云南地區恒河猴線粒體DNA控制區遺傳多態性研究
2012-01-30 08:02:04禹文海楊鳳梅魯帥堯王俊斌陳麗雄李艷艷和占龍
中國比較醫學雜志 2012年4期
禹文海,黃 芬,楊鳳梅,魯帥堯,趙 遠,王俊斌,陳麗雄,李艷艷,和占龍
(1.中國醫學科學院北京協和醫學院 醫學生物學研究所,云南省重大傳染病疫苗研發重點實驗室,昆明 650118;2.昆明理工大學生命科學與技術學院,昆明 650224)
恒河猴 (Macaca mulatta)隸屬于靈長目(Primates)猴科(Cercopithecidae)獼猴屬(Macaca),為國家Ⅱ級保護動物和重要的實驗靈長類動物,是非人靈長類中分布最廣且數量最多的一個類群,廣泛分布于中國、緬甸、尼泊爾、印度北部、孟加拉國、巴基斯坦等地。在我國主要分布于南方諸省(區),以廣東、廣西、云南、貴州等地分布較多,福建、安徽、江西、湖南、湖北、四川次之,海南、陜西、山西、河南、河北、青海、西藏等局部地點也有分布。由于恒河猴在形態、生殖生理特性和生化代謝等方面與人類非常相似,而且遺傳物質與人類具有較高的同源性,是進行醫學和生物學等領域研究較為理想的實驗動物[1]。
線粒體 DNA(mitochondrial DNA,m tDNA)呈母系方式遺傳,無重組,進化速率快。其中,mtDNA控制區的進化速度是線粒體DNA其它區域的3~5倍,而且不編碼任何基因,在生物遺傳進化及種群遺傳變異程度的研究中應用廣泛[2-6]。本研究以分布于云南不同地區的96份恒河猴樣品作為研究對象,采用DNA測序技術測定m tDNA控制區全序列,初步確定云南地區恒河猴群體遺傳多態性,為今后開展恒河猴遺傳資源保護和合理開發利用提供理論依據。
1 材料和方法
1.1 實驗材料
96份恒河猴全血樣品來自中國醫學科學院醫學生物學研究所全國醫學靈長類研究中心實驗動物部[實驗動物生產許可證:(滇)SCXK2010-0006]。其中,23份為本中心自繁恒河猴(昆明市),其余73份恒河猴為從野外捕獲的原種群,來源地清晰。在73份野外恒河猴中,景東縣21份、鎮沅縣21份 、寧蒗縣24份、保山市2份、牟定縣2份、鄧川鎮1份、鎮康縣1份、維西縣1份。樣品來源地理位置見圖1。
1.2 試劑和儀器
Axygen DNA提取試劑盒(杭州愛思進生物技術有限公司),2×PCR Master Mix、ddH2O、DNA marker DL2000和6×電泳上樣緩沖液(北京天根生化科技有限公司),臺式離心機(Legend Micro 21R),梯度PCR儀(Bio-Rad C1000),DYY-2C型電泳儀電源和DYCP-31DN型電泳儀(北京市六一儀器廠),UVP凝膠成像系統(美國基因有限公司)。
參考相關文獻設計引物[7,8],由上海生工生物工程有限公司合成。上游引物F:CCTTACCTGAA TTGGAAGCGAACC;下游引物R:GGCCAGGACC AAGCCTATTT。
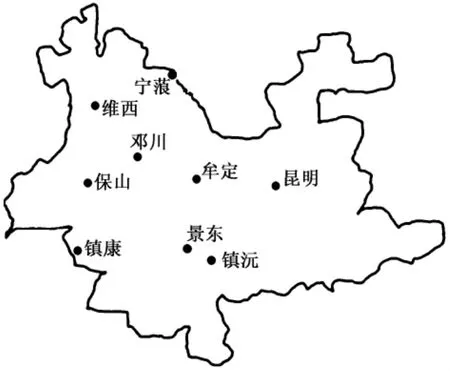
圖1 云南地區恒河猴樣品來源地理位置Fig.1 The sampling sites of the Macaca mulatta in Yunnan province
1.3 方法
1.3.1 基因組DNA的提取:靜脈抽取全血2m L,EDTA抗凝。采用 Axygen DNA提取試劑盒提取基因組DNA,詳細操作步驟見說明書。-20℃保存備用。
1.3.2 PCR擴增:反應體積為25μL,上游引物F和下游引物R各(20μmol/L)0.5μL,模板DNA 2μL,2×PCR Master Mix 12.5μL,ddH2O 9.5μL。反應條件為95℃預變性6m in;然后94℃變性1min,66℃退火 45s,72℃延伸 1m in,40個循環;最后72℃延伸10min。
1.3.3 瓊脂糖凝膠電泳及 PCR產物直接測序:用1.5%的瓊脂糖凝膠電泳檢測PCR產物,參照DNA marker DL2000,對1450bp左右的PCR產物進行純化測序,測序由上海杰李生物技術有限公司完成。
1.3.4 數據處理與結果分析:每個個體測得的正反鏈序列用DNAStar軟件進行分析拼接,Clustalx 1.8軟件進行序列比對并輔以人工校對,Mgea 4.0軟件分析序列的堿基組成、轉換和顛換,用 DNA SP (Version 4.10.8)軟件分析變異位點數(variation sites)、簡約信息位點數 (parsimony informative sites)、單倍型數(haplotypes,h)、單倍型多樣度(haplotype diversity,Hd)、核苷酸多樣度(nucleotide diversity,Pi)、平均核苷酸差異(average No.of nucleotide differences,K)等。以食蟹猴(GenBank登錄號:FJ906803)為外群,用鄰接法(neighbor-joining,NJ)和最小進化法(minimum-evolution,ME)構建分子系統發生樹。
