抗人DR5單克隆抗體誘導 Jurkat和 U937細胞凋亡的線粒體信號通路①
2012-01-23 12:03:22李淑蓮張舒曼劉廣超馬遠方
中國免疫學雜志 2012年12期
李淑蓮 蔡 靜 張舒曼 劉廣超 馬遠方
(河南大學細胞與分子免疫學重點實驗室河南大學免疫學研究所,開封475004)
腫瘤壞死因子相關的凋亡誘導配體(Tumor necrosisfactorrelated apoptosisinducing ligand,TRAIL)為TNF家族的成員之一,TRAIL有5種受體,4種膜結合受體(TRAIL-R1/DR4、TRAIL-R2/DR5、TRAIL-R3/DcR1、TRAIL-R4/DcR2)和 1 種可溶性受體 OPG[1],其中 TRAIL-R1/DR4 和 TRAIL-R2/DR5胞內段含有“死亡結構域(Death domain,DD)”[2,3]。TRAIL 與 DR4/DR5 結合后,能夠激活死亡受體通路及線粒體通路誘導多種腫瘤細胞凋亡[4-6],在臨床試驗中也顯示出較好的抗腫瘤活性,但是肝細胞損害及一些腫瘤細胞對TRAIL誘導凋亡的抵抗等原因,一定程度上阻礙了 TRAIL的應用[7,8]。
近年研究發現,一些抗DR4/DR5的激活型抗體能夠模擬TRAIL的作用,有效誘導腫瘤細胞凋亡,并且無 TRAIL 的肝細胞毒性[9,10]。本實驗室成功研制一株抗人DR5激活型抗體-mDRA-6,前期實驗表明,mDRA-6與細胞膜DR5結合,能夠有效激活caspase-8等死亡受體途徑信號分子,誘導多種腫瘤細胞凋亡[11],但mDRA-6誘導腫瘤細胞凋亡過程中,是否有線粒體通路信號分子的激活尚不清楚,為了更全面研究mDRA-6誘導腫瘤細胞凋亡機制,本實驗在白血病細胞系Jurkat和U937細胞上,探討mDRA-6對腫瘤細胞線粒體通路信號分子的激活改變,為其臨床抗腫瘤應用提供理論基礎。
1 材料與方法
1.1材料 鼠抗人DR5單克隆抗體mDRA-6由本實驗室制備;Jurkat細胞株由美國賓夕法尼亞大學醫學院陳有海教授饋贈,U937細胞購于中國醫學科學院上海細胞生物研究所;RPMI Medium 1640購于Gibco公司;胎牛血清購于天津TBD公司;四氮唑藍(MTT)、DMSO購自Sigma公司;FITC-AnnexinV/PI試劑盒購自BD公司;Western細胞裂解液購自Beyotime公司;caspase-9抑制劑 Z-LEHD-FMK及caspase-3抑制劑Z-DEVD-FMK購于RD公司;山羊抗人 caspase-9抗體、兔抗人 caspase-3抗體購自R&D公司;兔抗人Bcl-xl抗體購自Cell Signaling公司;兔抗人Bcl-2抗體、鼠抗人Bax、actin抗體購自Beyotime公司;山羊抗人Cytc單克隆抗體購自Santacruz公司;辣根過氧化酶標記羊抗鼠-HRP、羊抗兔-HRP及兔抗山羊IgG(H+L)購自北京中杉金橋公司;ECL顯影試劑盒購自Amershem Pharmacia公司。PVDF膜購自Milipore公司;流式細胞儀(FACSCalibur)為BD公司產品;垂直電泳儀及電轉裝置為Thermo EC公司產品;凝膠圖像分析儀(Alphalmager2200)為Alpha Inntech公司產品。
1.2方法
