重組β-葡萄糖苷酶基因和內切葡聚糖苷酶基因在大腸桿菌中的共表達
2013-02-21 12:59:06唐自鐘韓學易
食品工業科技 2013年24期
唐自鐘,劉 姍,2,韓學易,陳 惠,*,孫 蓉,單 志,茍 琳,吳 琦
(1.四川農業大學生命科學與理學院,四川雅安625014;2.攀枝花學院生物與化學工程學院,四川攀枝花617000)
當代社會高速發展,食品原材料和能源等問題越來越受到世界人民的關注,纖維素是自然界中含量最多,最豐富的材料,如何利用這一巨大的資源成為研究者最為關心的問題。最早美國進行纖維素材料降解防護問題時發現纖維素經微生物降解后,可產生經濟、豐富的生產原料,在擴大食品工業原料和植物原料的綜合利用,提高原料利用率,凈化環境和開辟新能源等方面具有十分重要的意義,且有望解決自然界不斷產生的固體廢物問題,于是纖維素酶得到了廣泛的關注。纖維素酶在動物飼料、紡織、食品加工、污水處理、中草藥有效成分提取等行業得到廣泛應用,有效地改善產品質量,提高產量,具有良好的經濟效益[1-3]。
目前纖維素酶的研究主要集中在菌種的篩選,單一基因的克隆與表達,及利用分子技術對已研究的纖維素酶進行合理化和非合理化突變,以期望得到高酶活的基因。W ilson等[4]利用定點突變技術對T.fusca的內切葡萄糖苷酶Cel6A進行突變,獲得了15個表面突變體和5個環突變體,使其對羧甲基纖維素的活性提高了175%。當前研究對于纖維素酶的整體沒有一個總的認識,纖維素酶是由三大酶類組成,它們是一個有機的整體,才能對纖維素有一個很好的降解作用[4-7],如何使其能夠共同表達,目前在大腸桿菌中其他基因的共表達已有的相關研究,如龐永奇等[8]將pET30a2Cre和pET23b2loxGFP在大腸桿菌中成功實現了不相容性共表達;范立強等[9]肉堿脫水酶基因caiB和輔酶合成因子基因caiE的共表達研究中,比較了相容和不相容表達系統,基于以前研究基礎,本研究以實驗室自主得到的β-葡萄糖苷酶基因和內切葡聚糖苷酶基因出發,構建了這兩種纖維素酶在大腸桿菌中的共表達,以期能提高降解纖維素的能力。
1 材料與方法
1.1 材料與儀器
本實驗所用的菌株與載體 列于表1;限制性內切酶BglⅡ、Bam HⅠ、XhoⅠ、T4DNA連接酶、SlutionⅠ連接酶、DNAMarker、protein Marker(Low)、高保真酶primeSTAR@HSDNA polymerase、氨芐青霉素鈉 大連TaKaRa Biotech公司;質粒提取試劑盒、瓊脂糖凝膠回收試劑盒和PCR產物純化回收試劑盒 上海OMEGA生物技術有限公司;2×PCR master m ix 天根生化科技有限公司;引物 由上海Invitrogen生物技術有限公司合成;胰蛋白陳和酵母粉 Oxoid公司;SDS-PAGE凝膠試劑盒 武漢博士德生物公司;Ni-Agarose His標簽蛋白純化試劑盒 GE公司美國;其他生物試劑 均為進口或國產分析純。
