薩能奶山羊乳腺組織5-HTR4基因的克隆序列分析與原核表達
2012-02-24 01:10:04江青東
中國獸醫雜志 2012年7期
關鍵詞:分析
江青東
(鄭州牧業高等專科學校動物醫學系,河南 鄭州 450011)
5-羥色胺(5-HT)是一種吲哚衍生物,分子式C10H12N2O。5-HT能與酸作用生成結晶鹽,其熔點167℃~168℃。5-HT是調節神經活動的一種重要物質[1-4]。美國辛辛那提大學醫學院納爾遜·霍斯曼等人研究發現,5-羥色胺能降低奶牛的產奶量。只要抑制奶牛乳腺中的5-HT含量,就可以將產奶量提高15%,這些作用可被5-HTR拮抗劑二甲麥角新 堿 阻 斷,提 示是經過 5-HTR 產 生 的[5-7]。目前,針對哺乳動物5-HTR 基因在乳腺組織中的分布尚未報道。
本試驗通過牛和鼠在其他組織中的5-HTR4基因序列設計同源性引物采用RT-PCR方法克隆出奶山羊乳腺組織中5-HTR4的基因組全序列,這將對下一步研究特異性針對乳腺中的5-HTR4,通過基因敲除技術,是一種綠色環保藥物,來提高哺乳動物的產奶量和養殖戶的經濟效益奠定理論和技術基礎。
1 材料與方法
1.1 樣品采集 取泌乳期奶山羊(購自陜西薩能奶山羊養殖基地)乳腺組織,液氮速凍后保存于-80℃。
1.2 菌株和質粒 p MD19-T 載體、E.coliJM109感受態細胞、質粒p GEX-4T-1,均購自上海生工生物工程技術服務公司,E.coliBL21(Promega公司)。
1.3 試劑 AMV反轉錄酶、TaqDNA聚合酶、動物 組 織 RNAout、RNase-Inhibitor、Oligo(d T)、Ta KaRa Agarose Gel、DNA Purification Kit Ver.2.0、DNA Marker、TaqDNA聚合酶、TaKaRa保存待測。BamHⅠ、XhoⅠ和IPTG,均購自Ta KaRa寶生物工程(大連)有限公司,T4 DNA連接酶(Promega公司);蛋白質標準Marker(Sigma公司)。
1.4 克隆與序列分析
1.4.1 引物設計 根據GenBank公布的牛和鼠的5-HTR4基因的cDNA序列,利用Primer 5.0引物設計軟件和BLAST鑒定設計克隆引物。上游:5′-GTGGAGAAGGTSGTGCTGCTCACG -3′,下 游:5′-GGTTGTGGTTGAACARGGGACAGTCTG-
3′,預期片段1 293 bp,引物由北京三博遠志生物科學技術有限公司合成。
1.4.2 RNA的提取 按照動物組織RNAout試劑盒的說明進行提取乳腺組織總RNA,用0.8%瓊脂糖凝膠電泳(130 V,15 min),紫外燈下觀察。紫外分光光度計測定OD260和OD280,檢測RNA的純度及含量(圖1)。
圖1 乳腺組織中總RNA電泳結果
1.4.3 RT-PCR擴增體系 RT體系:模板 RNA 2 μL,5×Buffer 4μL,d NTP Mix 4μL,RNase Inhibitor 0.5μL,Oligo(d T)1μL,AMV 0.5μL,DEPCH2O 6μL,總體積20μL。混勻室溫放置10 min,42℃1 h,置-20℃冰箱備用。PCR擴增體系:10×Buffer(Mg2+)5μL,d NTP Mix 6μL,25 mmol/L MgCl26μL,rTaq0.5μL,上、下游引物各1μL,cDNA 4μL,dd H2O 14μL,總體積50μL。PCR反應程序:95℃變性5 min;95℃10 s,52℃20 s,72℃30 s,共35個循環;最后72℃延伸10 min。取5μL PCR產物用1.2%瓊脂糖凝膠(100 V、30 min)電泳。凝膠照像并檢測PCR產物積分光密度值,用各基因與β-actin的比值相對定量轉錄水平。
1.4.4 PCR產物回收 產物回收根據 UNIQ-10膠回收試劑盒說明書進行。
1.4.5 目的基因的克隆與測序 連接產物轉化,堿裂解法提取重組質粒DNA,通過酶切和PCR鑒定,1%瓊脂糖凝膠電泳鑒定結果;陽性菌液送上海生工生物工程技術服務公司測序。
1.4.6 序列分析 通過 Blast、Smart及 BioXM2.6和DNAStar軟件對序列進行分析。
1.5 原核表達載體p GEX-5-HTR4的構建
