,49(3):271-278.
[10] Chaoui I,Sabouni R,Kourout M,et al.Analysis of isoniazid,streptomycin and ethambutol resistance in Mycobacterium tuberculosis isolates from Morocco.J Infect Dev Ctries,2009,3(4):278-284.
[11] Banerjee A,Dubnau E,Quemard A,et al.Inh A,a gene encoding a target for isoniazid and ethionamide in Mycobacterium tuberculosis.Science,1994,263(5144):227-230.
[12] 梁莉,樂軍,李瑤,等.利用基因芯片技術檢測結核分支桿菌耐利福平分離株rpoB基因突變的研究.中華醫院感染學雜志,2005,15(8):841-844.
[13] Aragón LM,Navarro F,Heiser V,et al.Rapid detection of specific gene mutations associated with isoniazid or rifampicin resistance in Mycobacterium tuberculosis clinical isolates using non-fluorescent low-density DNA microarrays.J Antimicrob Chemother,2006,57(5):825-831.
[14] 郭乃洲,王萬相,嚴愛華,等.顯色法芯片檢測結核分枝桿菌利福平和異煙肼耐藥基因.中華微生物學和免疫學雜志,2006,26(6):567-570.
[15] 袁瑛,顧德林,顧紅艷.耐多藥結核分枝桿菌 KatG及rpoB基因變異的研究.臨床薈萃,2006,21(12):863-864.
[16] 菅記涌,王嫩寒,易俊莉,等.M TBDR plus方法快速檢測北京地區臨床結核分枝桿菌分離株利福平和異煙肼耐藥性效果評價.中國防癆雜志,2010,32(10):611-614.
[17] Morris S,Bai GH,Suffys P,et al.M olecular mechanisms of multiple drug resistance in clinical isolates of Mycobacterium tuberculosis.J Infect Dis,1995,171(4):954-960.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
