高溫處理對脫脂豆粕中大豆分離蛋白結構的影響
2011-11-28 02:28:46黃友如陳義勇朱東興王嘆玉
中國糧油學報 2011年10期
關鍵詞:大豆
黃友如 陳義勇 朱東興 趙 陽 王嘆玉
高溫處理對脫脂豆粕中大豆分離蛋白結構的影響
黃友如 陳義勇 朱東興 趙 陽 王嘆玉
(常熟理工學院生物與食品工程學院,常熟 215500)
以不同溫度處理的脫脂豆粕為原料,制備大豆分離蛋白,通過紫外光譜、熒光光譜、電泳及濁度測定等方法探討了高溫處理對脫脂豆粕中大豆分離蛋白結構的影響,結果表明:熱處理可促使蛋白多肽鏈局部展開,色氨酸等殘基暴露并部分氧化,產生一些新熒光物質;凝膠電泳分析顯示,熱處理樣品的高分子區域條帶漸趨顯著,各亞基條帶呈現擴散趨勢,蛋白質相對分子質量分布范圍變寬;濁度分析表明,隨著樣品熱處理溫度升高,大豆分離蛋白的混濁度增加。
高溫處理 脫脂豆粕 大豆分離蛋白 結構
工業化生產的低溫脫脂豆粕中含有許多殘留脂質,在高溫貯藏過程中,殘留脂質尤其是多不飽和脂質的氧化將產生脂質自由基及其降解產物,這些衍生自由基及其降解產物可與豆粕中的蛋白質發生反應,改變蛋白質的結構,進而影響以此為原料制備的大豆蛋白的品質[1]。因此,對脫脂豆粕進行高溫處理,研究不同溫度條件下殘留脂質的氧化情況及其對蛋白質結構的影響,對延長大豆蛋白食品的貨架期、保證產品的品質與營養價值意義深遠。
本研究主要以低溫脫脂大豆粕為原料,分別采用室溫(30℃)、40、50、60℃4種不同的溫度對已粉碎的脫脂豆粕進行18 h的溫度處理,再以“堿提酸沉”工藝制備大豆分離蛋白,采用電泳及光譜分析方法如混濁度、紫外吸收光譜、280 nm激發的大豆分離蛋白內源性熒光光譜、激發光譜和發射光譜,SDSPAGE和原態PAGE等,分析高溫處理對脫脂豆粕中蛋白質的結構影響。
1 材料與方法
1.1 試驗材料、試劑和儀器
低溫脫脂豆粕由東海糧油工業(張家港)有限公司提供,其他試劑均為分析純。UV2450型紫外可見分光光度計、RF-5301PC型熒光分光光度計:日本島津公司。
1.2 低溫脫脂豆粕的高溫處理
稱取低溫脫脂豆粕1 000 g,粉碎后100%過60目標準篩,將其分成4等份,一份置于室溫(30℃)下做對照,另3份置于烘箱中,分別采用40、50、60℃3種不同溫度處理18 h(處理厚度為2 mm),取出冷卻后,分別放入密封袋中密封保存。
1.3 脂肪氧合酶活力的測定
脂肪氧合酶活力的測定參照黃友如等[2]的方法。
1.4 大豆分離蛋白的制備
將高溫處理后的脫脂豆粕粉按料液比1∶10的比例與水混合,溫度 30℃,低速攪拌(30~35 r/min),用1.0 mol/L NaOH 溶液調至 pH 7.5,繼續攪拌30 min。懸浮液離心15 min(2 000 r/min),回收上清液,沉淀(豆渣)棄去。上清液邊攪拌邊加入1.0 mol/L HCl溶液(30 ~35 r/min),調至pH 4.5,繼續攪拌 10 min后,懸浮液離心 15 min(2 000 r/min),棄去上清液(大豆乳清),取沉淀水洗,水洗后的蛋白沉淀分散于去離子水中并用1.0 mol/L NaOH調至pH 7.0。所得蛋白溶液冷凍干燥后粉碎并過80目標準篩。
