不同基質對葛蘿槭光合特性日變化的影響
2012-10-23 00:48:22原海云姚延梼
山西林業科技 2012年1期
原海云,姚延梼
(1.山西省關帝山國有林管理局,山西 文水 032104;2.山西農業大學,山西 太谷 030801)
葛蘿槭(Acer grosseri Pax)為槭樹科槭樹屬落葉喬木,冠幅較大,果實美觀,適應性強,在環境景觀建設中作為重要的彩色觀賞植物被廣泛應用。近幾年,關于槭樹科植物形態學、細胞學、化學成分等方面的研究較多,葉片生理特性的研究甚少。李倩中等研究了青楓和紅翅槭的光合速率,邱業先對槭樹科植物葉蛋白酶活性的季節變化進行了探索,而對于葛蘿槭光合特性研究尚未見報道。光合作用是植物生長發育的基礎和生產力高低的決定性因素,主要受到光合有效輻射、環境溫度、濃度、葉片生理成熟度的影響。研究植物光合特性的日動態變化特征有利于反映其對環境的適應能力,明確樹木的生態學特性。筆者對不同基質處理的葛蘿槭光合特性日變化規律進行了系統研究,探討了不同基質處理下葛蘿槭光合生產能力的差異,篩選出最佳基質配方,為今后葛蘿槭的栽培和資源開發利用提供參考。
1 材料與方法
1.1 試驗材料與設計
1.1.1 試驗材料
采用平均苗高7.36 cm,平均地徑0.282 cm的1年生葛蘿槭扦插苗,于2008年11月在山西省林業廳實驗苗圃陽曲縣基地的溫室內進行移植。試驗于2009年8月上旬進行,以生長在苗圃溫室內移植1年生的葛蘿槭(Acer grosseri Pax)葉片為試驗材料。培養基質有3種:1)醋渣,松針,菇渣。2)松針,菇渣,粉煤灰,羊糞。3)松針,醋渣,粉煤灰,羊糞。
1.1.2 試驗設計
試驗采用單因素隨機區組排列法,共設3個處理,1個對照(田園土)。每個處理20株,3次重復。
1.2 方法
采用北京雅欣理儀科技有限公司生產的CB-1102便攜式光合蒸騰儀完成各項光合生理生態指標的測定。測定時,直接取植株(選生長勢相近,上部發育狀態一致的植株)中上部葉5片,每片測量1次,取平均值。測定時間為早上8:30至下午18:30,時間間隔2 h.測定指標包括光合速率(Pn)、氣孔導度(C1)、細胞間隙CO2濃度(CO2int)、蒸騰速率(E)。采用Excel和Spss 11.0對數據進行統計分析并制作圖表。
2 結果分析
2.1 凈光合速率日變化
不同基質處理葉片的凈光合速率日變化如圖1.
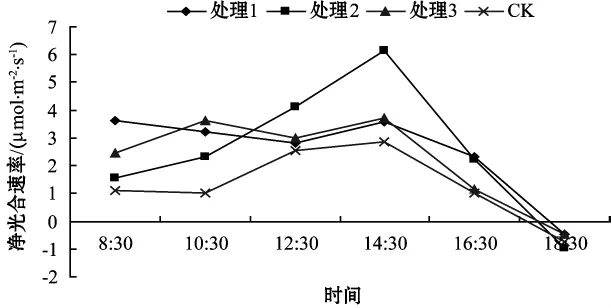
圖1 不同基質處理葉片的凈光合速率日變化
從圖1可看出,不同基質處理下葛蘿槭的Pn日變化曲線不一致。處理1,處理2和CK的Pn日變化曲線呈單峰型,處理1和CK從8:30到10:30呈現下降趨勢,可能與葉片所處的生態環境和太陽光較弱有關。隨后呈現上升趨勢,在下午14:30達到最大值。處理2隨著光照強度的增加呈上升趨勢,在下午14:30 達到最大值,為6.165 μmol/(m2·s),之后呈現下降趨勢。處理3的Pn日變化曲線呈雙峰型,從8:30到10:30,Pn呈上升趨勢,達到最大值。在12:30時,3種處理的葛蘿槭表現出明顯的“午休”現象。引起光合“午休”的主要原因是大氣與土壤干旱,在干熱的中午,葉片蒸騰失水加劇,使土壤水分虧缺。植株的失水大于吸水,就會引起萎焉與氣孔導度降低,使CO2吸收減少。“午休”后處理3再次出現急劇增長的趨勢,下午14:30出現的峰值高于上午10:30出現的峰值。最后由于光照強度的減弱,光合速率開始減小。3種處理的Pn在18:30都出現了負值,說明這時植物的光合積累不夠其自身的消耗,呼吸消耗占其主要地位。同時光照強度降低,溫度降低,光補償點降低。總體看,凈光合速率排序為處理2>處理3>處理1>CK.
2.2 蒸騰速率日變化
不同基質處理葉片的蒸騰速率日變化如圖2.