PCR儀(百樂T100)、GenePulser電穿孔儀(bio-rad Gene PulserMXcellTM)、UVP凝膠成像儀(LY-B)、恒壓恒流電泳儀(DYY-Ⅱ) 北京六一;冷凍離心機(Avanti J-E) Beckman公司等。
1.2 實驗方法
1.2.1 重組質粒的構建 根據本實驗室自行克隆得到的內切葡聚糖酶基因EG,并連于pUC18的基因序列pUC18-End和外切葡聚糖酶基因BGL并連于pMD19的基因序列pMD19-BGL,分別進行生物信息學分析,設計出不含有信號肽的引物序列,并在5’端和3’端分別引入了相應的酶切位點,引物序列及引入的酶切位點見表2。
高保真酶PCR擴增體系:EG基因片段,模板為質粒pUC18-End,擴增條件為:95℃預變性2m in;94℃變性1min,55℃退火1min,72℃延伸2min,30個循環;72℃延伸8m in。BGL基因片段,模板為質粒pMD19-BGL,擴增條件為:94℃預變性4m in;94℃變性50s,61℃退火50s,72℃延伸3m in,30個循環;72℃延伸10m in。產物經1%瓊脂糖凝膠電泳檢測回收,之后將PCR產物和表達載體質粒pET32a和pET30b進行相應的雙酶切,回收后分別進行連接:pET32a與EG相連接;pET30b與BGL相連接。連接產物轉化大腸桿菌DH 5α,經抗性篩選、菌落PCR、酶切篩選后挑取陽性轉化子,測序。結果正確的重組子命名為:pET32a-EG和pET30b-BGL。
1.2.2 重組質粒的轉化 pET32a-EG和pET30b-BGL分別用熱激法轉化入大腸桿菌表達菌株BL21(DE3),作為對照,同時將pET32a-EG和pET30b-BGL等量混合后進行電擊轉化入大腸桿菌BL21(DE3)進行電擊共轉化,電擊轉化程序:電脈沖為25μF,電壓2.5kV,電阻200Ω,時間4~5ms。pET32a-EG轉化子用100mg/L氨芐青霉素(Amp);pET30b-BGL轉化子用50mg/L卡那霉素(Kan);共轉化的轉化子經同時含有Amp和Kan的雙抗生素LB平板上篩選單菌落,37℃過夜培養后,挑取單菌落進行菌落PCR鑒定。
1.2.3 重組菌株的菌落PCR及高酶活菌株的篩選 將經相應抗生素篩選后的轉化子,分別轉入到另一個新鮮的帶有相應抗性的平板上,37℃過夜培養后,利用T7引物進行菌落PCR鑒定。分別挑取10株鑒定結果為陽性的轉化子進行終濃度1mmol/L IPTG誘導表達,收集菌體用1/15mol/L的PBS重懸,超聲破碎后,用上清經適當稀釋后測酶活。
1.2.4 酶活力測定方法

表1 本實驗所用的菌株與載體Table1 The strains and vectors used in this experiment
表2 本實驗中所用到的引物序列Table2 Sequences of primers used in this experiment