1.5 大豆分離蛋白的紫外吸收光譜
將大豆分離蛋白溶于0.01 mol/L pH 7.0的磷酸鹽緩沖液中,配制成2.0 mg/mL的蛋白質溶液,3 000 r/min離心10 min,以0.01 mol/L pH 7.0 的磷酸鹽緩沖液作參比,作紫外 -可見掃描,速度10 nm/s,范圍200~800 nm之間。
1.6 280 nm激發的大豆分離蛋白內源性熒光光譜
將大豆分離蛋白溶于0.01 mol/L pH 7.0的磷酸鹽緩沖液中,配制成1.0 mg/mL的蛋白質溶液,3 000 r/min離心10 min,以pH 7.0的磷酸鹽緩沖液作參比,進行熒光測定,激發波長280 nm,發射光譜掃描范圍在300~800 nm之間。
1.7 大豆分離蛋白的激發光譜和發射光譜測定[3]
將大豆分離蛋白溶于0.01mol/L pH 7.0的磷酸鹽緩沖液中,配制成2.0 mg/mL的蛋白質溶液,3 000 r/min離心10 min,以pH 7.0的磷酸鹽緩沖液作參比進行熒光測定,發射光譜:激發波長固定在355 nm,掃描范圍300~800 nm;激發光譜:發射波長固定在440 nm,掃描范圍220~800 nm。
1.8 大豆分離蛋白的SDS聚丙烯酰胺凝膠電泳(SDS - PAGE)[4]
分離膠12.75%;濃縮膠4%;電極緩沖液含0.05 mol/L Tris,0.384 mol/L 甘 氨 酸,0.1% SDS(pH 8.3);電泳樣品用樣品溶解液(內含2%SDS,5%β - 巰基乙醇,10% 甘油,0.02% 溴酚藍,0.01 mol/L pH 8.0 Tris-HCl緩沖液)配制成蛋白濃度為1 mg/mL。電泳采用0.75 mm凝膠板;上樣量為10 μL;開始電泳時電流為10 mA,待樣品進入分離膠后改為20 mA;考瑪斯亮藍染色。
1.9 大豆分離蛋白PAGE原態電泳
分離膠 5%;濃縮膠 4%;電極緩沖液含0.05 mol/L Tris,0.384 mol/L 甘氨酸,pH 8.3;電泳樣品用樣品溶解液(內含10%甘油,0.02%溴酚藍,0.01 mol/L pH 8.0 Tris-HCl緩沖液)配制成蛋白濃度為1 mg/mL。電泳采用0.75 mm凝膠板;上樣量為10μL;樣品直接溶于樣品溶解液中,在1 500×g下離心10 min后樣品溶解液不經過加熱處理,直接電泳進樣;開始電泳時電流為10 mA,待樣品進入分離膠后改為20 mA;考瑪斯亮藍染色。
2.0 大豆分離蛋白溶液混濁度的測定[5]
將大豆分離蛋白溶于0.01 mol/L pH 7.0的磷酸鹽緩沖液中,室溫下用磁力攪拌器攪拌60 min,分別配制成 20、40、60、80、100 mg/mL 的溶液,使用分光光度計在600 nm下測定其吸光度。
2 結果與分析
2.1 不同溫度處理下脫脂豆粕中脂肪氧合酶活力
表1為不同溫度處理下脫脂豆粕中脂肪氧合酶的活力。與室溫(30℃)條件下貯藏的脫脂豆粕相比較,經40、50、60℃處理后的脫脂豆粕,其脂肪氧合酶活力隨熱處理溫度上升均有所降低,但各樣品的脂肪氧合酶活力在P≥0.05水平上并沒有明顯差異,說明在此溫度范圍內,粉末狀脫脂豆粕(含水量為10.32%)中脂肪氧合酶的熱穩定性較好,在其后的分離蛋白制備過程中,脂肪氧合酶對大豆分離蛋白結構的貢獻相同。