2 結果
2.1 PCR產物電泳檢測結果
PCR產物經1.5%瓊脂糖凝膠電泳后得到片段大小約為1450bp左右的清晰條帶(圖2),與預期結果一致。
圖2 恒河猴線粒體DNA控制區及其兩側部分序列的PCR產物注:M.分子量標準DL2000;1~9.樣品編號Fig.2 PCR products of mtDNA control region and its flanking sequences in Macaca mulatta.Note:M.DNA marker DL2000;1~9.number of samples
2.2 單倍型分布
PCR擴增產物測序后,利用BLAST和GenBank中的恒河猴線粒體序列進行比較,同源性達到91.5%以上,說明該序列即為目的基因。經DNA SP軟件分析,所有96個序列定義了46個不同的單倍型(H1-H46),各單倍型在不同種群中的分布情況見表1。有7個單倍型(H1、H5、H13、H25、H26、H27、H28)為部分群體間共享的單倍型,單倍型H1為鎮沅、景東、維西三個群體共享,單倍型H5為鎮沅和昆明自繁群體共享,單倍型H13為鎮沅和景東群體共享,單倍型H25為保山和昆明自繁群體共享,單倍型H26為鎮沅和景東群體共享,單倍型H27、H28為寧蒗和昆明自繁群體共享,其余均為各群體特有單倍型。鎮沅群體以H1(6/21)和H13(4/21)為主要單倍型,景東群體以 H26(5/21)、H13(3/21)和H29(4/21)為主要單倍型,寧蒗群體以H11(3/24)、H18(7/24)、H27(4/24)和H28(4/24)為主要單倍型,昆明自繁群體以H22(5/22)和H37(3/22)為主要單倍型。
以本研究測定的單倍型H1為參考序列,在96份樣品中,共檢測出了149個多態性位點(變異位點分布情況見圖3),約占13.70%。其中單一多態位點42個,約占3.86%;簡約信息位點107個,約占9.84%。兩核苷酸間的單一多態位點為41個,約占3.77%;兩核苷酸間的簡約信息位點為105個,約占9.66%。三核苷酸間的單一多態位點為1個,約占0.09%;三核苷酸間的簡約信息位點為2個,約占0.18%。核苷酸變異包括轉換、顛換、插入和缺失,所有核苷酸多態位點主要以轉換為主。

表1 46個單倍型在云南地區不同恒河猴群體中的分布Tab.1 Distribution of 46haplotypes in different Macaca mulatta populations in Yunnan province
2.3 云南地區恒河猴遺傳多樣性
經DNA SP軟件分析,云南地區96份恒河猴樣品線粒體DNA控制區核苷酸序列共定義了46種單倍型,單倍型多樣度(Hd)為0.955±0.015,核苷酸多樣度(Pi)為0.020。各群體內單倍型多樣度和核苷酸多樣度等信息見表2。從該表中可以看出,云南地區不同恒河猴群體單倍型多樣度在0.866± 0.042和 1.000±0.500之間,核苷酸多樣度在0.003和0.042之間,平均核苷酸差異在3.000和60.000之間。變異位點數最多的是昆明自繁恒河猴群體(114個),其次是景東恒河猴群體(75個)。簡約信息位點數最多是昆明自繁群體(78個),其次是寧蒗恒河猴群體(37個)。云南地區不同恒河猴群體Tajima's D中性檢驗均不顯著(P >0.10)。
2.4 單倍型的系統進化關系
以食蟹猴為外群,借助 Mega軟件采用 NJ和ME法構建了云南不同地理種群46個單倍型的系統發生樹(圖4),各分支上的數值為1000次自舉(bootstrap)分析得到的支持率。從圖4中可以看出NJ樹和ME樹具有基本相同的拓撲結構,兩者均表明所分析的序列分為兩大分支,食蟹猴單獨聚為一支,云南地區獼猴聚為另一支。所分析的云南地區獼猴分為兩個平行的姐妹分支,來自不同地理位置的獼猴種群單倍型大多按照其來源地分別相對聚在一起,表現出了與地理位置一定的對應關系。其中單倍型 H36(牟定)、H30(牟定)、H42(昆明自繁)、H45(昆明自繁)、H33(景東)、H23(保山)和H41(昆明自繁)聚為第一分支,其他單倍型聚為第二分支。在第一分支中,除了景東來源的單倍型H33與其他單倍型沒有直接血統關系外,保山來源的獼猴與牟定來源的獼猴相對聚在一起(根據個體檔案記錄可知,單倍型H42、H45具有牟定來源獼猴的血統,H41具有保山來源獼猴的血統)。第二分支中,來源于鎮康的獼猴(H32)單獨聚為一支,其它單倍型聚為一支。其中,來源與鎮沅和景東獼猴相對聚在一起;寧蒗來源獼猴除了單倍型H18和H28外,其它單倍型均聚為一個亞分支(H11、H14、H15、H17、H27、H40、H44)。

表2 云南地區恒河猴群體內線粒體DNA控制區核苷酸序列多態性Tab.2 Nucleotide polymorphism of mtDNA control region within the Macaca mulatta population in Yunnan province