1.2.4.1 內切葡聚糖酶活力測定方法 以1m L 1%的羧甲基纖維素鈉(用1/15mol/L pH6.8磷酸鹽緩沖液配制)為底物,加入0.1m L粗酶液,50℃水浴反應30m in后,加入2.5m L DNS顯色液,沸水浴煮10m in,取出后在流水下迅速冷卻后定容至5.0m L搖勻,將滅活的酶液作為空白對照,其他同以上步驟。在530.0nm波長處測得各管溶液的吸光值[5,10]。
酶活的定義:1m L酶液每分鐘產生1μg還原糖的酶量作為1個酶活單位,用U/m L表示。
1.2.4.2 β-葡聚糖酶活力測定方法 加入0.5m L一定稀釋度的粗酶液,以0.5m L 0.5%的水楊苷為底物,加入0.5m L一定稀釋酶液,于55℃反應20m in,加入1m L DNS試劑,充分混合在沸水中煮沸5min,冷卻后用蒸餾水定容至5m L,在540nm波長處測定其吸光度,以加熱滅活的酶液為對照。根據葡萄糖標準曲線計算酶反應體系釋放的還原糖的量[6,11]。
酶活定義:1m L酶液在1m in內使底物產生1μg還原糖(以葡萄糖計)所需的酶量為1個酶活單位(U)。
1.2.4.3 共表達酶活力測定方法 共表達酶活以內切葡聚糖酶方法進行。
1.2.5 蛋白的分離純化 誘導表達的發酵液經離心收集菌體,按每100mg菌體(濕重)加入1~5m L細菌裂解液,經超聲裂解菌體后,10000r/min,4℃離心3min,收集上清用GE公司生產的預裝Ni+柱,用10倍柱體積濃度為50mmol/L的咪唑進行平衡后,上樣后用15倍柱體積濃度為500mmol/L的咪唑進行洗脫,每1m L收集一管,留樣進行SDS-PAGE分析。
1.2.6 重組酶酶學性質分析 最適反應pH將酶在pH3.0~10的底物中進行測定,pH穩定性分析將酶經pH 3.0~10的緩沖液中30m in后采用標準方法測定酶活力,最適反應溫度將酶液置于30~90℃反應后采用標準方法測定酶活力,熱穩定性的測定將酶液置于30~90℃下保溫30min,采用標準方法測定酶活力,以最高酶活作為100%。
2 結果與分析
2.1 重組質粒的構建
以pUC18-End質粒和pMD19-BGL為模板用高保真酶進行PCR擴增,獲得的片斷大小分別約為1500bp和2800bp,與預計大小一致,克隆連接入表達載體,經抗性篩選和菌落PCR鑒定后,提取質粒采用相應的酶進行雙酶切分析。pET32a-EG質粒,用Bam HⅠ和XhoⅠ酶處理,電泳結果如圖1所示,酶切過后的載體和目的基因片段大小正確分別為1500bp和5600bp,表達載體pET32a-EG構建成功。
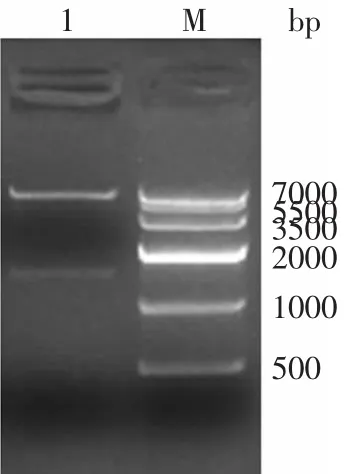
圖1 重組表達載體pET32a-EG的酶切電泳Fig.1 Restriction analysis of recombinant plasmid pET32a-EG
pET30b-BGL,用BglⅡ和XhoⅠ進行單、雙酶切鑒定,電泳檢測結果如圖2所示。
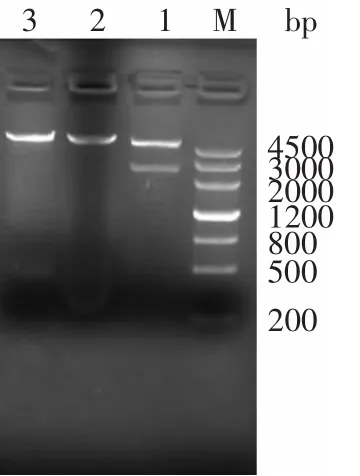
圖2 重組表達載體pET30b-BGL的酶切電泳Fig.2 Restriction analysis of recombinant plasmid pET30b-BGL