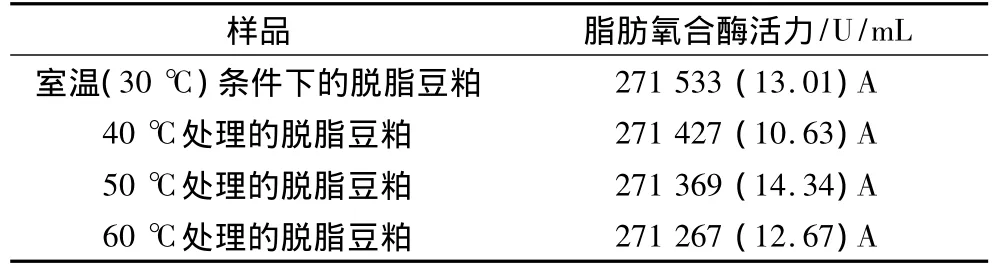
表1 不同溫度處理下脫脂豆粕中脂肪氧合酶的活力
2.2 大豆分離蛋白的紫外吸收光譜
紫外光譜可用來研究蛋白側鏈的變化(如酪氨酸殘基的裸露程度),探討熱處理對大豆蛋白三級結構的影響[6]。圖1為改性大豆分離蛋白的紫外可見光譜。經過40、50、60℃處理后的樣品在280 nm附近的吸收峰強度增加,峰型變窄,說明具有紫外吸收的芳香族氨基酸殘基如酪氨酸殘基等因熱處理而暴露在蛋白質分子的表面,發色基團所處的環境由非極性向極性變化。與對照樣品a相比,樣品 b,c,d的峰位沒有位移,峰型無明顯區別,但在280 nm附近的吸收峰都逐漸增強。我們知道,一般疏水性殘基大都埋藏于蛋白質分子內部[6],其紫外吸收增加表明經不同溫度處理后的大豆蛋白構象的變化趨勢基本相同,熱處理促使蛋白多肽鏈局部展開。
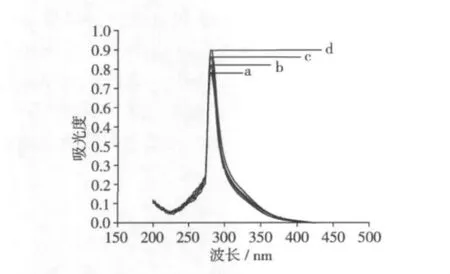
圖1 大豆分離蛋白的紫外可見光譜
2.3 大豆分離蛋白內源性熒光光譜
圖2為大豆蛋白樣品的內源性熒光光譜,激發波長為280 nm。對照樣品以及分別經過40、50、60℃處理后樣品的熒光峰位(λmax)分別位于333、338、334、336 nm,相對熒光強度分別為 718、873、913和847。可見樣品的熒光峰位(λmax)和強度隨熱處理溫度的不同而不同。
在280 nm激發的大豆蛋白樣品熒光發射光譜主要是由色氨酸和酪氨酸殘基所發射的[2,5],又名內源性熒光光譜。和其他含色氨酸和酪氨酸殘基的球蛋白一樣,由于其分子中從酪氨酸殘基到色氨酸殘基之間發生了能量轉移,從而導致了酪氨酸殘基的熒光熄滅和色氨酸殘基的熒光增加。因此大豆球蛋白的熒光峰實際上是色氨酸殘基的熒光峰,其峰位在325 ~350 nm 波長之間[7]。
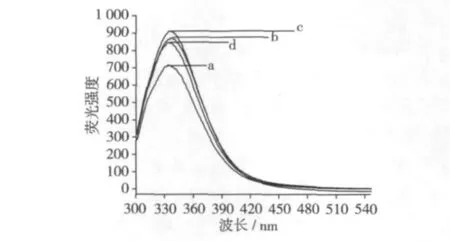
圖2 大豆分離蛋白的內源性熒光光譜(280 nm激發)
熱處理后樣品熒光強度和λmax的改變反映了色氨酸殘基被氧化的程度和所處微環境的變化[7-8]。與對照樣品相比,經過40、50、60℃處理后樣品的λmax向長波方向發生了不同程度的位移(紅移),表明色氨酸殘基暴露于蛋白質分子表面,說明蛋白質的疏水區域發生了局部的改變,生色基團如色氨酸殘基所處環境由非極性向極性轉化,這可能是由于豆粕的高溫處理導致蛋白質分子部分解折疊造成的;而熒光強度增加,說明雖然高溫處理,但此時的色氨酸殘基基本沒有被氧化,或被氧化的程度較低,為蛋白質解折疊暴露的色氨酸殘基所補充。值得注意的是,熒光峰位(λmax)和強度的這種變化并非線性走向。λmax改變最明顯的是經過40℃處理的樣品,而熒光強度改變最明顯的卻是經過50℃處理的樣品。這似乎也說明樣品經40、50℃處理時,色氨酸等疏水性氨基酸殘基雖然暴露但極少被氧化,而當處理溫度達60℃時,暴露的色氨酸等疏水性氨基酸殘基開始發生了氧化,致使位于蛋白質分子表面的色氨酸殘基減少,其熒光強度降低。Hidalgo等[8]在研究β-乳球蛋白與亞油酸13-氫過氧化物的相互作用時也發現,氧化β-乳球蛋白的λmax向短波方向移動且熒光強度降低的現象。這里脫脂豆粕經高溫處理后,蛋白樣品內源性熒光光譜的變化可能與色氨酸殘基的暴露與氧化有關。蛋白質解折疊暴露在表面的色氨酸殘基可加速其氧化的進程 。
