雜草對ACCase抑制劑的抗性
2011-11-20 08:17:21張朝賢黃紅娟魏守輝
雜草學報 2011年3期
郭 峰, 張朝賢, 黃紅娟, 魏守輝, 張 猛, 房 峰
(中國農業科學院植物保護研究所/中國農業科學院雜草害鼠生物學與治理重點開放實驗室,北京 100193)
雜草對ACCase抑制劑的抗性
郭 峰, 張朝賢, 黃紅娟, 魏守輝, 張 猛, 房 峰
(中國農業科學院植物保護研究所/中國農業科學院雜草害鼠生物學與治理重點開放實驗室,北京 100193)
以乙酰輔酶A羧化酶(ACCase)為作用靶標的除草劑是高效、選擇性的禾本科雜草除草劑,其在全球范圍內的廣泛、重復使用,導致了抗藥性雜草的發生和發展。到目前為止,已經在30個國家有37種抗此類除草劑的雜草生物型。抗藥性雜草嚴重威脅雜草治理和農業生產,由此引發的生態問題及糧食安全問題引起了廣泛的關注。文章概述了ACCase抑制劑抗藥性雜草的發生現狀,從雜草ACCase突變、代謝解毒等幾個方面綜述了雜草抗ACCase抑制劑的抗性機制,以期為ACCase抑制劑抗性研究提供參考。最后討論了阻止或延緩抗藥性發生的雜草管理措施。
ACCase抑制劑; 雜草; 抗藥性; 抗性機制; 管理措施
生物體有同質型(homomeric)和異質型(heteromeric)兩種類型的乙酰輔酶A羧化酶(acetyl-CoA carboxylase,ACCase,EC 6.4.1.2),全都包括生物素羧基載體蛋白(biotin carboxylase carrier protein,BCCP)、生物素羧化酶(biotin carboxylase,BC)、羧基轉移酶(carboxyltransferase,CT)[1]三個功能域,是一種以生物素為輔基,以HCO3-為羧基供體的羧化酶。ACCase在ATP的參與下將乙酰輔酶A羧化為丙二酸單酰輔酶A,而植物質體中脂肪酸生物合成及細胞質中脂肪酸延長、花青素和類黃酮等次生代謝產物合成都需要大量的丙二酸單酰輔酶A[2]。這個反應既是脂肪酸合成反應中的第一步反應,也是關鍵的反應步驟,同時也是限速步驟[3]。
1958年發現ACCase可以作為除草劑的作用靶標[4],目前針對該靶標的除草劑共有兩大類,芳氧苯氧基丙酸酯類(APP)和環己烯酮類(CHD)[5]。1971年,德國Hoechst公司首先開發出芳氧苯氧基丙酸酯類除草劑,并于第二年申請了禾草靈的專利。此后,日本石原產業公司發現將其一邊的苯環用吡啶取代后,吡啶取代物SL-501的除草活性為禾草靈的10倍[6]。從1975年開始,許多農藥公司在原有模式結構的基礎上,進行結構修飾和改造,相繼開發出一系列活性更高、選擇性更好的APP類除草劑,如吡氟禾草靈、精吡氟禾草靈、高效氟吡甲禾靈等。APP類除草劑存在立體異構,且這種異構體只有右旋體(R+)對ACCase有顯著活性。日本于1973年開發出第一個CHD類除草劑——禾草滅,而真正開創CHD類除草劑的是1981年開發出的高活性品種——烯禾啶。到目前為止,CHD類商品化除草劑主要有烯禾啶、烯草酮、禾草滅、噻草酮等10多個品種。
