不同種源無患子苗期生長性狀遺傳變異研究
2012-11-24 02:11:12邵文豪岳華峰姜景民董汝湘刁松鋒
浙江林業科技 2012年1期
關鍵詞:生長
邵文豪,岳華峰,姜景民*,董汝湘,刁松鋒
(1.中國林業科學研究院 亞熱帶林業研究所,浙江 富陽 311400;2.中國林業科學研究院 經濟林研究開發中心,河南 鄭州 450003)
無患子(Sapindus mukorossi)為無患子科無患子屬落葉喬木,廣泛分布于我國東部、南部至西南部[1]。無患子是亞熱帶地區重要的園林綠化樹種[2],其果皮含有表面活性物皂苷,是優良的天然洗滌劑[3~5],其種子含油率高,是開發生物質能源的理想原料[6]。目前,國內外對無患子果皮皂苷成分提取、測定的研究較多[7~11],對其種源選擇、定向選育等方面研究較少,盧芳[12]、王荔英[13]曾開展無患子引種對比試驗,但多為區域性種源材料測試結果。本文通過系統收集無患子自然分布區內不同種源、家系種子進行播種育苗,對1年生幼苗生長進行定期觀測,研究無患子不同種源苗期生長變異規律及其與種源地生態因子之間的關系,選擇無患子苗期生長優良種源、家系,以期為無患子種源區劃和遺傳改良提供理論依據。
1 材料與方法
2008年10-11月,在無患子不同分布區內進行果實采集工作,選擇正常生長、胸徑20 cm左右的無患子陽面中上部成熟果實,單株采集果實不少于2 kg,每采種母樹間距要求在50 m以上,共收集到9個種源90個家系的果實材料(表1)。果實采集帶回試驗室后,將種子剝離,低溫層積貯藏備播。播種試驗設在浙江省杭州市余杭長樂林場西山苗圃,地理位置22° 21′ N,120° 23′ E,海拔18 m,年均溫15.3℃,年降水量1 313 mm。2009年3月進行田間播種試驗,采用完全隨機區組設計,4次重復。

表1 無患子不同種源地生態因子Table1 Ecological factors of different provenances of S.mukorossi
幼苗出土后開始觀察記錄,4月15日出苗基本整齊,30日開始測量苗高,5月15日開始測量地徑。測量時,在每個重復的每個家系中隨機選取15株苗木作為固定樣株,每隔15 d測定苗高和地徑生長量。
應用Excel2003、SPSS18.0統計軟件進行數據分析。
2 結果與分析
2.1 不同種源無患子苗高和地徑生長節律