2.4 大豆分離蛋白激發光譜和發射光譜分析比較
圖3為經過熱處理的4種大豆分離蛋白的激發光譜,4種樣品的激發光譜均有兩個峰,分別位于240 nm和280 nm附近,其中在240 nm附近的峰強度較小,峰位也基本沒有偏移,但峰強度隨處理溫度升高增加較大;與240 nm附近的峰強度相比,各樣品在280 nm處的峰強度最大,但峰強度隨處理溫度升高增加的幅度較小,峰位也有所偏移。
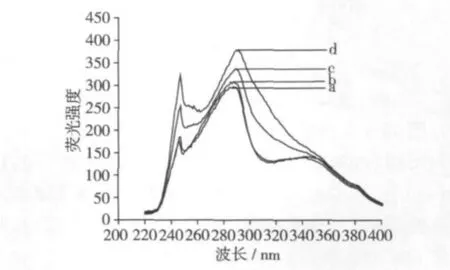
圖3 大豆分離蛋白的激發光譜(440 nm發射)
激發光譜280 nm附近的峰主要歸屬于蛋白質中芳族基團的吸收,4種樣品的激發光譜在280 nm附近峰強度隨處理溫度升高而增強,說明脫脂豆粕在高溫處理過程中大豆蛋白的芳族氨基酸殘基發生了氧化,其氧化產物可能和豆粕中的其他組分發生反應,生成某些新的熒光物質,且這些含有新熒光物質的相互作用產物,在其后的分離蛋白提取過程中,多不溶于水溶液而留存于分離蛋白中。正是這些新熒光化合物光譜疊加造成了激發光譜的變化。黃友如等[3,10]研究認為,形成新熒光物質原因有兩個:一是自由基(脂肪氧合酶催化脫脂豆粕中的殘留脂質如亞油酸過氧化產生的脂質過氧化自由基,·OH等)介導的C—C或C—N交聯(如二酪氨酸交聯);二是非自由基引發的反應機制,即脫脂豆粕貯藏期間殘留脂質的自氧化和/或蛋白浸提過程中脂肪氧合酶催化脫脂豆粕中的殘留脂質氧化降解產生的次生產物如醛類化合物與蛋白質賴氨酸等殘基的側鏈基團反應形成。前面脂肪氧合酶的活力分析也表明,不同溫度處理的脫脂豆粕中脂肪氧合酶的活力在P≥0.05水平上并沒有明顯差異,說明在此處理溫度范圍內,脂肪氧合酶的熱穩定性較好,在其后的分離蛋白制備過程中,脂肪氧合酶對大豆分離蛋白結構的影響一致。因此,本試驗中產生的新熒光化合物可能源于脫脂豆粕貯藏期間熱處理造成的殘留脂質自氧化降解產生的次生產物與蛋白質賴氨酸等殘基的側鏈基團反應形成。
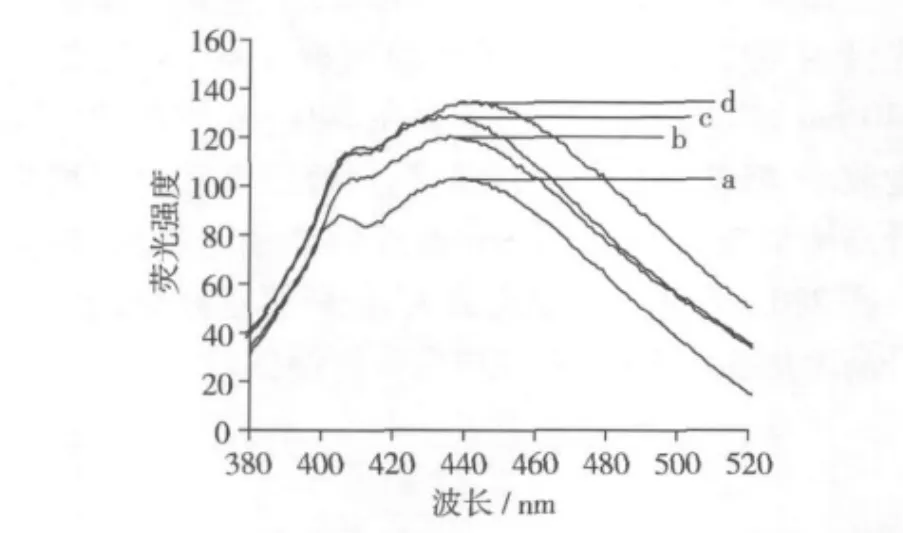
圖4 大豆分離蛋白的發射光譜(355 nm激發)
圖4為大豆分離蛋白的發射光譜(355 nm激發),可以看出,經過不同溫度處理后的4種樣品在440 nm附近均有最大的發射波長,從40℃到60℃的熱處理,處理溫度越高,樣品在440 nm附近發射光譜的峰強度越強。
Liang[1]在研究高溫(60℃)貯藏期間的大豆蛋白與大豆油相互作用時發現,發生氧化的大豆蛋白樣品的熒光光譜也出現上述類似的變化,其熒光光譜最大激發波長在355 nm,最大發射波長在440 nm。與對照樣品相比,經過40、50、60℃處理后的樣品,伴隨著色氨酸殘基的氧化,內源性熒光強度先增加而后又降低(圖2),而激發光譜280 nm附近峰強度和發射光譜440 nm附近峰強度增強,說明在脫脂豆粕的高溫處理期間產生了一些新的熒光物質,且這些新產生的熒光物質部分具有水不溶性的特點,在隨后的“堿提酸沉”工藝制備大豆分離蛋白過程中滯留于蛋白體系中。大豆蛋白激發光譜和發射光譜的這種變化應是這些新形成的熒光化合物光譜疊加造成的。