APP和CHD類除草劑主要通過抑制乙酰輔酶A的羧化,進而阻斷脂肪酸的合成,同時破壞膜的完整性,從而造成代謝物的滲漏和植物的快速死亡[7]。由于禾本科物種只存在同型ACCase,而非禾本科物種則同時含有同型和對ACCase抑制劑不敏感的異型兩種ACCase,所以ACCase抑制劑對非禾本科物種具有高度的選擇性,在全球范圍內被廣泛用來控制禾本科雜草[7]。除草劑的使用極大減輕了農民的負擔,但是對除草劑的過度依賴和使用也導致了抗藥性雜草種群的發生和發展,尤其是在ACCase抑制劑類這種作用靶標單一的除草劑更為嚴重,其全球抗藥性雜草生物型數量僅次于乙酰乳酸合成酶抑制劑類除草劑,是全球雜草抗藥性研究的熱點。近年來,由于分子生物學手段的廣泛應用,發現了與雜草對ACCase抑制劑敏感性改變至關重要的ACCase羧基轉移酶區域的7個氨基酸位點,闡明了雜草基于ACCase位點突變的抗性基礎,非靶標位點突變的抗性主要是代謝解毒酶活性的增強,雖然這類抗性機制普遍存在,但是目前對其還知之甚少。本文重點概述了ACCase抑制劑抗藥性雜草的發生現狀、雜草抗藥性機制,并討論了阻止或延緩抗藥性發生的雜草管理措施,以期為我國的雜草抗藥性研究提供參考。
1 雜草對ACCase抑制劑抗性的發生和發展

2 雜草對ACCase抑制劑抗性的作用機制
ACCase抑制劑類除草劑作為獨有的控制禾本科雜草的除草劑一直在作物田里被大量的施用,很快便導致能夠忍受ACCase抑制劑類除草劑田間劑量的禾本科雜草種群出現[7]。抗藥性雜草的發生不但增加了除草成本,也增加了環境壓力[11],闡明雜草對除草劑的誘導抗性特征與機制,延緩雜草抗性產生和延長ACCase抑制劑類除草劑使用年限,并為抗藥性雜草檢測和治理提供理論依據,都具有重要的實踐意義[12]。
2.1 作用靶標位點的突變


表1 部分國家2011年抗ACCase抑制劑類除草劑雜草生物型數量
2.2 代謝能力的增強
雜草種群的個體多實性、易變性及多型性是對除草劑產生抗性的內在因素[24]。除草劑在對雜草種群產生強大的篩選壓力的同時,也促使雜草體內做出一系列代謝反應,通過阻止除草劑活性分子到達靶標位點、削弱除草劑的毒害作用或者兩方面綜合作用,使得雜草對ACCase抑制劑類除草劑敏感性降低。雜草對除草劑的代謝不但改變了除草劑分子的化學性質,也改變了他們的功能,且代謝速度越快,對這些除草劑的抗性也就越高[15]。除草劑在植物體內代謝一般分為3個過程:一是通過氧化、水解或軛合等作用使得除草劑母體分子水溶性提高,毒性降低;二是將除草劑或其代謝產物軛合到GSTs、糖基或氨基酸上,從而進一步削弱母體分子的毒性;三是通過進入液泡或結合到細胞壁中將第二階段的代謝物進一步排除[25-26]。
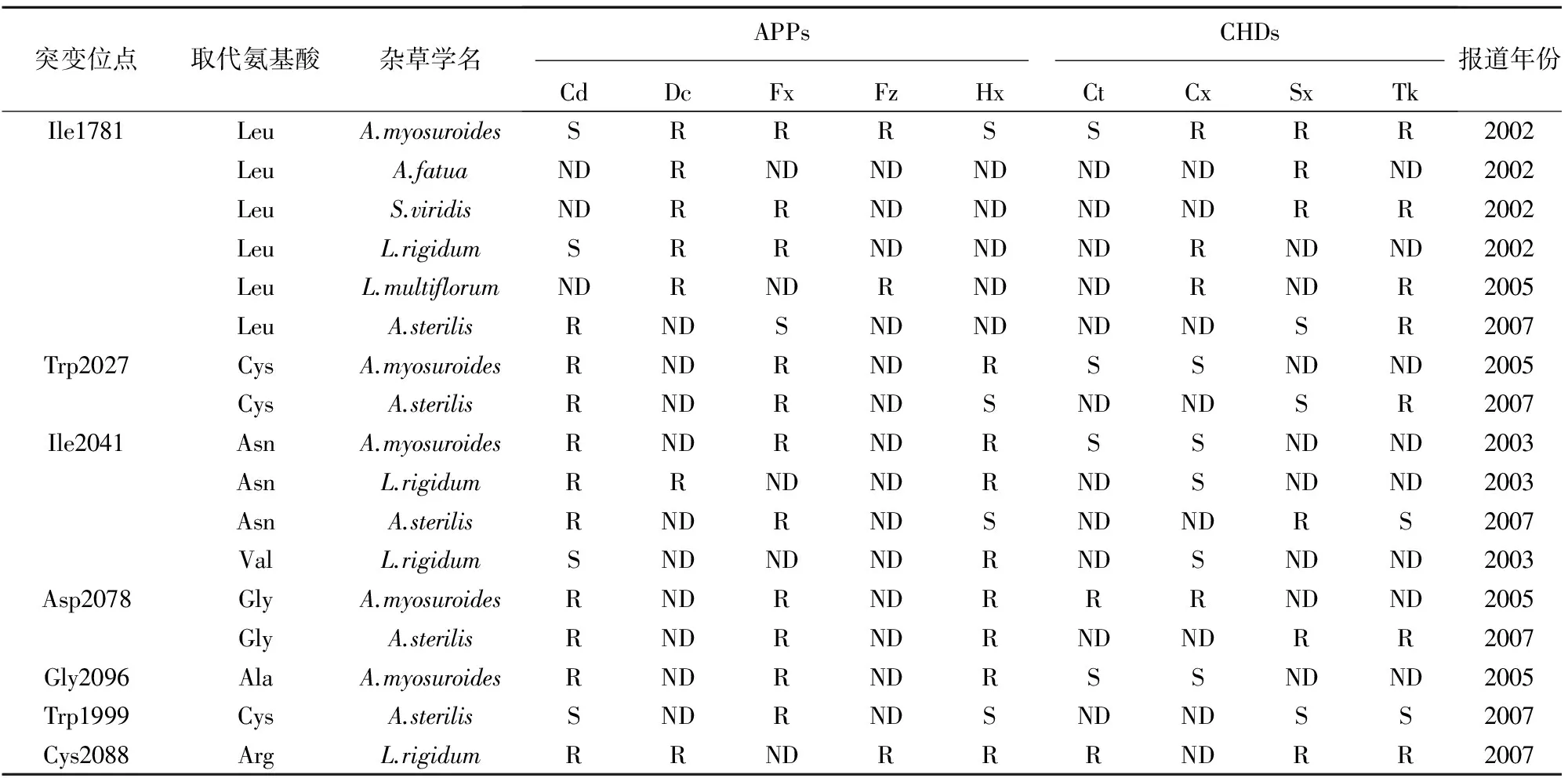
表2 抗ACCase抑制劑雜草的ACCase基因突變位點及交互抗性
2.3 其他方面的抗性機制
2.3.1 對ACCase抑制劑類除草劑具有天然耐性的同型質體ACCase 一些禾本科雜草的同型ACCase顯示出與胞質ACCase相類似的敏感性[15],其對ACCase抑制劑類除草劑具有天然的耐性。Catanzaro等發現,與敏感的狼尾草相比,4個藍羊茅栽培品種分別對吡氟禾草靈顯示70~88倍的抗性,對烯禾啶顯示216~422倍的抗性[34]。Herbert等通過ACCase測定,發現紅羊茅和早熟禾對喹禾靈、吡氟禾草靈和烯禾啶都存在較高的抗性[35]。