由圖1、圖2可知,5月30前為苗高生長初期,此階段溫度不高,生長較為緩慢,隨后苗木開始快速增長,8月30日至9月15日,每15 d的苗高凈生長量達到高峰,此后開始減小,直到11月15日至12月15日凈生長量減到最低。河南西峽種源為北部種源,其苗高凈生長量停止時期明顯早于其它種源。地徑的生長節律與苗高略有不同,5月 30日前為地徑凈生長初期,每15 d的地徑凈生長量在9月15日至9月30日達到高峰,此后,地徑的凈生長量逐漸減小,直到 12月中旬。可見,無患子苗高生長與地徑生長不完全同步,苗高生長高峰早于地徑生長高峰。
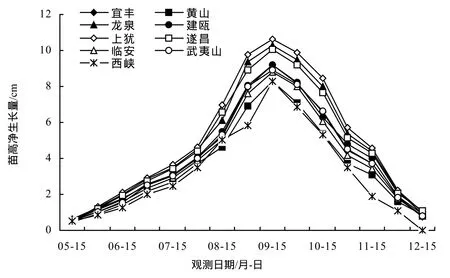
圖1 不同種源無患子1年生苗高生長節律Figure1 Height growth rhythm of 1-year seedling of S.mukorossi
依據無患子1年生苗生長節律特點,將1年生苗生長劃分為3個時期:生長初期、速生期和硬化期。生長初期是從開始發芽到苗高和地徑生長大幅度增加為止,大致從 4月初至5月30日,苗木在此時期增加體內干物質的積累,為生長高峰做準備[14];速生期從5月 30日至10月15日,該時期溫度相對較高,水分充足,苗木生長最快;10月下旬日照時間縮短,氣溫陸續下降,苗木生長日趨減慢進入硬化期。苗木木質化程度高,抗逆性強,為越冬做準備,這是苗木對自然環境的適應結果。
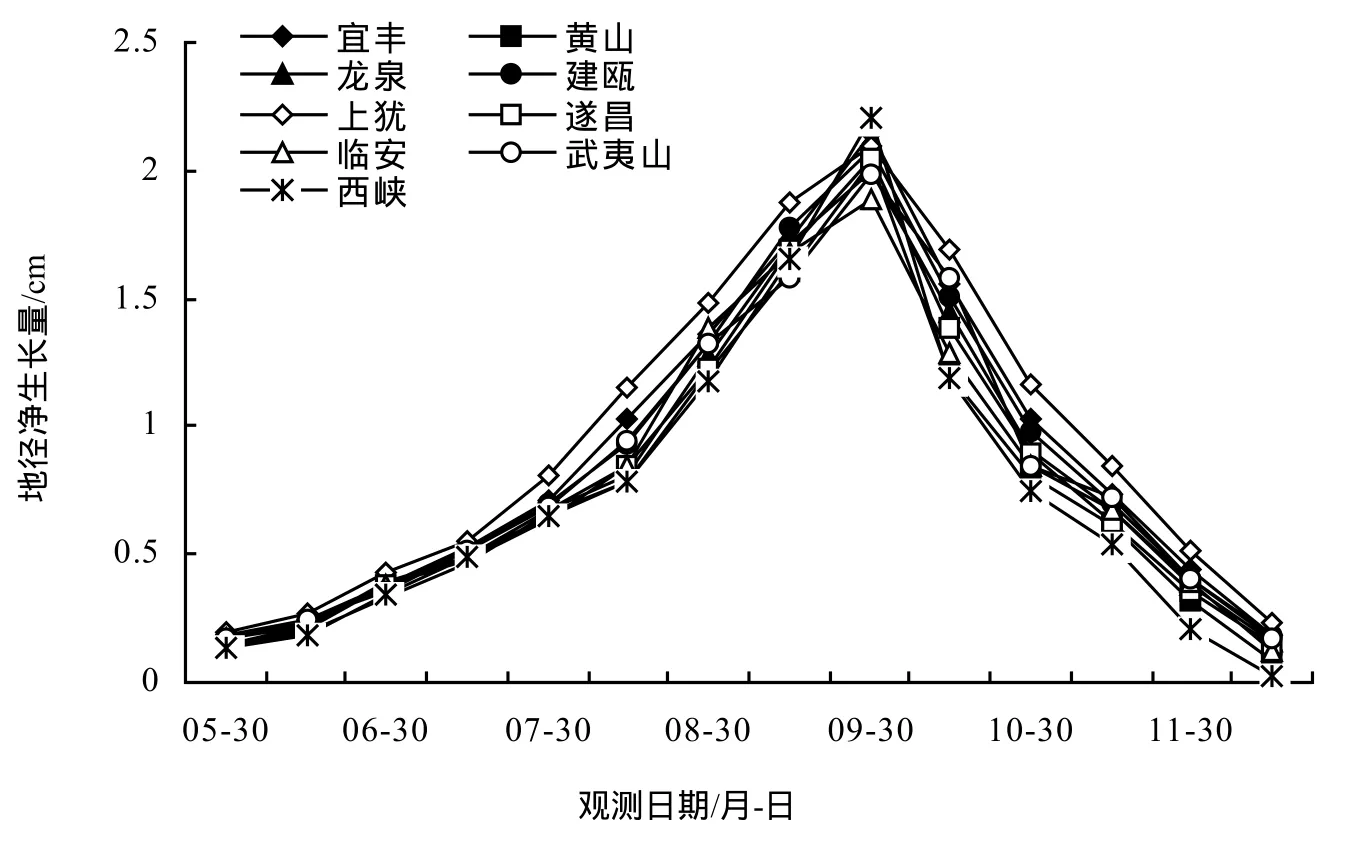
圖2 不同種源無患子1年生地徑生長節律Figure2 Ground diameter growth rhythm of 1-year seedling of S.mukorossi
2.2 無患子苗期生長性狀的遺傳變異
2.2.1 不同種源間苗高、地徑生長的變異性分析 苗木硬化期后,對不同種源的無患子苗高、地徑性狀進行觀測,方差分析結果表明(表2),不同種源間苗高和地徑生長差異均達到極顯著水平。從遺傳參數來看,苗高、地徑的遺傳力均較高,分別為0.923和0.869,說明無患子種源苗高、地徑的變異主要是由種源本身的遺傳特性引起的。苗高性狀變異系數為24.73%,大于地徑的變異系數15.89%,說明苗高比地徑更具有可選擇性。

表2 不同種源無患子苗期性狀生長方差分析及遺傳估算Table2 ANOVA and genetic estimation of seedling growth traits from different provenances