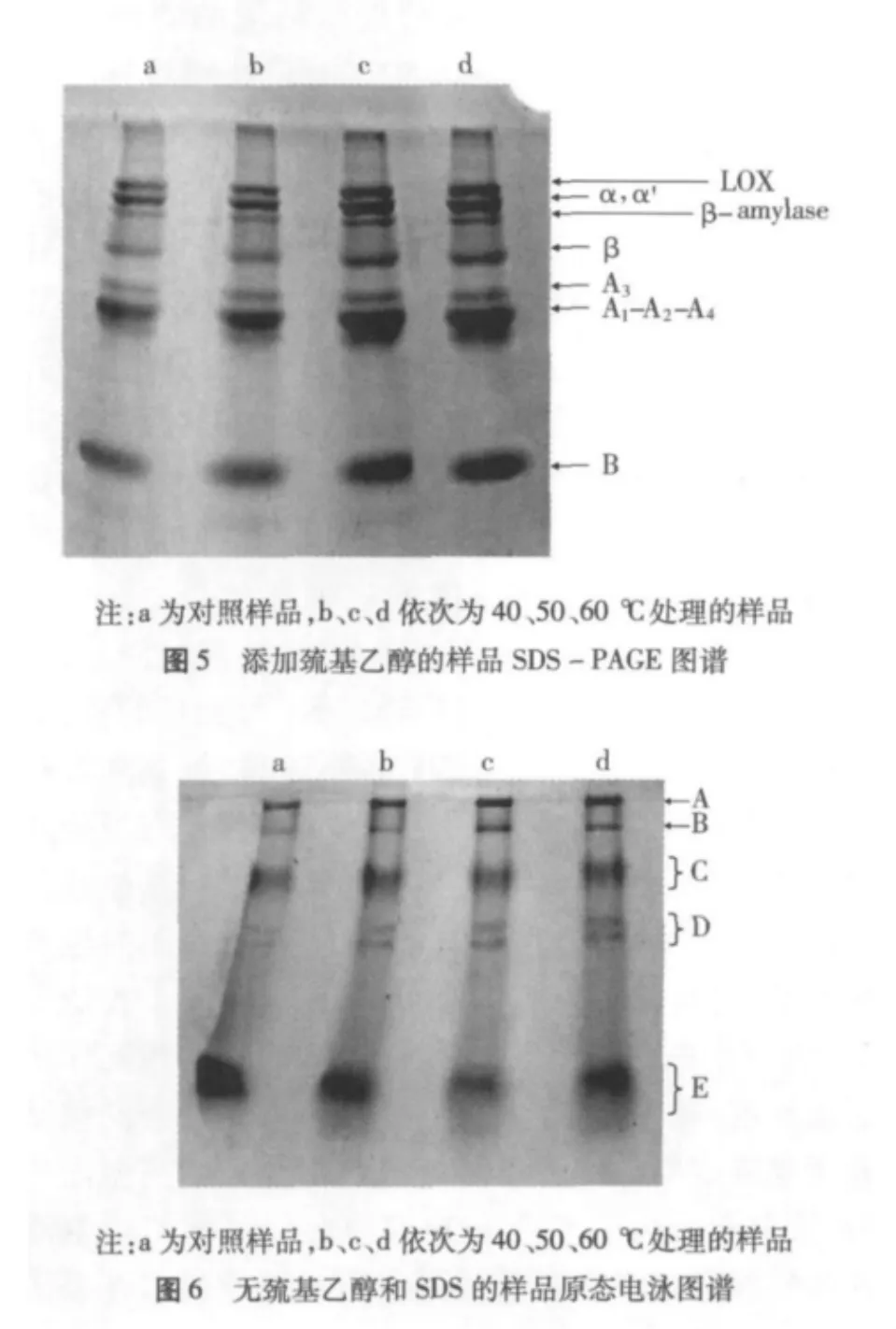
2.5 大豆分離蛋白凝膠電泳
為了確定脫脂豆粕經高溫處理后,大豆蛋白質的各亞基是否發生了變化,我們進行了SDS-PAGE電泳分析,結果見圖5。可以看出,對照樣品呈現為典型的大豆蛋白電泳圖譜[2]。相比之下,脫脂豆粕經40、50、60℃處理后的蛋白電泳圖譜有了明顯的變化。7S蛋白的α,α'和 β3個特征性亞基條帶著色變深且有融合的現象。11S蛋白酸性亞基A的電泳行為也有類似的變化,但11S蛋白的堿性亞基B條帶變寬了。此外,在高溫處理后樣品電泳圖譜的高分子區域有明顯可見的著色區域。這些事實表明,在脫脂豆粕的高溫處理過程中,除了發生蛋白質的部分氨基酸殘基的氧化外,脫脂豆粕中的殘留脂質也可能發生了氧化,處理樣品的擴散條帶可能是屬于一種共價類型的脂-蛋白[10]。至于有大豆蛋白亞基之間共價交聯的產生,也是可能發生的。處理樣品電泳圖譜中7S區域的變化很有可能是由于在某些蛋白亞基之間形成了共價交聯,如A-B,β-B或β-A等,結果造成該區域電泳條帶的重疊,著色強度增加。且這種共價交聯不屬于—S—S—,應是—C—C—或—C—N—之類,因為電泳樣品溶解液中含有β-巰基乙醇,該試劑可打開電泳樣品中的—S—S—。值得注意的是上述反應是隨機的,因而蛋白質各亞基之間以及蛋白質各亞基和脂質之間的交聯也是隨機的,結果造成電泳條帶的擴散,相對分子質量分布范圍變寬。的確,上述分析僅是理論推導,圖5反映的樣品亞基含量增加信息,其確切歸屬有待進一步證實,相關的研究正在進行中。
脫脂豆粕經高溫處理后蛋白質的交聯情況在原態電泳圖譜中得到充分的體現。圖6為沒有添加巰基乙醇和SDS的樣品原態電泳圖譜。從6可以看出,各樣品的原態電泳圖譜主要呈現A,B,C,D和E 5條區帶。相比之下,隨著處理溫度的升高,A,B,D3條區帶著色漸趨明顯,C帶變化不大,而E帶(低分子區域)漸寬但著色漸趨模糊。這些結果表明脫脂豆粕經高溫處理后,蛋白質分子不僅有共價的交聯,也有斷裂現象的產生。交聯會導致蛋白質顆粒粒度變大,溶液濁度增加,而斷裂則會使其溶解度增加。
2.6 大豆分離蛋白的混濁度分析
圖7為不同濃度下大豆分離蛋白的混濁度,可見每一樣品的混濁度隨蛋白質濃度的增加而升高,在同一蛋白質濃度下,熱處理溫度升高,大豆分離蛋白的混濁度增加。處理后樣品溶液混濁度增加可能是蛋白質分子共價交聯引起蛋白質顆粒粒徑增加所致。
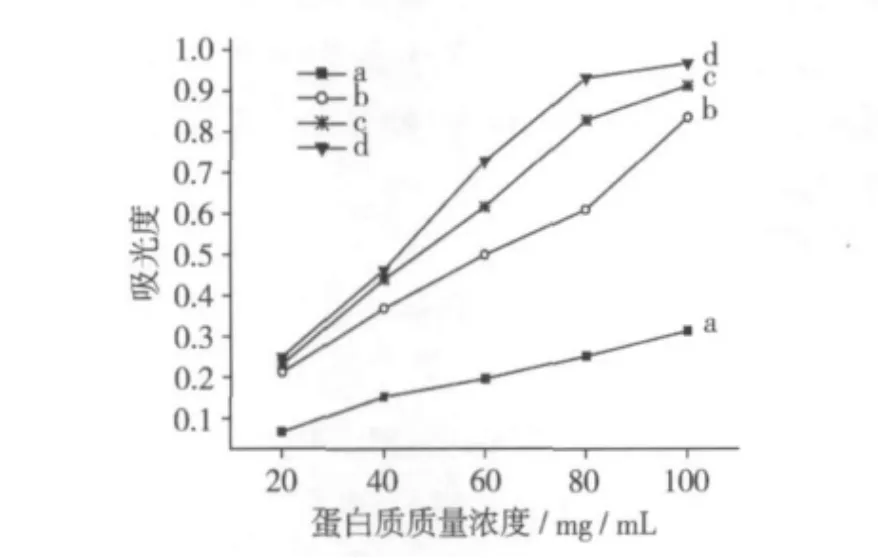
圖7 大豆分離蛋白的混濁度
3 討論與結論