重組質粒經BglⅡ和XhoⅠ分別單酶切之后的目的條帶大小與預計大小一致,酶切條帶單一。經BglⅡ和XhoⅠ雙酶切之后,目的基因片段與載體分離,目的基因片段大小和載體大小均正確,確定目的基因和載體連接成功。
2.2 共表達菌株的菌落PCR篩選
將構建好的內切葡聚糖酶基因表達載體和外切葡聚糖酶基因表達載體通過電轉化法轉入BL21菌株中,經(含Amp 100μg/m L和Kan 50μg/m L)雙抗篩選后,得到的陽性轉化子進行菌落PCR鑒定,以T7up和T7down為引物,PCR擴增結果如圖3所示,兩條帶大小分別為1500bp和2800bp。
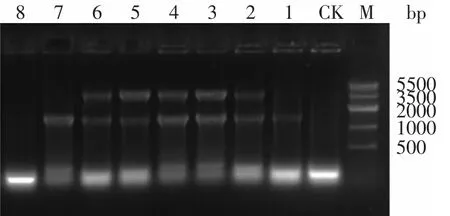
圖3 菌落PCRFig.3 Colony PCR identification
2.3 重組工程菌的IPTG誘導表達篩選
分別從相應抗性的平板上各挑取10個陽性轉化子,經終濃度為1mmol/L IPTG誘導后,測酶活。測得酶活如表3所示,從表中分析結果可知,共轉化菌株酶活最高的是2號,酶活可達到1196.8U/m L,將其命名為pET32a-EG-pET30b-BGL;pET30b-BGL重組轉化菌株酶活最高是5號,酶活可達到219.6U/m L,命名為pET30b-BGL;pET32a-EG重組轉化菌株酶活最高的是10號,酶活可達到826.5U/m L,命名為pET32a-EG。
2.4 蛋白的分離純化SDS-PAGE分析
分別對篩選出三種酶活較高的基因工程菌pET32a-EG-pET30b-BGL、pET30b-BGL、pET32a-EG的細胞裂解物進行SDS-PAGE凝膠電泳分析,如圖4所示,5號和6號都出現一條帶,相對分子量大約為55ku或90ku,7號出現兩條帶,相對分子量大約為55ku和90ku,與理論計算大小相符,結果表明內切葡聚糖酶和β葡萄糖苷酶在大腸桿菌中共同得到高效表達。1號和2號是含空載體大腸桿菌的細胞裂解物中,不存在目的條帶,與預期結果相符。
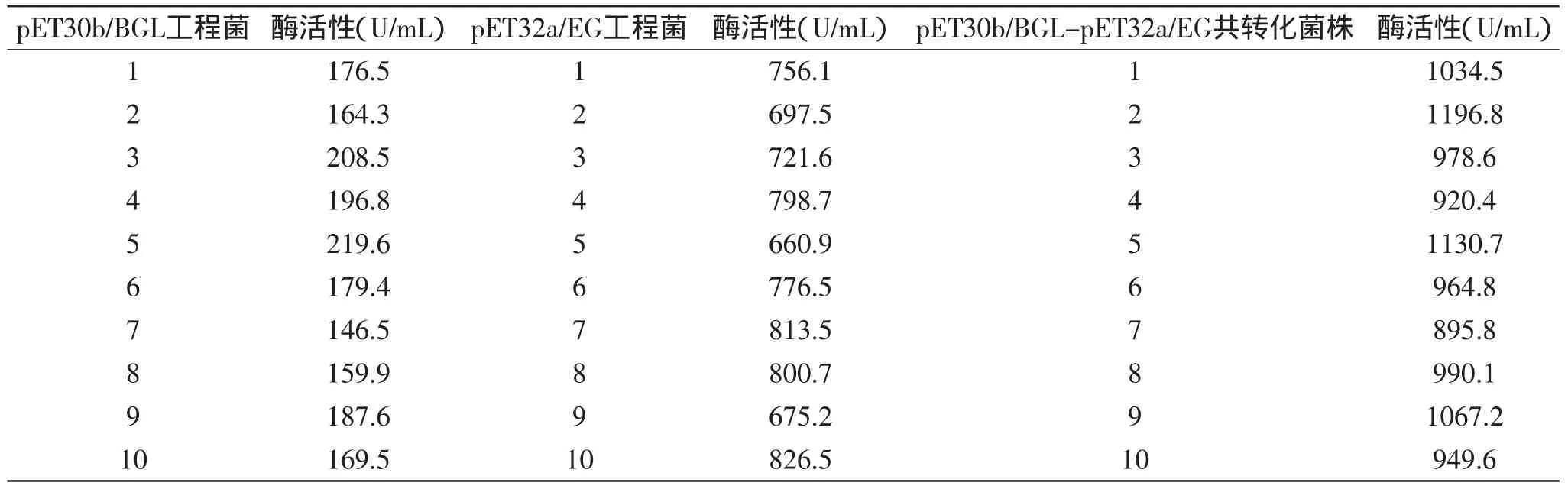
表3 三種酶活性比較Table3 Enzymatic activity comparison of three enzyme

圖4 三種酶表達產物的SDS-PAGE分析Fig.4 SDS-PAGE analysis of expression of three reconstructed enzyme
蛋白質的分離純化方法參照Ni2+柱純化操作方法純化后,得到單一的條帶,條帶大小也與目的蛋白大小預期一致,分別為55ku和90ku,如圖5所示。
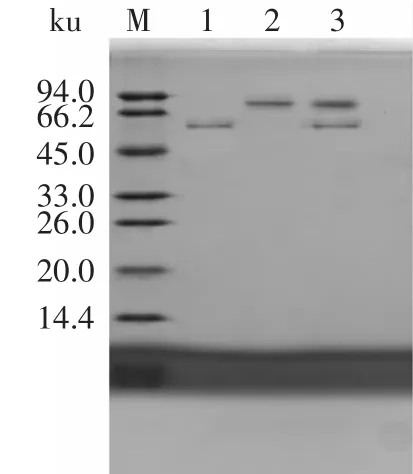
圖5 純化后目的蛋白SDS-PAGEFig.5 SDS-PAGE analysis of purification protein
2.5 重組酶酶學性質分析
