煙臺近海浮游植物的時空變化特征
2013-03-20 01:29:16王妍董志軍劉東艷邸寶平
海洋通報 2013年4期
王妍,董志軍,劉東艷,邸寶平
(1.中國科學院煙臺海岸帶研究所 中國科學院海岸帶環境過程與生態修復重點實驗室(煙臺海岸帶研究所)山東省海岸帶環境過程重點實驗室,山東 煙臺 264003;2.中國科學院大學,北京 100049)
浮游植物是海洋中重要的初級生產者,其種類組成與數量變動直接影響著整個海洋食物網中的物質循環和能量傳遞。近年來,越來越多的研究表明,受近海環境變化的脅迫,浮游植物群落結構正處于不穩定狀(王云龍等,2005; 孫翠慈等,2006;戴明等,2007),赤潮頻發。因此,定期觀測浮游植物的群落結構特征對于了解海灣的生態環境狀態具有重要意義。
煙臺近海曾經是我國煙威漁場的主要海區,目前已成為北方的重要海水養殖基地。近年來,高密度養殖和頻繁的人類活動導致該海域環境質量下降,生態系統出現了不穩定性。春夏季有毒有害赤潮爆發已成為常態,如:1998年8-9月在四十里灣發生了血紅哈卡藻赤潮(吳玉霖等,2001),2004年5月在四十里灣發生夜光藻赤潮(遲守峰,2008),2009年8月在四十里灣發生了赤潮異彎藻赤潮(山東省海洋與漁業廳,2010),對當地海洋生態環境及海洋漁業等造成了極大的危害。然而,煙臺近海浮游植物群落結構的研究主要集中于港口(李偉才 等,2006)、垃圾傾倒區(紀靈 等,2003;劉旭 等,2010) 等局部區域(中國海灣志編纂委員會,1991) 或個別月份(寧璇璇 等,2011),對于該海域全面綜合調查的研究資料積累卻不多,僅蔣金杰等對煙臺四十里灣的浮游植物群落的季節變化開展了研究(蔣金杰等,2011)。本研究于2010年4月-2011年3月期間,在煙臺近海包括四十里灣和套子灣28 個站位開展了為期一年的雙月調查,對浮游植物的群落結構、季節變化與物種演替特征進行了詳細研究,以期為深入了解浮游植物對環境的響應機制以及建立該海域健康生態環境的管理提供基礎資料。
1 材料和方法
1.1 研究站位及采樣方法
煙臺位于山東半島北部,是北黃海沿岸的重要組成部分,具有典型溫帶季節特征。煙臺近岸海域夏季水溫通常為23.3 ℃~27.4 ℃,冬季水溫為2.5 ℃~3.5 ℃,水深多為8~10 m,平均水深約12 m。煙臺近岸海域主要由套子灣和四十里灣兩個海灣組成,以芝罘島為分界線,東到養馬島,西到龍洞咀。沿岸多條中、小型河流入海,其中大沽夾河為最大的河流。本研究分別于2010年4月、6月、8月、10月、12月和2011年3月對煙臺近海共28 個站位進行了6 個航次的生態環境綜合調查(圖1),對浮游植物的群落特征進行了綜合分析。
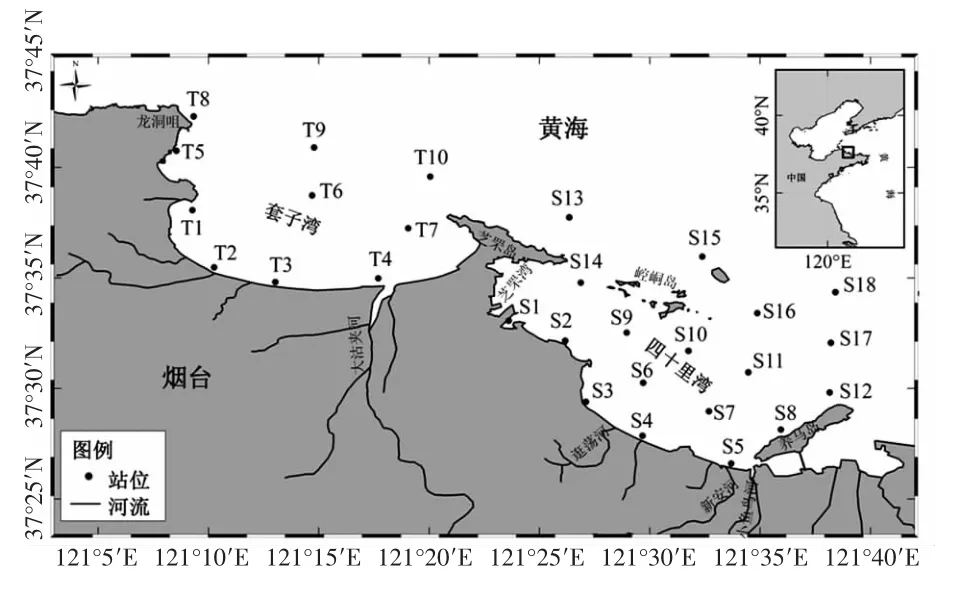
圖1 煙臺近海采樣站位
調查期間,在28 個站位利用Niskin 采水器采集500 mL 表層海水,加入最終濃度為3%~5%福爾馬林溶液固定后,帶回實驗室進行浮游植物的種類組成和數量分析。浮游植物鑒定和計數在Olympus IX 51 型倒置光學顯微鏡下進行。浮游植物計數采用國際上通用的Uterm?hl 方法(Uterm?hl,1958)。樣品充分搖勻后取出10 mL 放入Uterm?hl計數框,靜置24 h,使浮游植物充分沉淀,用于鑒定和計數。浮游植物種類的鑒定參考書為《中國海藻志》、《海藻學概論》、《海藻學》、《中國近海赤潮生物圖譜》、《中國海域常見浮游硅藻圖譜》、《赤潮生物》 (郭玉潔 等,2003; 李偉新 等,1982; 錢樹本 等,2005; 郭皓,2004; 楊世民等,2006;林永水等,2001) 等。
另將600 mL 表層海水用0.45μm 的醋酸纖維濾膜過濾后,用德國Bran+Luebbe AA3 營養鹽自動分析儀測定其營養鹽的濃度,包括無機氮(DIN=NH4-N+NO2-N+NO3-N)、活性磷酸鹽(SRP)、活性硅酸鹽(DRSi)。