苗高和地徑生長量是判斷苗木質量優劣的重要指標。由表3可知,無患子各種源間1年生苗高、地徑均值分別為67.07 cm、11.78 mm,其中江西上猶種源苗高均值最大,其次是浙江龍泉和遂昌;苗高均值最小的是最北部種源河南西峽,偏北部的黃山和臨安種源苗高均值亦較小。地徑生長均值最大的是上猶種源,最小的是河南西峽。總體而言,江西上猶和浙江龍泉種源苗期生長較好,而河南西峽、安徽黃山種源相對較差。

表3 不同種源無患子1年生苗高、地徑均值Table3 Mean of height and ground diameter of 1-year seedling from different provenances
2.2.2 不同時期無患子苗高、地徑生長的差異性比較 對無患子不同家系1年生苗高、地徑生長量進行方差分析(表4)可見,不同時期苗高、地徑生長在家系間差異均達到極顯著水平。苗高性狀遺傳力在各時期均較大,都在 0.8以上,說明苗高受遺傳控制程度較高,可選擇性更強,而地徑的家系遺傳力初期較低,隨苗木生長其遺傳力逐漸增大,說明無患子苗期選擇應注重生長后期,11月中旬以后苗木生長進入比較穩定的狀態,可以進行優良家系選擇。
2.3 無患子苗期生長性狀與種源地生態因子相關性分析
從表5可知,苗高生長性狀與種源地緯度極顯著負相關,與種源地年均溫顯著正相關,與種源地經度、年降水量、年均日照、無霜期和海拔等相關性不顯著,說明苗高性狀隨種源地緯度南移,年均溫升高,生長極顯著升高,而地徑性狀僅與種源地年平均溫度呈顯著正相關。
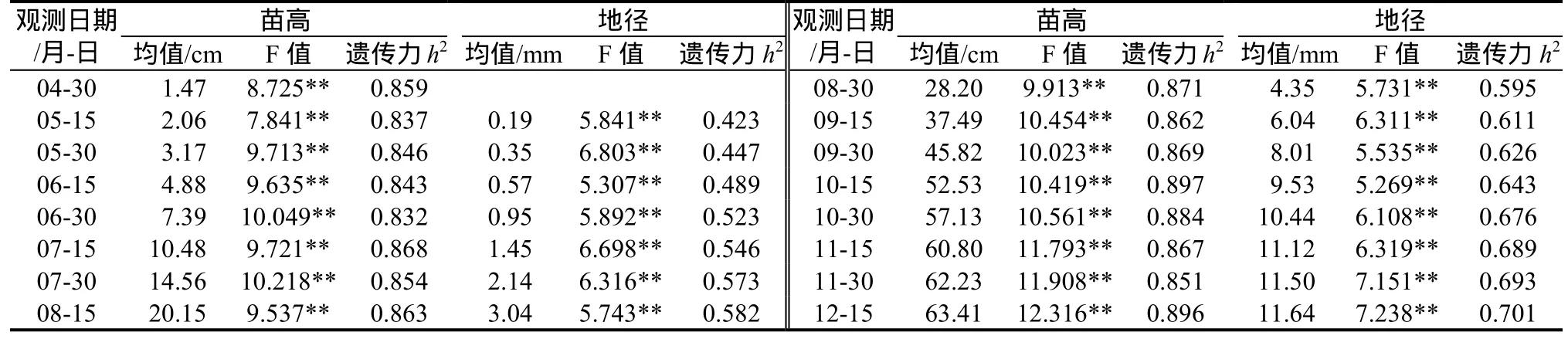
表4 無患子不同生長時期性狀差異分析及遺傳估算Table4 ANOVA and genetic estimation of seedling traits at different growth periods

表5 無患子苗期生長性狀與種源地生態因子間相關性Table5 Correlation between seedling traits and ecological factors of different provenances
2.4 無患子優良種源、家系的苗期選擇
通過對苗高、地徑生長表現及性狀遺傳增益的估算(表6),初步選擇出江西上猶、浙江龍泉和江西宜豐3個優良種源,上猶7號、上猶8號、上猶9號、宜豐1號、宜豐6號、龍泉4號以及龍泉10號共計7個優良家系。無患子廣布我國長江以南地區,地理環境的變化導致各地無患子種群間產生巨大的差異,因此從中選擇優良種源具有較大的潛力,本文初步選擇出的無患子優良種源和家系均來自無患子自然分布區的南部種源,南部地區應是今后繼續深化無患子種源選擇的重點關注地區。