2.3.2 ACCase的過度表達 Bradley等運用放射性同位素的方法研究了對烯禾啶、精喹禾靈和精吡氟禾草靈存在較低抗性及敏感的假高粱對這些除草劑的吸收、運輸及代謝,發現抗性種群和敏感種群無明顯區別。在ACCase酶活性測定中發現,抗性種群和敏感種群有著相似的I50,不施用除草劑的情況下,抗性種群的ACCase比活力是敏感種群的2~3倍,添加除草劑的情況下,抗性種群ACCase比活力仍然明顯高于敏感種群。這些結果表明,假高粱對精喹禾靈和烯禾啶的抗性是由于ACCase的過度表達[9]。但是這種抗性機制不常見,僅僅在假高粱中有過報道[15]。
2.3.3 膜的去極化恢復 APP及CHD類除草劑能使原生質膜去極化,膜去極化導致細胞內pH值下降,Ca2+濃度上升,使細胞衰老死亡,而抗性生物型能通過復極化來降低傷害[36]。Hotum等發現抗AOPP除草劑的一些瑞士黑麥草和野燕麥生物型在去掉除草劑后可以恢復膜電位[37],Prado等認為質膜的復極化是瑞士黑麥草、多花黑麥草、鼠尾看麥娘、野燕麥等禾本科雜草對AOPP及CHD類除草劑抗性的一種機制[38]。雖然在一些研究中證實AOPP類除草劑對抗性及敏感性生物型的質膜電動勢(Em)影響不同,卻并沒有明顯的證據證明與質膜相關的除草劑作用位點或抗性機制的存在[39],這有可能是某種抗性機制引發的一個次要結果,而其本身并不是一種抗性機制[15]。
2.3.4 除草劑作用的屏蔽 Dinelli等對意大利兩種抗禾草靈黑麥草的研究排除了其抗藥性與作用位點突變、吸收減少或解毒增強有關,而認為其抗藥性是由于雜草對除草劑的屏蔽作用或與作用位點的隔離作用使除草劑不能到達作用位點,從而不能發揮除草作用[39]。
3 抗ACCase抑制劑類除草劑雜草抗性的治理
除草劑對于糧食安全具有重要的意義,也是機械化、規模化作物種植模式不可缺少的除草措施。但是新產品開發費用的不斷增加、新作用方式除草劑的缺乏及抗藥性雜草的發展,對化學防除這種最有效的除草方法威脅不斷增加[15]。在當前形勢下,我們必須采取有效措施阻止或延緩雜草抗藥性發生,延長除草劑使用年限,為農業增產及可持續發展提供保障。抗藥性雜草的治理是一個復雜的、持續的過程,單一的某一種措施并不能有效解決抗性問題,需要運用包括合理使用除草劑、農業防除、生物防除等各種管理措施, 綜合治理雜草并加強抗性的監測、抗性機制的研究,尤其是目前還不甚清楚的基于除草劑代謝的抗性機制、努力開發新的作用靶標的除草劑,來阻止和延緩抗性雜草的發生、發展。
3.1 抗性監測
施用除草劑后應檢查除草效果,區分不能防除的雜草是由于藥劑噴灑不均勻還是抗性所致,并拔除這些雜草。加強抗藥性雜草的監測預警,建立快速、準確的抗藥性測定方法,評估抗性發生的風險。
3.2 除草劑輪換使用
正確使用除草劑,避免連續使用同一種或同一類作用機制的除草劑。不同的生長季節,使用不同作用機制的除草劑。除草劑的混用也是延緩抗性發生的一種有效方法,但是具有相同殺草譜的除草劑最好分開使用。不要連續使用除草劑,以便使敏感雜草生物型在雜草群落組成中占據優勢,從而淘汰生態適應性較差的抗性生物型。