2.5.1 重組酶最適反應pH測定 取適當稀釋的純化后的酶液在不同pH條件下與底物共同反應30m in后,測定其酶活,結果如圖6所示,重組共表達酶(pET32a-EG-pET30b-BGL)、重組內切葡聚糖酶(pET32a-EG)和重組β-葡萄糖苷酶(pET30b-BGL)的最適反應pH均為6.0,在pH為6.0的環境下,三種重組酶均能達到最大酶活力。

圖6 三種酶的最適反應pHFig.6 pH profile of the three reconstructed enzyme
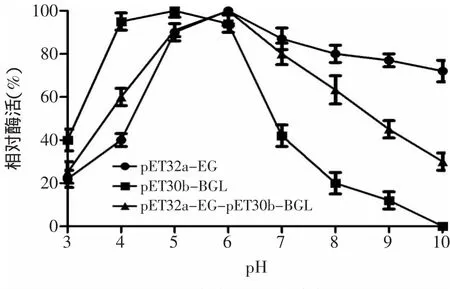
圖7 三種酶的pH穩定性Fig.7 pH stability of the three reconstructed enzyme
2.5.2 重組酶pH穩定性的測定 取適當稀釋的酶液在不同pH下處理后采用標準方法測定其酶活結果如圖7所示,共表達酶(pET32a-EG-pET30b-BGL)在pH 5~7之間較為穩定,能保持最高酶活的80%以上,β-葡萄糖苷酶(pET30b-BGL)在pH偏酸性的環境中較為穩定,在pH4~6的環境中處理后,其殘余酶活力仍能保持最高酶活的80%。內切葡聚糖酶(pET32a-EG)在pH5~9酶活力相對穩定,可以保持在最高酶活的80%以上。
2.5.3 重組酶最適溫度測定 取適當稀釋的酶液不同的溫度下與底物共同反應30m in后,測定其酶活,結果如圖8所示,重組共表達酶(pET32a-EG-pET30b-BGL)、重組內切葡聚糖酶(pET32a-EG)的最適反應溫度均為60℃,重組β-葡萄糖苷酶(pET30b-BGL)的最適反應溫度為50℃,在最適反應pH為6.0的環境下,三種重組酶在各自的最適反應溫度下均能達到最大酶活力。

圖8 三種酶的最適反應溫度Fig.8 Optimal temperature of the three reconstructed enzyme
2.5.4 重組酶溫度穩定性測定 取適當稀釋的酶液在不同的溫度下處理后,采用標準方法測定其酶活,結果如圖9所示,共表達酶共表達酶(pET32a-EG-pET30b-BGL)和內切葡聚糖酶(pET32a-EG)在30~60℃范圍內穩定性良好,β-葡萄糖苷酶(pET30b-BGL)在30~50℃穩定性良好,隨著溫度的逐漸升高,超過50℃,β-葡萄糖苷酶(pET30b-BGL)酶活急劇下降,60℃時酶活僅為最高酶活的10%,溫度高于80℃時共表達酶和內切葡聚糖酶酶活迅速下降。
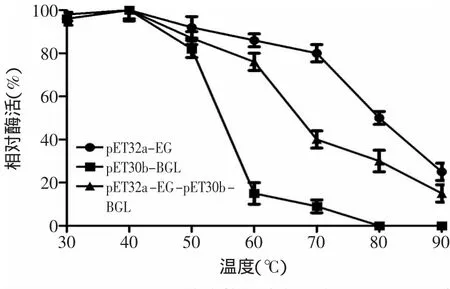
圖9 三種酶熱穩定性分析Fig.9 Heat stability of the three reconstructed enzyme
3 討論
