棗樹果實水分含量變化的研究
2012-10-23 00:48:22楊延青盧桂賓
山西林業科技 2012年1期
楊延青,盧桂賓,劉 和
(1.山西省林業科學研究院,山西 太原 030012;2.山西農業大學,山西 太谷 030801)
水分是樹體的重要成分之一,水分經過吸收、運輸、分布及散失循環過程,支撐著樹體的整個生命活動。自由水和束縛水的含量與樹體的生長及抗性有密切關系。自由水較多時,代謝較強,生長較快;而束縛水較多時,抗性較強。故在植物生理的研究上,常將束縛水和自由水作為重要指標進行測定。棗裂果與樹體和環境的水分變化有密切關系,因此,關于棗樹水分代謝的研究具有重要意義。關于果樹水分的研究,國內外對其它樹種已有報道,而對棗樹水分代謝的研究,尤其是棗樹體內水分動態變化的研究很少。筆者通過試驗對棗樹果實含水量的周期變化進行了研究,旨在分析棗果實組織水分的變化規律,探討果實水分代謝活性和生長、抗性特點及相互間的關系,進一步明確棗樹水分生理和器官生長發育特點,為今后的栽培管理提供理論依據。
1 材料和方法
1.1 試驗材料和設計
試驗在山西農業大學園藝站和園藝學院實驗中心進行。試驗地年均溫9.8℃,年日照時數2400 h~3 000 h,年均降水量414.2 mm.園地平坦,砂壤土,活土層厚20 cm~30 cm,土壤有機質含量0.9%.
試材為2003年春季大田栽培的梨棗(Z.jujuba Mill.Lizao)(鮮食、易裂)、金絲小棗(Z.jujuba Mill.Jinsixiaozao)(制干、易裂)和陜抗2號(Z.jujuba Mill.Shankang No.2)(兼用、較抗裂),南北行向,株行距2 m×4 m,紡錘形整枝,常規管理。試材順序排列,選株型、樹勢一致的植株為試驗樹,3株為1小區,3次重復。
1.2 采樣與處理
選取每株樹樹冠中部多年生2次枝棗股抽生的棗吊中部生長正常且發育期一致的5個~7個果實。每7 d采樣1次,時間為上午9點至10點。采后放于冰壺帶回試驗室立即處理,進行測定。
1.3 測定內容與方法
采用馬琳契克法測定各器官的總含水量(TWC)和自由水含量(FWC),并計算出束縛水含量(BWC)。用重量法測定土壤含水量(SWC)。計算公式如下:

1.4 數據分析
用SAS軟件對數據進行相關性分析,用Excel軟件作圖。為使曲線圖清晰,含水量變化趨勢圖的觀察時間為2周1次。
2 結果分析
2.1 棗樹果實含水量變化
2.1.1 棗樹果實總含水量變化
不同品種棗樹果實總含水量變化如圖1.

圖1 不同品種棗樹果實總含水量變化
由圖1可知,幼果期棗樹果實總含水量緩慢增加,白熟期保持較穩定的水平,到全紅期總含水量快速下降,直至采收。果實總含水量由初期的79%左右升高到85%左右,較幼果期升高了6%.
果實生長發育過程中,各品種總含水量的變化趨勢相近,但在不同時期存在差異。幼果期金絲小棗果實的總含水量顯著低于陜抗2號和梨棗果實,到白熟期梨棗果實的總含水量顯著高于金絲小棗和陜抗2號。同時,金絲小棗總含水量低且平穩,陜抗2號出現波動,變化較大。
2.1.2 棗樹果實自由水含量變化
不同品種棗樹果實自由水含量變化如圖2.
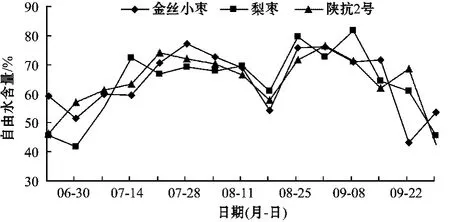
圖2 不同品種棗樹果實自由水含量變化
從圖2可以看出,棗樹果實自由水含量變化近似于“M”形,與棗果實雙“S”形的生長曲線變化一致。幼果期自由水含量處于增長階段,7月中、下旬為棗果實快速增長期,出現了第1次增長高峰,自由水含量分別為:金絲小棗77.10%,陜抗2號73.95%,梨棗72.47%,平均值為75.17%.8月中旬為果實生長緩慢期,果實自由水含量處于低谷,分別為:梨棗60.90%,陜抗2號57.76%,金絲小棗54.36%,平均值為57.67%.8月下旬至9月上旬為果實第2次快速生長期,果實代謝旺盛,出現第2次增長高峰,自由水含量分別為:梨棗81.65%,陜抗2號76.30%,金絲小棗76.12%,平均值為78.02%.隨后含水量快速下降,直至采收。分析數據可得出,棗果實幼果期、第1次快速生長期、緩慢生長期、第2次快速生長期的自由水含量比值為1.00∶1.50∶1.15∶1.55.