3.3 生物防除
1902年美國引進天敵昆蟲防除惡性雜草馬櫻丹,開創了雜草生物防除的先例,澳大利亞利用繡菌防除麥田雜草燈心草、粉苞苣成為國際上首例利用病原微生物防除雜草的成功例證[40]。生物防除對環境和作物安全、控制效果持久、防除成本低廉,是控制或延緩雜草抗藥性的有效措施[41]。
3.4 農業管理措施
合理的作物輪換制度可以改變田間雜草群落的組成,切斷一些惡性雜草與作物的聯系,從而可以使用不同類別的除草劑,降低抗性產生風險。土壤的淺耕不但可以翻松土壤,還能將先前萌發的雜草幼苗翻入地下,減少雜草的數量。薄膜覆蓋不但可以保水保肥,同時也可以抑制雜草的萌發。人工拔除能有效降低抗性生物型的數量。經常清洗農業機械設施,可以避免抗性雜草的傳播。
4 結語
絕大多數的抗性雜草發生在除草劑應用水平較高的發達國家。與發達國家相比,我國雜草抗藥性研究起步較晚,研究技術和儀器設備相對落后,正式報道的抗藥性雜草種類不多。除草劑混劑應用占有較大比例,一定程度上延緩了雜草抗藥性發展。同時我們不得不意識到,相對滯后的雜草抗藥性研究也一定程度上掩蓋了我國雜草抗藥性程度的真相。因此,我們必須關注我國雜草抗藥性的發展,尤其應關注多位點突變引起的雜草抗藥性研究。必須針對我國雜草抗藥性現狀,借鑒國外先進的抗藥性研究及監控技術,建立抗藥性雜草預測預報、快速檢測、抗性雜草的綜合治理體系,在雜草治理中充分發揮農藝措施、生態調控等措施的作用,科學合理地應用除草劑,延緩雜草抗藥性的發生,延長除草劑的使用壽命,以保障雜草的有效治理和農業可持續發展。
[1]Nikolau B J,Ohlrogge J B,Wurtele E S. Plant biotincontaining carboxylases[J]. Arch Biochem Biophys,2003,414:211-222.
[2]Harwood J L. Fatty acid metabolism[J]. Annu Rev Plant Physiol,1988,39:101-138.
[3]Herbert D,Walker K A,Price L J. Acetyl-CoA carboxylase-a graminicide target site[J]. Pestic Science,1997,50:67-71.
[4]Wakils J. A malonic acid derivative as an intermediate in fatty acid synthesis[J]. J Am Chem Soc,1958,80:64-65.
[5]Secor J,Cseke C,Owen J W. The discovery of the selective inhibition of acetyl-coenzyme A carboxylase activity by two classes of graminicides[J]. Brighton Crop Prot Conf Weeds,1989,3B:145-154.