脫脂豆粕貯藏期間因殘留脂質氧化導致大豆蛋白結構、功能及營養價值的變化現象已引起人們的關注,但相關研究較少[1-3,5,11-13]。本試驗以低溫脫脂大豆粕為原料,采用不同溫度(40~60℃)處理,以“堿提酸沉”工藝制備大豆分離蛋白,應用紫外光譜、熒光光譜、電泳及濁度測定等方法對經不同溫度處理后制備的大豆分離蛋白的結構進行了分析與表征。初步的結果表明,脫脂豆粕的高溫處理可促使蛋白多肽鏈局部展開,原來位于蛋白質內部的疏水性氨基酸殘基如色氨酸、酪氨酸、苯丙氨酸等較多地裸露在蛋白質分子的表面,從而造成蛋白樣品的紫外與熒光特征峰強度增強。伴隨著蛋白質多肽鏈的局部展開,暴露的疏水性氨基酸殘基部分發生了氧化,產生某些新熒光物質,且這些新產生的熒光物質部分具有水不溶性的特點,使得大豆蛋白的激發光譜280 nm附近和發射光譜440 nm附近的峰強度增強。其外,原料脫脂豆粕的干熱處理亦可造成殘留脂質的氧化,蛋白質各亞基和脂質之間可能發生交聯,蛋白質各亞基之間不僅有共價的交聯,也有斷裂現象的產生,從而造成蛋白凝膠電泳圖譜的高分子區域條帶隨熱處理溫度的升高而漸趨明顯,電泳條帶亦呈現擴散趨勢,相對分子質量分布范圍變寬。黃友如等[3,10]在脂肪氧合酶催化亞油酸氧化與大豆蛋白相互作用的研究中也發現類似的現象,并將其解釋為自由基介導的C—C或C—N交聯或非自由基引發的反應機制。但本試驗的結果證明,不同溫度處理的脫脂豆粕中脂肪氧合酶的活力在P≥0.05水平上并無明顯差異,說明在此處理溫度(40~60℃)范圍內,脂肪氧合酶的熱穩定性較好,在其后的分離蛋白制備過程中,脂肪氧合酶誘導的自由基反應機制對大豆分離蛋白結構的貢獻相同。因此,本試驗中大豆分離蛋白結構的變化當歸因于脫脂豆粕貯藏期間不同溫度處理造成的殘留脂質自氧化。至于產生的新熒光化合物,因其成分復雜,純化與定量研究有一定的難度,目前尚停留在定性階段,其確切歸屬尚待進一步證實。
[1]Liang J H.Fluorescence due to interactions of oxidizing soybean oil and soy proteins.Food Chemistry[J],1999,66:103 -108
[2]黃友如,華欲飛,裘愛泳.脂質氧化誘導的大豆蛋白質聚集機理的研究[J].中國糧油學報,2006,21(1):80 -87
[3]黃友如,華欲飛,盧祥云,等.脂肪氧合酶催化亞油酸氧化與大豆蛋白相互作用的熒光光譜分析[J].食品科學,2008,29(5):134 -139
[4]郭堯軍.蛋白質電泳實驗技術[M].北京:科學出版社,1999:123-160
[5]Huang Y R,Hua Y F,Qiu A Y.Soybean protein aggregation induced by lipoxygenase catalyzed linoleic acid oxidation[J].Food Research International,2006,39(2):240 - 249
[6]Yamagishi T,Yamauchi F,Shibasaki K.State of aromatic acid residues in soybean 11S globulin heated in the presence of N - Ethylmaleimide by derivative spectrophotometry[J].Agricultural and Biological Chemistry,1981,45(2),459 -467