1.2.1mDRA-6對Jurkat及U937細胞生長增殖抑制作用 分別收集對數生長期的Jurkat和U937細胞,加入96孔培養板,細胞密度為3×104/孔,分別以不同濃度 (10、5、2.5、1.25、0.64、0.32、0.16 mg/L)的mDRA-6培養細胞10小時,或用終濃度為10 mg/L 的 mDRA-6 分別處理細胞 1、2、4、6、8、10小時。更換新培養液,加入MTT(5 mg/L)20 μl/孔,繼續培養4小時。離心,吸棄上清,每孔加入150 μl DMSO,振蕩混勻,全自動酶標儀測OD570值。每一實驗濃度設3個復孔,實驗重復3次。按以下公式計算細胞生長抑制率,細胞生長抑制率=(1-實驗組OD值/對照組OD值) ×100%。
1.2.2DNA ladder檢測mDRA-6對細胞的凋亡作用 調整Jurkat及U937細胞濃度為1×107/瓶,加入終濃度為10 mg/L的 mDRA-6,置于37℃ 5%CO2培養箱內孵育2小時,陰性對照為相同濃度的小鼠IgG1。收集細胞,800 r/min離心5分鐘,棄上清,PBS洗細胞2次。提取細胞DNA,1%瓊脂糖凝膠電泳,紫外下觀察并拍照。
1.2.3Western blot檢測凋亡信號分子變化 取對數生長期的Jurkat及U937細胞,加終濃度為10 mg/L的mDRA-6,分別培養0分鐘、5分鐘、15分鐘、30分鐘、1小時、2小時、3小時、4小時后收集細胞,用預冷PBS洗細胞2次,加細胞裂解液(20 mmol/L HEPES pH7.5,0.35 mmol/L NaCl,20%甘油,1%NP-40,1 mmol/L MgCl2·6H2O,0.5 mmol/L EDTA,0.1 mmol/L EGTA,1 mmol/L PMSF)冰上裂解30分鐘,10 000 r/min離心15分鐘,收集上清,用BCA法檢測上清液蛋白濃度,取細胞總蛋白60 μg,進行SDSPAGE電泳,并轉移PVDF膜。用含有5%脫脂奶粉的TBST室溫封閉PVDF膜1小時,洗滌4次,分別加入抗人 caspase-9、caspase-3、bax、bcl-2、bcl-xl及Cyt c抗體,室溫孵育2小時,洗滌4次。將膜封閉于HRP標記的二抗稀釋液中,室溫孵育1小時,TBST洗膜后,ECL反應液中反應1分鐘,暗室顯影。
1.2.4Caspase-9、3抑制劑對mDRA-6抑制細胞生長增殖的影響 收集Jurkat和U937細胞,接種于96孔板中,細胞密度為3×104細胞/孔,分別加入終濃度為15 μmol/L的caspase-9、3抑制劑,對照組加入同體積的 RPMI1640完全培養液。置37℃ 5%CO2培養箱內孵育1小時后,加入終濃度為10 mg/L的 mDRA-6,終體積為 200 μl/孔,置于37℃ 5%CO2培養箱內孵育8小時。每一濃度設3個復孔,實驗重復3次。上述MTT法檢測細胞生長抑制率。
1.2.5Caspase-9抑制劑對mDRA-6誘導細胞凋亡的影響 分別制備Jurkat和U937細胞懸液,加入24孔培養板,細胞數為3×105/孔,分別加入終濃度為15 μmol/L的 caspase-9抑制劑,終體積為1 ml/孔,對照組加入同體積的RPMI1640完全培養液。37℃孵育1小時后,加入終濃度為10 mg/L的mDRA-6,置37℃ 5%CO2培養箱內孵育細胞2小時。收集細胞,用結合液洗細胞2次,并將細胞懸浮于100 μl標記液中,加 FITC-AnnexinV/PI染液各5 μl,混勻后避光冰浴15分鐘,流式細胞術檢測細胞凋亡率。Cellquest軟件分析細胞凋亡率。