[6]鮮 明. 高效芳氧苯氧羧酸類手性除草劑糖草酯的合成工藝研究[D]. 貴陽:貴州大學,2003.
[7]Devine M D,Shimabukuro R H. Resistance to acetyl coenzyme A carboxylase inhibiting herbicides[M]//Powles S B,Holtum J A M. Herbicide resistance in plants. Boca Raton:CRC Press,1994:141-169.
[8]Heap I M,Knight R. The occurrence of herbicide cross resistance in a population of annual ryegrass,Loliumrigidum,resistant to diclofop-methyl[J]. Australian Journal of Agricultural Research,1986,37(2):149-156.
[9]Bradley K W,Wu J R,Kriton K H,et al. The mechanism of resistance to aryl oxyphenoxypropionate and cyclohexanedione herbicides in a johnsongrass biotype[J]. Weed Science,2001,49:477-484.
[10]Heap I. International survey of herbicide resistance weeds[EB/OL]. [2011-3-15]. http:/www.weedscience.org.
[11]李宜慰,梅傳生,李永豐,等. 麥田菵草和日本看麥娘對綠磺隆抗性的初步研究[J]. 江蘇農業學報,1996,12(2):34-38.
[12]張朝賢,錢益新,胡祥恩,等. 農田化學除草與可持續發展農業[J]. 農藥,1998,37(4):8-12.
[13]De Prado R,Gonz?lez-Guriérrez J,Menéncez J,et al. Resistance to acetyl CoA carboxylase-inhibiting herbicides inLoliummultiflorum[J]. Weed Science,2000,48:311-318.
[14]Yu Q,Alberto C,Zheng M Q,et al. Divesity of acetyl-coenzyme A carboylase mutations in resistantLoliunpopulations:evaluation using clechodim[J]. Plant Physiology,2007,145:547-558.
[15]Délye C. Weed resistance to acetyl coenzyme A carboxylase inhibitors:an update[J]. Weed Science,2005,53:728-746.