棗樹不同品種果實整個生長發育期內自由水含量的變化相近。但在幼果期,梨棗和金絲小棗果實自由水含量先降后升,陜抗2號果實自由水含量持續升高。
2.1.3 棗樹果實束縛水含量變化
不同品種棗樹果實束縛水含量變化如圖3.

圖3 不同品種棗樹果實束縛水含量變化
由圖3可以看出,棗樹果實束縛水含量變化近似于“W”形,與棗果實雙“S”形的生長曲線變化相反。6月底至7月下旬期間迅速下降,7月下旬出現第1次最低值,束縛水含量分別為:梨棗10.00%,陜抗 2號 9.67%,金絲小棗 7.40%,平均值為9.02%.隨后含水量升高,在8月中旬出現峰值,分別為:金絲小棗29.95%,陜抗2號24.12%,梨棗19.94%,平均值為24.67%.在9月上旬出現第2次最低值,分別為:梨棗4.70%,陜抗2號9.12%,金絲小棗8.17%,平均值為7.33%.然后再次升高,直至果實成熟。由數據可知,棗果實幼果期、第1次快速生長期、緩慢生長期、第2次快速生長期的束縛水含量比值為1.00∶0.31∶0.86∶0.26.
不同棗品種果實束縛水含量變化趨勢相似。但在6月下旬,金絲小棗和梨棗的果實束縛水含量先升后降,而陜抗2號呈下降趨勢。8月中旬不同品種峰值顯著不同,其中金絲小棗束縛水含量最高,梨棗束縛水含量最低。在緩慢生長期金絲小棗自由水含量最低,但束縛水含量最高,梨棗與金絲小棗正好相反,表明,此時梨棗果實的代謝活性高于金絲小棗。
綜上所述,棗樹果實總含水量在幼果期到白熟期基本穩定,全紅期后總含水量迅速下降。試驗結果與劉嘉芬的結果相同,在棗果發育過程中,從幼果至硬核期前,水分含量最高約達85%以上,在此期間水分含量變化非常緩慢;全紅期水分含量迅速降低。自由水和束縛水含量變化趨勢相反,此變化反映了棗果實的雙“S”形曲線生長發育過程。果實在幼果期、第1次快速生長期、緩慢生長期和第2次快速生長期的束縛水與自由水含量比值為1.0∶1.8,1.0 ∶8.3,1.0 ∶2.4,1.0 ∶10.6,此結果反映了果實在不同時期自由水與束縛水的相互轉化。幼果期金絲小棗果實的總含水量顯著低于陜抗2號和梨棗果實,白熟期梨棗的總含水量和自由水含量顯著高于金絲小棗和陜抗2號。
2.2 棗樹果實含水量的相關性
棗樹果實總含水量(TWC)、自由水含量(FWC)及束縛水含量(BWC)之間的相關性見表1.

表1 棗樹果實含水量相關性
由表1可以看出,棗樹果實總含水量與自由水含量呈極顯著正相關,與束縛水含量相關性不顯著;自由水含量與束縛水含量呈極顯著負相關。果實內的水分以自由水為主,自由水含量的增減會使得果實總含水量水平相應的升降。果實內自由水和束縛水含量出現相反的變化,以適應不同條件下果實生長發育的需求。當果實代謝活躍時,自由水含量大幅度提高,而束縛水含量卻相應減少,此時表現出果實代謝旺盛而抗性低,反之亦然。
2.3 棗樹枝葉與果實含水量的相關性
棗樹枝條、葉片與果實含水量的相關性見表2.