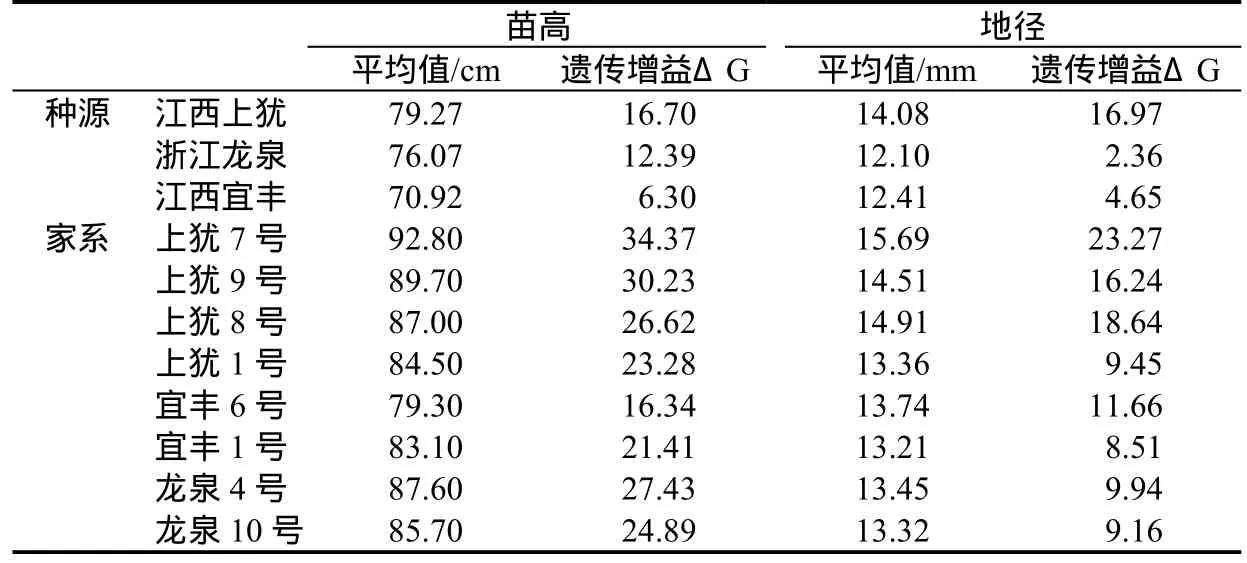
表6 無患子優良種源、家系苗期性狀遺傳增益估算Table6 Genetic gains estimation of seedling traits of superior provenances and families
3 小結
掌握苗木在一年中的生長動態過程,可以為苗期管理提供科學依據,培育出優質苗木。在苗木生長的速生期要加強肥水管理,及時除草,防止病蟲害,使苗木快速長大,而在苗木硬化期要停止一切促進苗木生長的管理措施,提高苗木抗逆性,尤其是對低溫和干旱的抵抗能力,防止苗木徒長不能越冬。無患子1年生苗生長期大致分為生長初期、速生期和硬化期,其中生長初期時間較短,速生期時間較長,從5月下旬持續到10月上旬。無患子苗高生長與地徑生長不完全同步,地徑生長高峰要遲于苗高生長高峰,說明無患子苗木在生長過程中存在營養分配的重心轉移。
不同種源苗高、地徑生長差異明顯,苗高、地徑生長最好的是江西上猶種源,生長最差是北部種源河南西峽。通過對遺傳參數的估算表明,苗高、地徑生長性狀均具有較高的遺傳力,同時,相關性分析表明苗高生長性狀與種源地緯度極顯著負相關,與種源地年均溫顯著正相關,說明南部種源無患子苗高生長明顯好于北部種源,而地徑性狀僅與種源地年均溫呈顯著正相關。
苗期試驗是早期選擇的重要途徑之一,其目的一是了解各種源苗期的差異,初步掌握種源的地理變異趨勢,二是培育造林試驗用的苗木。國內外研究表明,雖然選擇的效果依對象和條件而異,但將苗期觀測的項目作為早期選擇指標是有效可行的[15]。在進行良種選育時,將種源選擇和家系選擇結合起來,既能利用種源內的遺傳變異,又能利用種源間的遺傳變異,從而選出不同培育目標的最佳種源或家系。本文初步選擇出無患子3個優良種源7個優良家系,均表現出明顯的苗期生長優勢,遺傳增益顯著,但是,以上僅是無患子不同種源苗期生長性狀的初步研究結果,其結論尚需在區域性造林試驗中進一步驗證和選擇。
[1]中國科學院中國植物志編輯委員會.中國植物志:47(1)[M].北京:科學出版社,1985.14-15.
[2]范理璋.無患子育苗技術[J].林業實用技術,2000(12):18-19.
[3]孫潔如,陳孔常,周鳴方,等.無患子表面活性物及其復配體系的性質研究[J].日用化學工業,2002,32(4):16-18.