隨著基因克隆技術的飛速發展,許多外源基因在不同的表達載體中進行表達,但是,很多生物學功能并不是由一個基因或一種蛋白來控制和完成的,它需要多個相關基因或蛋白的共同作用[12-14]。比如纖維素酶,就是有三種酶共同作用來分解纖維素的,因此,研究多個外源基因在同一宿主細胞中的共表達具有現實的意義和重要的應用價值。雙質粒共表達系統是將兩個外源基因,分別構建到不同抗性的載體中,然后把兩個載體通過電轉化法整合到大腸桿菌中,實現共表達[8,15-16]。主要分為兩種,一種是復制子相同的不相容性雙質粒,另一種是復制子不同的相容性雙質粒。由于目前較難找到復制子不相同的兩種載體,因此復制子相同的不相容雙質粒系統操作更加簡便,更具有實用性[9,14]。楊巍等[14]利用不相容雙質粒系統,將DNA撕裂因子的2個亞基DFF45和DFF40基因利用不同抗性不相容雙質粒系統在大腸桿菌BL21(DE3)中進行了共表達。范立強等[9]在關于大腸桿菌肉堿脫水酶基因caiB及其輔酶合成因子基因caiE的共表達研究中,對相容和不相容這2種雙質粒系統進行了比較,結果證明了2種相容質粒和不相容質粒都能共存于宿主細胞中,其轉化子的數量相近;同時證明了含相容質粒菌體的存活率的不恒定性,不相容雙質粒系統成功因素主要有有質粒選擇、轉化和培養時間,對于質粒的選擇,既要考慮抗性的不同,還要保證外源基因能高效表達,因此還要為基因工程的下游研究打好基礎[17-20]。
本實驗采用的是不相容雙質粒表達系統,分別構建好pET32a-EG和pET30b-BGL,然后通過電擊轉化法,將構建好的兩種重組質粒轉化到制備好的感受態細胞中,通過氨芐和卡那這兩種抗生素進行雙抗篩選,在雙抗的脅迫下,使其兩種不相容質粒在同一宿主菌中穩定存在,使兩個外源基因在大腸桿菌中首次實現了共同表達,其酶活為1196.8U/m L,比單一酶組分的酶活力要高,通過酶學性質分析顯示共表達酶的最適反應pH為6.0,最適反應溫度60℃,在pH5~7范圍區間內能保持較高的pH穩定性,能夠達到最高酶活的80%以上。在30~60℃范圍能有較高的溫度穩定性,酶活維持在80%以上。這一結果符合纖維素酶的協同學說,內切葡聚糖酶主要作用于纖維素的非結晶區,從還原端或非還原端切下纖維二糖,β-葡聚糖苷酶水解纖維二糖成二個葡萄糖。在本研究中,共表達產生的兩種纖維素酶,內切葡聚糖酶和β-葡聚糖苷酶在對可溶性底物羧甲基纖維素鈉降解過程中,纖維二糖的生成累積,被β-葡聚糖苷酶降解成了葡萄糖,使得纖維二糖對內切葡聚糖酶的一個反抑制作用減弱了,這就使得其能持續穩定地降解[5-7]。從而使酶活力發揮最大作用。這一研究結果將具有重大的經濟價值和工業應用價值,同時為纖維素酶共表達的進一步研究奠定基礎。
4 結論
本研究利用pET32a和pET30b兩個載體的不同抗性的特性,在大腸桿菌中將β-葡萄糖苷酶基因和內切葡聚糖苷酶基因,實現了不相容性共表達,獲得了產兩種酶的工程菌,酶活力可達1196.8U/m L。比單一酶組分的酶活力高,酶學性質分析顯示共表達酶的最適反應pH為6.0,最適反應溫度60℃,在pH 5~7范圍內能保持較高活性,酶活可達最高酶活的80%以上。在30~60℃范圍能有較高的溫度穩定性,酶活維持在80%以上。這一結果將為工業應用的進一步研究提供理論基礎。
[1]宋京城,蔡健.纖維素酶在食品工業中的應用[J].農產品加工·學刊,2010(3):69-71.