[7]Kalapathy U,Hettiarachchy N S,Rhee K C.Effect of drying methods on molecular properties and functionalities of disulfide bond - cleaved soy proteins[J].Journal of the American Oil Chemists'Society,1997,74(3):195 -199
[8]Hidalgo F J,Kinsella JE.Changes induced in β -lactoglobulin Bfollowing interactions with linoleic acid 13-hydroperoxide[J].Journal of Agricultural and Food Chemistry,1989,37:860-866
[9]Papiz M Z,Sawyer L,Eliopoulos E E,et al.The structure of β-lactoglobulin and its similarity to plasma retinal-binding protein[J].Nature,1986,324:383
[10]黃友如.脂肪氧合酶催化亞油酸誘導大豆蛋白聚集機理[D].無錫:江南大學,2006
[11]Huang Y R,Yu D,Hua Y F,et al.Detection of free radical transfer in lipoxygenase I-B-catalyzed linoleic acid-soybean proteins interaction by electron spin resonance spectroscopy(ESR)[J].Journal of Agricultural and Food Chemistry,2006,54(24):9216 -9220
[12]Wu W,Zhang C M,Kong X Z,et al.Oxidative modification of soy protein by peroxyl radicals[J].Food Chemistry,2009,116(1):295 -301
[13]Wu W,Wu X J,Hua Y F.Structural modification of soy protein by the lipid peroxidation product acrolein[J].LWT -Food Science and Tech-nology,2010,43(1):133 -140.
Influence of High Temperature on Structure of Soy Protein Isolates Prepared with Defatted Soy Flour
Huang Youru Chen Yiyong Zhu Dongxing Zhao Yang Wang Tanyu