[16]Shukla A,Dupont S,Devine M D. Resistance to ACCase-inhibitor herbicides in wild oat:evidence for target site-based resistance in two biotypes from Canada[J]. Pestic Biochem Physiol,1997,57:147-155.
[17]Volenberg D,Sroltenberg D. Altered acetyl-coenzyme A carboxylase confers resistance to clethodim,fluazifop and sethoxydim inSetariafaberiandDigitariasangrinalis[J]. Weed Research,2002,42:342-350.
[18]Délye C,Zhang X Q,Chalopin C,et al. An isoleucine residue within the carboxyl-transferase domain of multidomain acetyl-CoA carboxylase is a major determinant of sensitivity to aryloxyphenoxypropionate but not to cyclohexanedione inhibitors[J]. Plant Physiol,2003,132:1716-1723.
[19]Marles M A S,Devine M D,Hall J C. Herbicide resistance inSetariaviridisconferred by a less sensitive form of acetyl coenzyme A carboxylase[J]. Pestic Biochem Physiol,1993,46:7-14.
[20]Liu W J,Harrison D K,Chalupska D,et al. Single-site mutations in the carboxyltransferase domain of plastid acetyl-CoA carboxylase confer resisitance to grass-specific herbicides[J]. Proc Natl Acad Sci USA,2007,104:3267-3632.
[21]黃世霞,王慶亞,董立堯,等. 乙酰輔酶A羧化酶抑制劑類除草劑與雜草的抗藥性[J]. 雜草科學,2003(2):1-5.
[22]DePrado R,Osuna M D,Fischer A J. Resistance to ACCase inhibitor herbicides in a green foxtail(Setariaviridis) biotype in Europe[J]. Weed Science,2004,52:506-512.
[23]Neve P,Sadler J,Powles S B. Multiple herbicide resistance in a glyphosate-resistance rigid ryegrass(Loliumrigidum) population[J]. Weed Science,2004,52(6):920-928.
[24]黃春艷. 雜草抗藥性研究概況[J]. 黑龍江農業科學,1997(6):45-47.
[25]Kreuz K,Tommasini R,and Martinoia E. Old enzymes for a new job:Herbicide detoxification in plants[J]. Plant Physiol,1996,111:349-353.
[26]VanEerd L L,Hoagland R E,Zablotowicz R M,et al. Pesticide metabolism in plants and microorganisms[J]. Weed Science,2003,51:472-495
[27]歐曉明,唐德秀. 除草劑作用機理研究的新進展[J]. 世界農業,2000(10):28-30.
[28]Werck-Reichhart D,Feyereisen R. Cytochromes P450:a success story[J]. Genome Biol,2000,1(6):30031-30039.
[29]Reade J P H,Lucy J M,Andrew H C. A role for glutathioneS-transferases in resistance to herbicides in grasses[J]. Weed Science,2004,52:468-474.
[30]Brazier M,Cole D J,Adwards R E. O-glycosyltransferase activities towards phenolic natural products and xenobiotics in wheat and herbicide-resistance and berbicide-susceptible black-grass(Alopecurusmyosuroides)[J]. Phytochemistry,2002,59:149-156.
[31]Stephen M I C. Glutathione transferases in herbicide-resistant and herbicide-susceptible black-grass[J]. Pest Science,1997,51:244-250.
[32]Reade J P H,Cobb A H. New quick tests for herbicide resistance in black-grass(AlopecurusmyosuroidesHuds) based on increased glutathioneS-transferase activity and abundance[J]. Pest Management Science,2001,58:26-32.
[33]Kuk Y I,Wu J R,Derr J F,et al. Mechanism of fenoxaprop resistance in an accession of smooth crabgrass(Digirariaischoemum)[J]. Pesticide Biochemistry and Physiology,1999,64(2):112-123.
[34]Catanzaro C J,Burton J D,Skroch W A. Graminicide resistance of acetyl-CoA carboxylase from ornamental grasses[J]. Pestic Biochem Physiol,1993,45:147-153.