[2]張明霞,段長青,張文娜.纖維素酶在食品工業中的應用與展望[J].釀酒科技,2005(4):99-100.
[3]閆訓友,史振霞,張惟廣,等.纖維素酶在食品工業中的應用進展[J].食品工業科技,2004,25(10):140-142.
[4]Escovar-Kousen JM,Wilson D,Irwin D.Integration of computermodelingand initialstudiesofsite-directedmutagenesis to improve cellulase activity on Cel9A from Thermobifida fusca [J].Appl Biochem Biotechnol,2004,113-116:287-297.
[5]高培基.纖維素酶降解機制及纖維素酶分子結構與功能研究進展[J].自然科學進展,2003,13(1):21-29.
[6]Parker A,Maw B,Fedor L.The beta-glucuronidase catalyzed hydrolysis of a glucopyranosiduronamide and a glucopyranoside:evidence for the oxocarbonium ion mechanism for bovine liver beta-glucuronidase[J].Biochem Biophys Res Commun,1981,103(4):1390-1394.
[7]Churilova IV,Maksimov VI,Klesov AA.Cellobiose as a regulator of endoglucanase activity of cellulase complexes.Mechanism of the regulation[J].Biokhimiia,1979,44(11):2100-2102.
[8]龐永奇,賈洪革,方榮祥,等.利用不相容質粒共轉化大腸桿菌對Cre重組酶體內重組活性的可視檢測[J].微生物學報,2005,45(1):125-128.
[9]范立強,袁勤生,吳祥甫.不相容雙質粒共表達大腸桿菌肉堿脫水酶基因caiB及其輔因子合成酶基因caiE[J].中國醫藥工業雜志,2002,33(3):104-108.
[10]吳振芳,陳惠,曾民,等.內切葡聚糖酶基因在畢赤酵母中高效表達研究[J].農業生物技術學報,2009,17(3):529-535.
[11]朱龍寶,湯斌,陶玉貴,等.黑曲霉β-葡萄糖苷酶基因克隆及在畢赤酵母中分泌表達[J].食品與生物技術學報,2012,31(9):973-977.
[12]Sun J,Phillips CM,Anderson CT,et al.Expression and characterization of the Neurospora crassa endoglucanase GH5-1 [J].Protein Expr Purif,2011,75(2):147-154.
[13]Sommer B,Friehs K,Flaschel E.Efficient production of extracellular proteinswith Escherichia coli bymeans ofoptimized coexpression of bacteriocin release proteins[J].J Biotechnol,2010,145(4):350-358.
[14]楊巍,張嵐,盧智剛.利用兩種不相容質粒在大腸桿菌中共表達DFF45和DFF40[J].生物化學與生物物理學報,2001,33:238-242.
[15]孫劍,王健琪,翟朝陽.不相容雙質粒共表達人內皮抑素及簡化人纖溶酶原餅環區5[J].四川大學學報,2006,37(6):839-843.
[16]Ervin SE,Small PA Jr,Gulig PA.Use of incompatible plasmids to control expression of antigen by Salmonella typhimurium and analysis of immunogenicity in mice[J].Microb Pathog,1993,15(2):93-101.
[17]鐘聲,劉丹平,劉素偉,等.血管內皮生長因子121和骨形態蛋白2雙基因共表達重組腺病毒載體的構建及鑒定[J].中國組織工程研究與臨床康復,2011,15(20):3741-3744.
[18]王彥,吳奎,畢玉田,等.FasL和Der p2雙基因共表達真核表達載體的構建及其在樹突狀細胞中的表達[J].重慶醫學,2010,39(20):2697-2703.
[19]Cheng CY,Yu YJ,Yang MT.Coexpression of omega subunit in E.coli is required for themaintenance of enzymatic activity of Xanthomonas campestris pv.campestris RNA polymerase[J].Protein Expr Purif,2010,69(1):91-98.
[20]Wang F,Qu H,Zhang D,etal.Production of1,3-propanediol from glycerolby recombinant E.coli using incompatible plasmids system[J].Mol Biotechnol,2007,37(2):112-119.