圖4 基于線粒體DNA控制區構建的NJ(A)和ME(B)系統樹Fig.4 Phylogenetic trees constructed using NJ(A)and ME(B)methods based on the complete sequences of mtDNA control region
3 討論
恒河猴在我國《國家重點保護野生動物名錄》中被列為國家二級保護動物,在《中國瀕危動物紅皮書獸類》中被列為易危種。而遺傳多樣性的高低是評判物種是否能夠長期生存的依據,遺傳多樣性的喪失對物種生存帶來直接的不利影響,可以使物種更易滅絕[9]。因此,對恒河猴遺傳資源進行保護及合理開發利用顯得非常的重要。許多學者從蛋白質水平對中國恒河猴進行了分析,結果表明中國恒河猴在蛋白質水平上的遺傳標記數量有限、變異不夠豐富、多態性有局限。目前認為DNA分析是最有效的遺傳多樣性分析方法,中國恒河猴DNA分子水平的遺傳多態性主要見于對主要組織相容性復合體(MHC)、微衛星座位及線粒體 DNA的研究[10-14],研究結果表明中國恒河猴表現出了豐富的遺傳多樣性。
本研究從分子水平研究了云南地區恒河猴遺傳多樣性,在96個恒河猴個體中,共檢測出了149個多態性位點,定義了46種單倍型,其中僅有7個單倍型為部分群體間共享,其余均為各群體特有單倍型。各群體的核苷酸多樣度(Pi)均較高,從0.003(牟定)至0.055(保山),說明云南地區恒河猴遺傳多態性較高,遺傳資源較豐富。此外,云南地區不同恒河猴群體Tajima's D中性檢驗均不顯著(P>0.10),這些群體均符合中性突變,說明云南地區恒河猴在過去沒有出現群體擴張,群體大小較穩定。
[1]郅琦,陳永昌,陳學進.獼猴在生物醫學研究中的應用[J].上海實驗動物科學,2003,23(1):59-62.
[2]Nishibori M,Shimogiri T,Hayashi T,et al.Molecular evidence for hybridization of species in the genus Gallus except for Gallus varius[J].Anim Genet,2005,36(5):367-375.
[3]Wenink PW,Baker AJ,Tilanus MG.Mitochondrial controlregion sequences in two shorebird species,the turnstone and the dunlin,and their utility in population genetic studies[J].Mol Biol Evol,1994,11(1):22-31.
[4]潘登,符云新,張亞平.基于線粒體控制區的滇金絲猴群體遺傳學研究[J].遺傳,2006,28(4):432-436.
[5]何麗,張于光,李迪強,等.川金絲猴mtDNA D-loop序列遺傳多樣性分析[J].動物學雜志,2010,45(1):70-76.
[6]徐懷亮,李地艷,程安春,等.四川地區獼猴線粒體DNA控制區遺傳多樣性及其種群遺傳結構[J].獸類學報,2010,30(3):247-255.
[7]Blancher A,Bonhomme M,Crouau-Roy B,et al.Mitochondrial DNA sequence phylogeny of 4 populations of the widely distributed cynomolgus macaque(Macaca fascicularis fascicularis)[J].J Hered,2008,99(3):254-264.
[8]董高華,徐懷亮,帥素容.藏酋猴線粒體基因組序列測定與進化分析[D].碩士學位論文,2008,四川農業大學.
[9]David P.Heterozygosity-fitness correlations:new perspectives on old problems[J].Heredity(Edinb),1998,80(pt5):531-537.
[10]閆熙,李萬波.利用微衛星遺傳標記對恒河猴進行遺傳同質性分群的探索[J].中國比較醫學雜志,2009,19(11):11-18.
[11]張卉,李進華,趙健元,等.皖南山區獼猴種群的形態特征與微衛星遺傳多樣性初步分析[J].實驗動物與比較醫學,2008,28(4):225-229.
[12]萬玉玲,季芳,饒軍華,等.中國不同地域恒河猴MHC-I型部分等位基因的調查[J].動物學雜志,2007,42(2):1-5.
[13]李瑞生,趙爽,高蓉,等.恒河猴與食蟹猴微衛星DNA多態性的比較分析[J].中國實驗動物學報,2009,17(1):57 -60.
[14]徐懷亮,汪宴廷,程安春,等.獼猴MHC-DPB1基因外顯子2多態性研究[J].遺傳,2010,32(6):588-598.