1.3統計學分析 實驗數據用±s表示,組間t檢驗進行統計學分析,P<0.05認為有統計學差異。
2 結果
2.1mDRA-6對Jurkat及U937細胞生長增殖影響
不同劑量(10、5、2.5、1.25、0.64、0.32、0.16 mg/L)的mDRA-6孵育Jurkat及U937細胞10小時,兩種細胞均呈現濃度依賴性地生長抑制;10 mg/L的mDRA-6也呈時間依賴性地抑制Jurkat及 U937細胞的生長增殖,10 mg/L的 mDRA-6孵育 Jurkat細胞6、8和10小時,細胞增殖抑制率分別達59.38%、72.56%和76.28%;10 mg/L的mDRA-6孵育U937細胞6、8和10小時,細胞增殖抑制率分別達38.67%、47.54%和50.59%,見圖1。
2.2DNA ladder檢測mDRA-6對Jurkat及U937細胞的凋亡作用 10 mg/L的mDRA-6作用Jurkat及U937細胞2小時,提取細胞DNA,進行1%瓊脂糖凝膠電泳,結果顯示,mDRA-6處理的 Jurkat及U937細胞均呈現凋亡細胞所特有的DNA ladder條帶,而對照組細胞無梯形圖譜產生,結果見圖2。
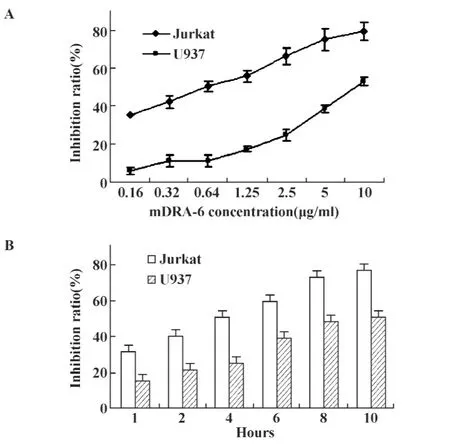
圖1 mDRA-6對Jurkat和U937細胞的生長抑制作用Fig.1 Inhibition of mDRA-6 on Jurkat and U937 cells
2.3Western blot檢測細胞凋亡分子 mDRA-6作用后,Jurkat和U937細胞內的凋亡分子發生變化。隨著mDRA-6作用時間延長,Jurkat和U937細胞內的促凋亡分子bax激活增多,Cyt c釋放增多,而同時細胞抗凋亡分子bcl-2及bcl-xl隨mDRA-6作用時間延長不斷減少。mDRA-6作用后,Jurkat和U937細胞內34 kD的caspase-9裂解片段明顯增多;Jurkat和U937細胞caspase-3也顯示明顯的激活表現,Jurkat細胞 caspase-3激活尤為顯著,10 mg/L的mDRA-6作用Jurkat細胞5分鐘,Jurkat細胞caspase-3即有17 kD的裂解條帶顯示,且隨著mDRA-6作用時間延長,12 kD的裂解片段明顯呈現。結果見圖3。
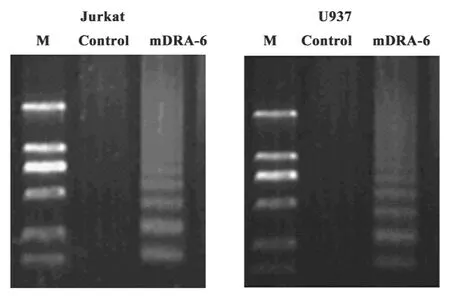
圖2 DNA梯形條帶分析Fig.2 DNA ladder analysis
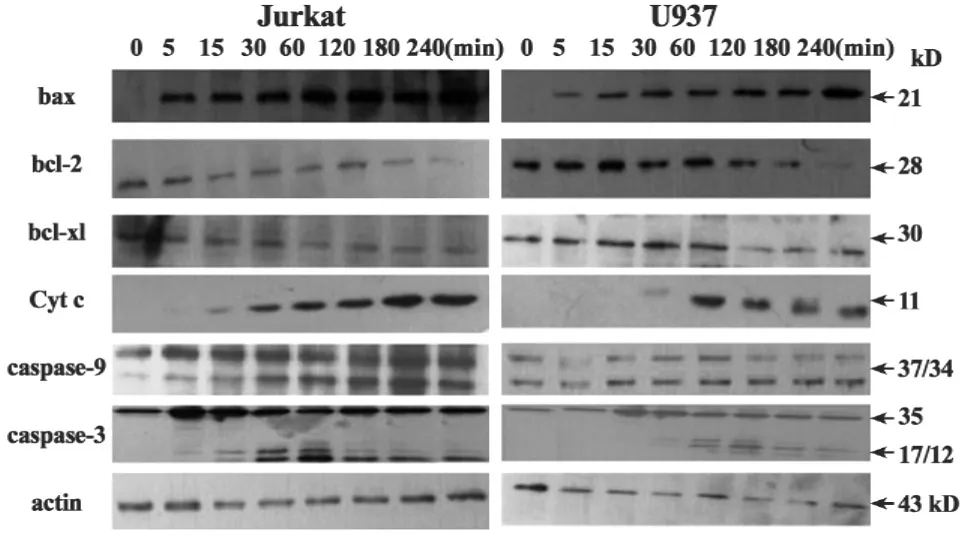
圖3 mDRA-6激活Jurkat及U937細胞凋亡分子Fig.3 Appototic molecule activation in Jurkat and U937 cells treated with mDRA-6
2.4Caspases-9、3抑制劑對mDRA-6抑制細胞生長增殖的影響 Caspase-9、3抑制劑能夠不同程度降低mDRA-6對Jurkat和U937細胞的生長抑制作用。預先使用caspase-9、3抑制劑孵育細胞1小時,mDRA-6所致Jurkat細胞生長抑制率分別降低了24.36%(t=5.44,P<0.01)和31.18%(t=7.97,P<0.01);mDRA-6所致U937細胞生長抑制率分別降低了20.82%(t=4.29,P<0.01)和50.20%(t=12.98,P<0.01)。Caspase-9、3抑制劑單獨使用,對Jurkat和U937細胞生長增殖無明顯影響,結果見圖4。