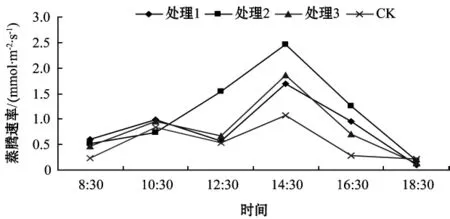
圖2 不同基質處理葉片的蒸騰速率日變化
從圖2可看出,不同基質處理下葛蘿槭E日變化曲線呈現單峰型和雙峰型兩種類型。處理2的E日變化曲線呈現單峰型,隨著光照強度的增強和氣溫的升高,E逐漸增大,下午14:30達到最大值,之后逐漸下降。處理1,處理3及CK葉片的E日變化曲線均呈現雙峰型,在10:30出現了第1次高峰,而后逐漸下降;至12:30,3者均出現明顯的“午休”現象;第2次峰值出現在14:30,而處理3的E日變化明顯高于其它處理。
2.3 氣孔導度日變化
不同基質處理葉片的氣孔導度日變化如圖3.

圖3 不同基質處理葉片的氣孔導度日變化
由圖3可知,不同基質處理下葛蘿槭C1日變化曲線呈現單峰型和雙峰型兩種類型。CK的C1日變化呈現雙峰型,第1次峰值出現在10:30,第2次峰值出現在14:30,之后C1持續下降,約18:30有緩慢增加的趨勢。處理1和處理3均屬單峰型,且C1日變化曲線類似,呈降低—升高—降低的趨勢,最小值出現在12:30,峰值都出現在下午14:30,而處理1在16:30達到了最大值。處理2也表現為降低—升高—降低的曲線,但谷底出現在10:30,在14:30達到最大值,為 63.6 mmol/(m2·s).
2.4 細胞間隙CO2濃度日變化
不同基質處理葉片的細胞間隙CO2濃度日變化如圖4.
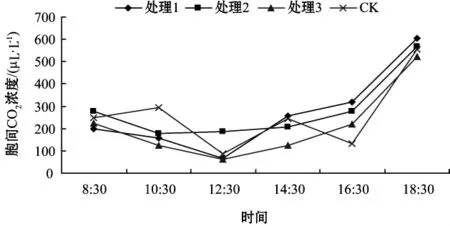
圖4 不同基質處理葉片的細胞間隙CO2濃度日變化
從圖4可看出,3個處理胞間CO2濃度在清晨較高,8:30之后呈下降趨勢,最小值出現在12:30,之后呈上升趨勢,在18:30達到最大值。CK的胞間CO2濃度呈現升—降—升—降—升的趨勢,在整個趨勢中分別在上午10:30和下午14:30形成了兩個小高峰,在18:30達到了一天中的最大值。
3 結論
1)處理1,處理2和CK葉片的凈光合速率日變化曲線呈現單峰型,凈光合速率大小為處理2>處理1>CK.處理2的光飽和點出現在下午14:30,且蒸騰速率、氣孔導度與光合速率的變化曲線一致,存在著明顯的正相關關系。處理2(松針、菇渣、粉煤灰、羊糞)有利于光合作用的順利進行和光合產物的積累,為最佳基質配方。
2)處理3的葛蘿槭葉片凈光合速率為典型雙峰曲線,第1次高峰出現在10:30,第2次出現在14:30.凈光合速率最小值出現在12:30,表現出明顯的“午休”現象,且蒸騰速率與葉片的光合速率有顯著的正相關關系,呈平行變化趨勢。由于“午休”造成的損失可達光合生產的30%,緩減“午休”現象的情況下,處理3(松針、醋渣、粉煤灰、羊糞)基質配方利于提高葛蘿槭光合生產力。
[1]王 忠.植物生理學[M].北京:中國農業出版社,2000.
[2]邱業先,陳尚钘,杜天真,等.幾種槭樹科植物葉蛋白酶活性的季節變化[J].江西農業大學學報,2003(5):652-655.
[3]李倩中,蘇家樂,陳尚平,等.兩種類型槭樹光合作用及其影響因子初探[J].江西農業學報,2007,19(9):35-37.
[4]張 璐,張紀林.不同光照條件下3種冬青屬植物的光合特征日變化研究[J].西北植物學報,2006,26(3):490-495.
[5]蘇培璽,杜明武,張立新,等.日光溫室草莓光合特性及對CO2濃度升高的響應[J].園藝學報,2002,29(5):423-426.
[6]張川紅,沈應柏,尹偉倫,等.鹽脅迫對幾種苗木生長及光合作用的影響[J].林業科學,2002,38(2):27-31.
[7]高 超,閆文德,田大倫,等.杜仲光合速率日變化及其與環境因子的關系[J].中南林業科技大學學報,2011(5):100-104.
[8]孫佳音,楊逢建,龐海河,等.遮陰對南方紅豆杉光合特性及生活史型影響[J].植物研究,2007,27(4):439-444.
[9]石玉波,卓麗環,辛雅芬,等.不同遮光處理對白櫟光合生理特性的影響[J].安徽農業科學,2010(2):956-958.
[10]劉厚誠,雷 雨,陳日遠.遮光處理對節瓜光合作用特性的影響[J].植物資源與環境學報,2005,14(3):33-36.
[11]燕麗萍,金 芳,鄭平生.四種草莓光合特性的研究[J].甘肅農業大學學報,2004,12(6):620-624.
[12]徐立祥,唐雪東,劉曉嘉.根外施鉀對K9蘋果樹光合速率的影響研究[J].北方園藝,2010(4):23-26.
[13]王立新,田 麗.2種彩葉植物光合特性的研究[J].安徽農業科學,2010(2):715-717.
[14]吳永波,薛建輝.鹽脅迫對3種白蠟樹幼苗生長與光合作用的影響[J].南京林業大學學報:自然科學版,2002,26(3):19-22.