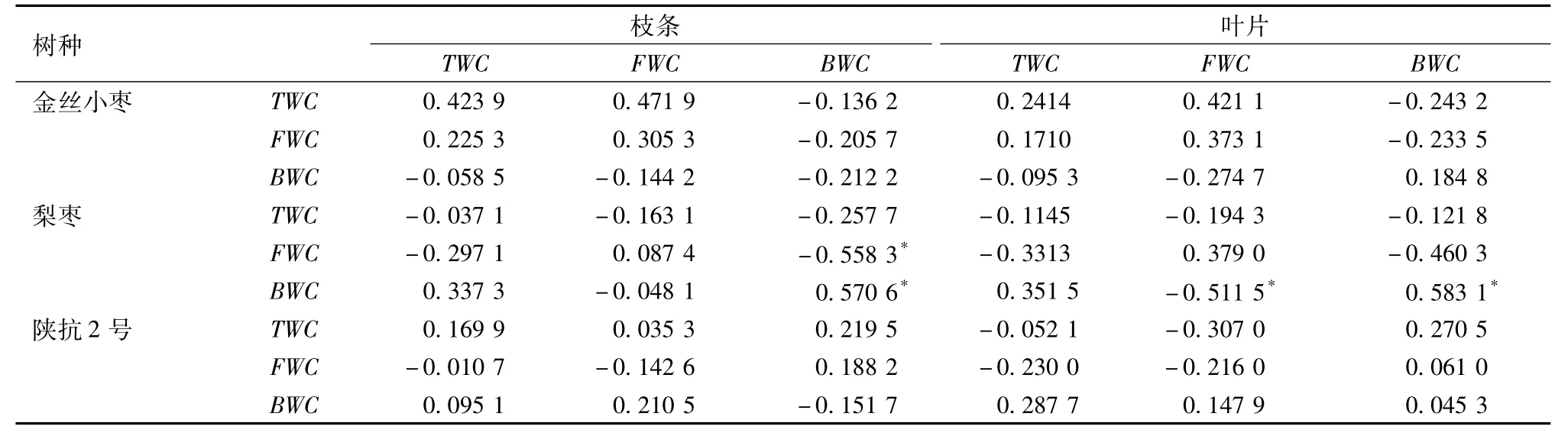
表2 棗樹枝葉與果實含水量的相關性
根據表2所示,棗樹枝條、葉片與果實的含水量相關性均不顯著。這可能是由于在正常情況下,果實與枝條、葉片的水分代謝變化規律均不相同,受到環境影響時由于其各自適應能力的差異,所表現出的反應也不一樣。從另一方面可以看出,果實內的水分變化受枝條或葉片的影響很小,在遇到水分脅迫時,水分的運輸及各器官的相互關系可能會發生變化,但同樣也證明了果實自身水分的改變受環境的影響更大。所以,影響裂果的主要原因應是環境水分因素。
綜上所述,在枝條、葉片和果實內總含水量與自由水含量均表現出了顯著的正相關性,葉片和果實內也均表現出自由水和束縛水的極顯著負相關性。枝條束縛水含量與葉片總含水量、自由水含量呈顯著或極顯著負相關,與葉片束縛水含量呈極顯著正相關。棗樹枝條、葉片與果實的含水量相關性均不顯著。
3 結論
1)棗樹果實總含水量在幼果期到白熟期基本穩定,到全紅期后迅速下降。自由水含量變化近似于“M”形,束縛水含量變化近似于“W”形,與其生長發育動態相符。果實的總含水量變化不能完全反映果實的生長發育規律,而果實自由水和束縛水含量的變化規律可以反映在不同時期果實生長的內在變化情況。
2)棗樹果實總含水量與自由水含量呈極顯著正相關,與束縛水含量相關性不顯著;自由水含量與束縛水含量呈極顯著負相關。果實內自由水和束縛水含量出現相反的變化,以適應不同條件下果實生長發育的需求。當果實代謝活躍時,自由水含量大幅度提高,但束縛水含量卻相應減少,此時表現出果實代謝旺盛而抗性低,反之亦然。
由于棗樹枝條、葉片與果實含水量的相關性均不顯著,所以,引起棗裂果的關鍵因素可能是環境水分。器官水分含量變化與裂果性狀的關系需進一步研究。
[1]潘瑞熾,主編.植物生理學[M].第3版.北京:高等教育出版社,2000:6-9.
[2]王慶江.光照、溫度、二氧化碳、水分等生態因子對贊皇大棗光合作用的影響[D].河北:河北農業大學,2002.
[3]賀潤平.棗樹冬季休眠特性的研究[D].山西:山西農業大學,2006.
[4]姜衛兵,高光林.水分脅迫對果樹光合作用及同化代謝的影響研究進展[J].果樹學報,2002,19(6):416-420.
[5]劉 和,楊佩芳,高美英.駿棗枝葉水勢周年動態研究[G]∥中國科協第二屆青年學術年會園藝學論文集.北京:北京農業大學出版社,1995:87-90.
[6]喬富廉.植物生理學實驗分析測定技術[M].北京:中國農業科學技術出版社,2002:5-8.