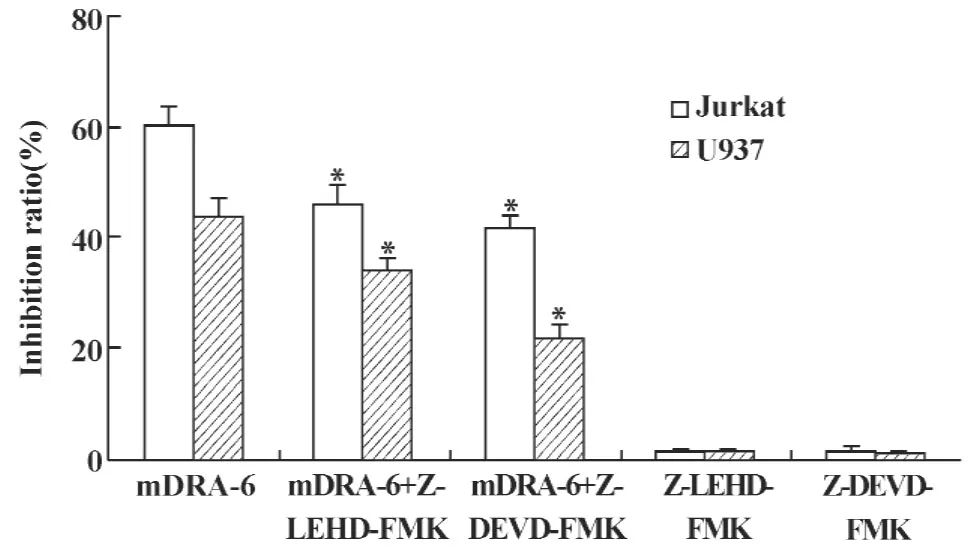
圖4 Caspases抑制劑對mDRA-6抑制Jurkat及U937細胞生長的影響Fig.4 The effects of caspases inhibitors on cell inhibition ratio of Jurkat and U937 traeted with mDRA-6
2.5Caspase-9抑制劑對mDRA-6誘導細胞凋亡的影響 AnnexinⅤ/PI雙染,流式細胞術檢測結果表明,10 mg/L的mDRA-6作用Jurkat和U937細胞2小時,細胞凋亡率分別為66.64%和38.50%。預先用15 μmol/L的 caspase-9抑制劑孵育 Jurkat或U937細胞1小時,mDRA-6誘導的細胞凋亡率分別降低了32.89%和23.97%。15 μmol/L的caspase-9抑制劑對Jurkat和U937細胞無明顯凋亡作用,結果見圖5。
3 討論
TRAIL與細胞膜上的相應受體(TRAIL-R1/DR4,TRAIL-R2/DR5)結合,啟動胞外凋亡通路(死亡受體通路)和胞內凋亡通路(線粒體通路),誘導多種腫瘤細胞凋亡。TRAIL與含有死亡結構域(Death domain,DD)的細胞膜DR4或DR5結合,通過銜接蛋白 FADD(Fas associated death domain,FADD),激活caspase-8/caspase-10,啟動外源性凋亡途徑導致細胞凋亡[4,5]。
TRAIL胞內凋亡通路是線粒體依賴性的凋亡通路,受Bcl-2蛋白家族中促凋亡因子和抗凋亡因子相互調控,二者的平衡對于調節線粒體功能有重要價值。胞內凋亡通路主要的促凋亡成員Bax和Bak通過破壞線粒體膜的完整性發揮作用,bax是bcl-2家族的重要的促凋亡蛋白,在正常細胞中主要分布在胞質,在TRAIL等多種凋亡刺激因子的作用下,可以發生轉位與線粒體相互作用,損傷線粒體造成線粒體膜電位變化,導致線粒體內某些促凋亡分子,如Cyt c等外流,進而活化caspase-9,激活效應caspases分子,啟動細胞凋亡過程。抗凋亡成員bcl-2和bcl-xl在線粒體外膜發揮作用,bcl-2和bcl-xl可以和bax等結合形成異構二聚體,阻止bax對線粒體的損傷,以維持線粒體膜的完整性,抑制細胞凋亡。bax表達水平增加可拮抗bcl-2的作用,并促進細胞凋亡[12]。
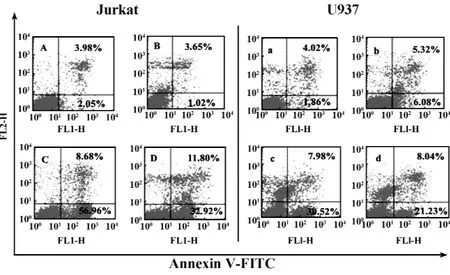
圖5 Caspases-9抑制劑抑制mDRA-6誘導的Jurkat/U937細胞凋亡Fig.5 Caspases-9 inhibitor inhibit the apoptosis of Jurkat and U937 cells treated with mDRA-6