1.5.1 引物 根據克隆的序列設計一對表達引物,上游:5′-CGCCCATCGTCGCATCGCATCG-3′,下游:5′-CCGCGTGAGATCACGCTCTCATG-3′,并在引物中引入BamHⅠ/XhoⅠ酶切位點(劃線)。
1.5.2 PCR 擴增體系 dd H2O 15.3μL,10×Buffer(Mg2+)2.5μL,d NTP Mix 3μL,25 mmol/L MgCl21.8μL,P1、P2 各 0.5μL,克隆載體p MD19-T-5-HTR4 1μL,rTaq0.4μL,總體積25 μL。反應程序:95℃預變性5 min;95℃變性30 s,59.5℃退火30 s,72℃延伸30 s,共30個循環;最后72℃延伸10 min。PCR產物純化按照純化試劑盒說明書進行。
1.5.3 質粒pGEX-4T-1和純化后的 PCR產物進行BamHⅠ和XhoⅠ雙酶切 純化回收產物用T4 DNA連接酶進行連接,22℃3 h,然后轉化E.coliBL21(DE3)感受態細胞,在含有氨芐青霉素的LB固體培養基中37℃培養過夜。提取重組質粒,并進行PCR和雙酶切鑒定,測序由北京三博遠志生物科學技術有限公司完成。并將重組質粒命名為p GEX-5-HTR4。
1.6 重組質粒的誘導表達與鑒定 將含重組質粒(pGEX-5-HTR1)的E.coli單 菌 落 和 含 空 載 體(pGEX-4T-1)E.coli單菌落分別接種于含有氨芐青霉素的2×YT液體培養基,于37℃搖震培養過夜。然后將培養物按1∶100的比例接種于新鮮的含有氨芐青霉素的2×YT液體培養基,37℃搖震培養至A600=0.5~0.6時,加入IPTG 35℃誘導9 h,每2 h取出1 m L培養物。離心收集菌體,用20μL PBS重懸菌體,并加入20μL 2×SDS-PAGE電泳上樣緩沖液,100℃變性10 min,進行SDS-PAGE,分析目的蛋白表達。
2 結果與分析
2.1 克隆與序列分析
2.1.1 奶山羊5-HTR4基因的克隆序列 克隆奶山羊5-HTR4基因包含一個完整的開放閱讀框架(ORF),長為1 293 bp,酸性氨基酸42個,堿性氨基酸81個,蛋白質分子量為:44.89 k Da,等電點:11.972。
2.1.2 奶山羊5-HTR4基因的種屬差異分析
DNAStar分析發現,奶山羊5-HTR4基因的ORF與兔 (Z50162)、豬 (NM-001173418)、人 (human-5HT1D)、狗(NM-001003280)和鼠(AK082016)的同源性分別為46.1%、47.2%、78.9%、45.8%及89.8%。Blastp序列比較分析,奶山羊5-HTR4基因氨基酸序列與鼠和人的同源性分別為88%和77%,相似性分別為83%和79%;Smart分析發現,奶山羊5-HTR4基因推測氨基酸序列信號肽與人、鼠5-HTR4基因氨基酸序列信號肽均為1~23aa,Sap B結構域均為49~126aa,結構特征與人、鼠的5-HTR4相一致。
2.2 原核表達載體的鑒定與誘導表達
2.2.1 原核表達載體p GEX-5-HTR2的鑒定 以克隆的序列設計表達引物進行PCR,得到了長度約為1 293 bp的目的片段;重組質粒經BamHⅠ和XhoⅠ酶切鑒定,切出約4 700 bp和1 293 bp的DNA片段,與理論值相符(圖2)。
2.2.2 重組質粒的誘導表達 p GEX-5-HTR4轉化的菌株,在約44.89 k Da處出現一條新生蛋白條帶,與預期結果一致,而pGEX 4T-1轉化的菌株則只在44.89 k Da處有一新生條帶,說明5-HTR4基因已在大腸桿菌中成功表達,并與GST形成融合蛋白(圖3)。
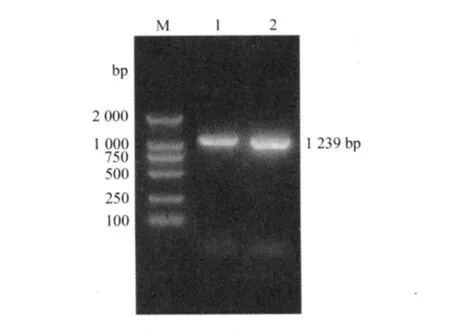
圖2 重組質粒的雙酶切鑒定結果
2.2.3 表達產物的可溶性分析 重組表達載體p GEX-5-HTR1宿主菌經IPTG誘導,離心沉淀,用化學和超聲方法裂解細菌后,SDS-PAGE凝膠電泳分析,蛋白主要以不溶性包涵體形式存在。
3 討論