(School of Biological Science and Food Engineering,Changshu Institute of Tech-nology,Changshu 215500)
Defatted soy flour was treated with different high temperatures and soy protein isolates were prepared with the treated flour.The protein structure of the samples were studied by UV - vis spectra,fluorescence spectroscopy,polyacrylamide gel electrophoresis and turbidity.The results showed that polypeptide chains of the protein were partially unfolded after the dry heating,tryptophan residues of the proteins were exposed and oxidized along with unfolded polypeptide chains,and some new fluorescent compounds were produced during the dry heating.Polyacrylamide gel electrophoresis revealed that the bands of the heated samples in high molecular area became more and more notable,each subunit band appeared diffused and its molecular weight distribution were broadened.Turbidity analysis showed that the turbidity of the samples increased along the heating temperature.
high temperature treatment,defatted soy flour,soy protein isolates,structure
TS201.2+1
A
1003-0174(2011)10-0051-06
江蘇省高校自然科學研究項目(10KJB550001),蘇州市科技計劃項目(SYND201003)
2010-12-07
黃友如,男,1966年出生,博士,副教授,食品科學與工程
猜你喜歡
農業科技通訊(2023年1期)2023-02-12 07:09:18
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農產品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
讀寫算(中)(2015年10期)2015-11-07 07:24:12