研究表明激活型抗DR5抗體能夠通過死亡受體通路,激活 caspase級聯反應,啟動細胞凋亡[13,14]。本實驗中我們發現,激活型抗 DR5 抗體mDRA-6呈濃度、時間依賴性地誘導Jurkat和U937細胞凋亡,同時細胞內線粒體通路主要凋亡分子bax表達隨mDRA-6作用時間而增多,而主要抗凋亡分子bcl-2及bcl-xl隨mDRA-6作用時間而減少,提示線粒體通路可能參與了mDRA-6誘導的細胞凋亡,為了進一步明確線粒體通路的激活,我們應用Western blot方法,檢測線粒體釋放的凋亡因子Cyt c變化,結果發現mDRA-6作用Jurkat細胞15分鐘,即可檢測到Cyt c釋放,并隨mDRA-6作用時間延長而增多;mDRA-6作用U937細胞也顯示有Cyt c釋放增多。線粒體釋放的Cyt c等凋亡因子,能夠使細胞procaspase-9自身催化形成有活性的caspase-9,隨后激活下游caspase-3等效應caspases分子,導致細胞凋亡。caspase-9激活是線粒體通路激活的重要環節,本實驗中 Western blot檢測發現,mDRA-6作用Jurkat及U937細胞后,細胞caspase-9、caspase-3均有激活片段產生,且預先應用caspase-9抑制劑能夠有效抑制mDRA-6所致的Jurkat及U937細胞凋亡率,進一步表明線粒體通路激活是mDRA-6誘導Jurkat及U937細胞凋亡的重要機制之一。
抗人DR5的激活型單克隆抗體具有TRAIL的誘導腫瘤細胞凋亡作用,又克服了TRAIL的受體多樣性及肝細胞毒性作用,可能較TRAIL有更好的抗腫瘤應用前景。本實驗中的抗DR5單克隆抗體mDRA-6,能夠有效誘導白血病Jurka及U937細胞凋亡,細胞線粒體通路激活是其誘導細胞凋亡的機制之一。對mDRA-6誘導腫瘤細胞凋亡機制的進一步研究,可能對提高mDRA-6抗腫瘤臨床應用提供有用的理論基礎。
1 Sarah S,Alexandre M,Olivier M.Regulating TRAIL receptor-induced cell death at the membrane:a deadly discussion[J].Recent Pat Anticancer Drug Discov,2011;6(3):311-323.
2 Duiker E W,Mom C H,Jong S et al.The clinical trail of TRAIL[J].Eur J Cancer,2006;42:2233-2240.
3 Mérino D,Lalaoui N,Morizot A et al.Differential inhibition of TRAIL-mediated DR5-DISC formation by decoy receptors 1 and 2[J].Mol Cell Biol,2006;26:7046-7055.
4 Gaelle P,Valerie T,Stephane T et al.TRAIL receptor signaling and therapeutic option in bone tumors:the trap of the bone microenvironment[J].Am J Cancer Res,2012;2(1):45-64.
5 Pennarun B,Meijer A,de Vries E G et al.Playing the DISC:turning on TRAIL death receptor-mediated apoptosis in cancer[J].Biochim Biophys Acta,2010;1805(2):123-140.