5-HT分布全身,尤其在血小板和腸內更多,腦內主要是在中縫核及延腦中。迄今為止已發現人類5-HTR至少有7大類,這7種類型又可進一步分成若干亞型[8-11]。按照受體轉導方式的不同,可分為G蛋白偶聯體超家族和配體門控離子通道族兩大類。5-HTR4主要分布于基底節及新皮層區,周圍組織血管及神經節中也有分布[12]。目前5-HTR激動劑和抑制劑都是用于神經系統的藥物,治療精神疾病[13]。況且沒有專門針對5-HTR的藥物,將這些化學藥物用于奶牛,一是會影響奶牛的健康,最重要的是這些化合物會在奶里殘留,進而影響人的健康。
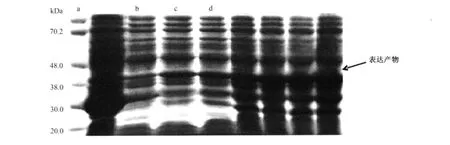
圖3 重組蛋白表達的SDS-PAGE分析
[1]Filip M,Bader on 5-HT receptors and their role physiology and pathology of the central nervous system[J].Pharmac Rep,2002,61(5):761-777.
[2]陶明,施慎,顧牛范.5-羥色胺受體的研究現狀[J].中華精神科雜志[J].1998,31(3):184-186.
[3]Wahher D J,Peter.Bashammakh S,etal.Synthesis of serotonin by second tryptophan hydroxylase isoform [J].Science,2003,29(5603):76-77.
[4]Hensler J G.Regulations of 5-HTl A receptorfunctionin brainfollowing agonist or antidepressant administration[J].Life Sci,2003,72:1665-1682.
[5]Nichols D E,Niehols C D.Semtonin receptors[J].Chem Rev,2008,108(5):1614-1641.
[6]Lanfumey,L,Hamon M.5-HT1,Receptors.Current Drug.Targe-CNSNeurologicalDisorders[J].2004,3(1):1-10.
[7]武勝昔,王亞云,劉翔宇,等.大鼠脊髓內5-HT1,5-HT3,5-HT6和5-HT7受體亞型mRNA的表達[J].中國神經科學雜志,2003,19(2):69-73.
[8]MnieFilali O,Lamb6sSenas L,Zimmer L,etal.5-HT7 receptor antagonists as a new class of antidepressants[J].Drug News Pespect,2007,20(10):613-618.
[9]Clada P,Puig,Amarg6s-Bosch M,etal.The therapeutic role of 5-HTl A and 5-HT2A receptors in depression[J].J Psychiatr-Neuresoi,2004,29(4):252-265.
[10]Dueottet C,Griebel.Effects of the selective nonpeptide cortieotropinreleasing factor receptor antagonist antalarmin inthe chronic mild stress model ofdepression in mice[J].Prog Neure·psychopharmacol BiolPsychiatry,2003,27(4):625-631.
[11]Banesr M,Agomelatine,a new antidepressant,induces regional changes in hippocampal neurogenesis[J].Biol Psychintzy,2006,59(11):1087-1096.
[12]Hedlund PB.The5-HT1 receptor influences stercotypic behavior in a model of obsessive-compulsive disorder[J].Neurosci Lett.2007,414(3):247-251.
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