[4]Huang H C,Tsai W J,Liaw C C,et al.Anti-platelet aggregation triterpene saponins from the galls of Sapindus mukorossi[J].Chem Pharm Bull,2007,55(9):1 412-1 415.
[5]Kuo Y H,Huang H C,Yang L M,et al.New dammarane-type saponins from the galls of Sapindus mukorossi[J].J Agri Food Chem,2005(53):4 722-4 727.
[6]黃素梅,王敬文,杜孟浩,等.無患子籽油脂肪酸成分分析[J].中國油脂,2009,34(12):74-76.
[7]魏鳳玉,方春.酶法提取無患子皂苷的工藝研究[J].應用化工,2010,39(8):1 149-1 151.
[8]饒厚曾,郭隆華.無患子皂苷提取工藝研究[J].江西科學,2002,20(1):55-58.
[9]饒厚曾,桑成濤.微波萃取法提取無患子皂苷工藝[J].遼寧石油化工大學學報,2006,26(4):70-72.
[10]魏鳳玉,解輝,余錦城,等.超濾法分離提純無患子皂苷[J].膜科學與技術,2008,28(2):85-88.
[11]Jie Ru Sun,Kong Chang CHENG,Tie Yin PAN,et al.A new acyclic sesquiterpene oligoglycoside from pericarps of Sapindus mukorossi[J].CN Chem Let,2002,13(6):555-556.
[12]盧芳,吳雨龍.無患子種源引種苗期試驗[J].林業實用技術,2004(7):26-27.
[13]王荔英,姚湘明.無患子種源引種苗期試驗初報[J].林業試驗勘察設計,2009(2):68-71.
[14]李冬林,金雅琴,向其柏.浙江楠苗期生長節律[J].浙江林學院學報,2004,21(3):349-352.
[15]佘誠棋,楊萬霞,方升佐,等.青錢柳種源間苗期性狀變異分析[J].南京林業大學學報(自然科學版),2010,34(1):34-38.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