6 Walczak H,Haas T L.Biochemical analysis of the native TRAIL death-inducing signaling complex[J].Methods Mol Biol,2008;414:221-239.
7 Koschny R,Walczak H,Ganten T M.The promise of TRAIL-potential and risks of a novel anticancer therapy[J].J Mol Med,2007;85(9):923-935.
8 Nadzeva G K,Denis L,Isabelle M et al.Targeted ovarian cancer treatment:the TRAILs of resistance[J].Am J Cancer Res,2012;2(1):75-92.
9 Greco F A,Bonomi P,Craw ford J et al.Phase 2 study of mapatumumab,a fully human agonistic monoclonal ant ibody which target sand activates the TRAIL receptor1 inpatients with advanced nonsmall cell[J].Lung Cancer,2008;61(1):82-90.
10 Tolcher A W,Mita M,Mer opol N J et al.PhaseⅠpharmacokinetic and biologic correlative study of mapatumumab,a fully human monoclonal antibody with agonist activity to tumor necrosis factor-related apoptosis-inducing ligand receptor[J].J Clin Oncol,2007;25(11):1390-1395.
11 杜耀武,劉廣超,王 靖et al.抗人DR5功能性抗體mDRA-6誘導人白血病 Jurkat細胞凋亡的機制[J].癌癥,2009;28(2):136-142.
12 Brooks C,Dong Z.Regulation of mitochondrial morphological dynamics during apoptosis by Bcl-2 family proteins:a key in Bak[J].Cell Cycle,2007;6(24) :3043-3047.
13 Fulda S.Caspase-8 in cancer biology and therapy[J].Cancer Lett,2009;281:128-133.
14 Jing Q,Zhu H,Liang B.Apotosis-inducing antibody D-6 alone or with cisplatin on A2780 ovarian cancer cells[J].Mol Med Reptor,2012;6(2):316-320.