[35]Herbert D,Cole D J,Pallett K E,et al. Susceptibilities of different rest systems from maize(Zeamays),PoaannuaandFestucarubrato herbicides that inhibit the enzyme acetyl-coenzyme A carboxylase[J]. Pestic Biochem Physiol,1996,55:129-139.
[36]Incledon B J,Hall J C. Acetyl-coenzyme A carboxylase:quarternary structure and inhibition by graminicidal herbicides[J]. Pestic Biochem Physiol,1997,57:255-271.
[37]Holtum J A M,Hausler R E,Devine M D. Recorvy of transmembrane potentials in plants resistant to aryloxyphenoxypropionate herbicides:a phenomenom awaiting explanation[J]. Weed Science,1994,42:293-301.
[38]De Prado J L,De Prado R A,Shimabukuro R H. The effect of diclofop on membrane potential,ethylene induction,and herbicide phytotoxicity in resistance and susceptible biotypes of grasses[J]. Pesticide Biochemistry and Physiology,1999,63:1-14.
[39]Dinelli G,Bonetti A,Marotti I,et al. Possible involvement of herbicide sequestration in the resistance to diclofop-methyl in Italian biotypes ofLoliunspp.[J]. Pesticide Biochemistry and Physiology,2005,81(1):1-12.
[40]萬方浩,王 韌. 世界雜草生防的歷史成就及我國雜草生防的現狀與建議[J]. 生物防治通報,1991,7(2):81-87.
[41]張希福,熊建偉,尹 健. 雜草生物防治的現狀與展望[J]. 河南職業技術師范學院學報,1997,25(4):8-14.
ResistanceofWeedtoACCase-InhibitingHerbicides
GUO Feng,ZHANG Chao-xian,HUANG Hong-juan,WEI Shou-hui,ZHANG Meng,FANG Feng
(Institute of Plant Protection,Chinese Academy of Agricultural Sciences/ Key Laboratory of Weed and Rodent Biology and Management,Beijing 100193,China)
Hebicides targeting acetyl-CoA carboxylase(ACCase) are effective and selective graminicides. Their widespread and repeated application in worldwide resulted in occurrence and evolution of resistant weed species. There have been 37 weed biotypes resistant to ACCase herbicides in 30 countries by now. The herbicide resistant weeds threatted to weed management and crop production,and the consequences leading to biological damage and food safety caused extensive concern in the world. This review summarized the current status of ACCase-inhibiting herbicide resistant weeds and the major mechanisms of resistance involed in the ACCase mutations,metabolic detoxification and so on. In order to provide useful information and references for researches of ACCase-inhibiting herbicide resistance. Potential management strategy for preventing and delaying the herbicide resistance evolution of weeds was discussed in the end.
ACCase-inhibiting herbicides; weeds; herbicide resistance; mechanism of resistance; management strategy
S-1
A
1003-935X(2011)03-0001-06
郭 峰,張朝賢,黃紅娟,等. 雜草對ACCase抑制劑的抗性[J]. 雜草科學,2011,29(3):1-6.
2011-05-12
國家轉基因生物新品種培育重大專項(編號:2009ZX08012-025B)。
郭 峰(1985—),男,山西大同人,碩士研究生,主要從事雜草抗藥性研究。E-mail:guofeng5866@126.com。
張朝賢,博士,研究員,主要從事雜草科學研究。Tel:(010)62815937; E-mail:cxzhang@wssc.org.cn。
猜你喜歡
科教新報(2022年22期)2022-07-02 12:34:28
今日農業(2021年5期)2021-11-27 17:22:19
高中生學習·閱讀與寫作(2019年2期)2019-09-10 07:22:44
今日農業(2019年15期)2019-01-03 12:11:33
現代園藝(2017年19期)2018-01-19 02:50:21
現代農業(2016年5期)2016-02-28 18:42:36
營銷界(2015年23期)2015-02-28 22:06:18
現代農業(2015年5期)2015-02-28 18:40:49
新疆農墾科技(2014年3期)2014-02-28 19:19:34
中國質量與標準導報(2013年8期)2013-03-11 19:54:07