1.2 數據處理方法
浮游植物的優勢種以優勢度指數(Y >0.02)判斷,其計算公式為:
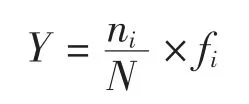
物種多樣性指數采用香農-威弗指數(H′,Shannon-Wiever index)(Shannon et al,1949),其計算公式為:
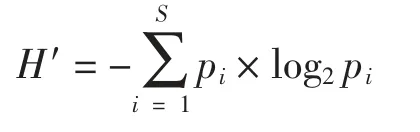
物種均勻度指數(J) (Pielou,1969),其計算公式:
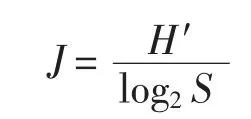
式中,ni為第i 種的總個體數,N 為所有物種的總個體數,fi為第i 種在各站位出現的頻率,S 為樣品中總種類數,Pi為第i 種的個體數與樣品中的總個數的比值。
利用SPSS 17.0 軟件包中的T 檢驗分析四十里灣與套子灣浮游植物細胞豐度的差異性。利用PRIMER 6.0 進行聚類分析(Cluster analysis),對群落結構空間分布進行研究。應用單因子相似性分析ANOSIM(Analysis of Similarities) 檢驗四十里灣和套子灣種類組成的差異顯著性,該分析給出一個統計量global R,其值一般介于0-1:當R=0 時,表示群落結構完全相同;當0<R<0.5 時,表示群落結構差異不明顯;當0.5<R<0.75 時,表示群落之間雖有重疊但能清楚分開;當0.75<R<1 時,表示群落之間有顯著差異;當R=1 時,表示群落完全不同。利用相似性百分比分析SIMPER(Similarity Percentages) 找出造成四十里灣和套子灣浮游植物差異的特征種類(Clarke,1993;Clarke et al,2006)。
2 結果與討論
2.1 營養鹽季節性特征
煙臺近海DIN 全年平均值是12.0 μM,變動范圍是5.3~17.9 μM,10月濃度最高,8月最低,秋季(10月)、冬季(12月) 和初春(3月) DIN 濃度較高。SRP 的全年平均值是0.9 μM,濃度范圍是0.1~2.5 μM,秋季和冬季磷酸鹽水平較高。DRSi 濃度全年平均值是2.0 μM,季節變化趨勢較平穩,變動范圍是1.0~2.7 μM,8月DRSi 濃度最高,6月最低。煙臺近海各月受到不同程度的營養鹽硅限制:6月、10月和3月超過2/3 的站位處于硅限制中;8月約1/2 的站位受到硅限制;4月、12月約有1/3 的站位受到硅限制。
2.2 浮游植物種類組成特征
調查期間,共鑒定記錄浮游植物78 屬187 種,其中硅藻50 屬103 種,甲藻21 屬73 種,褐胞藻5 屬7 種,綠藻1 屬3 種,還包括一個未定類(三深裂醉藻,Ebria tripartite) (表1,見附錄)。在硅藻門中角毛藻屬(Chaetoceros) 的種類最多,共有12 種;甲藻門中原多甲藻屬(Protoperidinium)的種類最多,為25 種。浮游植物生態類型主要以溫帶近岸種和廣布性種為主。
煙臺近海物種豐富度、多樣性指數和均勻度指數呈現出明顯的季節變化與團塊分布特征(圖2,3)。夏季(8月) 物種豐富度最高,為106 種;冬季(12月) 最低,為47 種。夏初(6月) 和秋季(10月) 的物種多樣性指數和均勻度指數較高,夏初(6月) 兩種指數平均值分別為2.33、0.64,秋季(10月) 分別為2.55、0.68;夏末(8月) 多樣性指數和均勻度指數最低,分別為1.23、0.28。同時,煙臺近海的多樣性指數和均勻度指數隨季節變化表現出較高的空間分布差異。春季(3,4月),套子灣多樣性指數和均勻度指數斑塊分布特征顯著,呈現近岸向外逐漸遞增的趨勢,而四十里灣的多樣性指數和均勻度指數表現為中部低,沿岸和灣外高的特征;夏季(8月),四十里灣多樣性指數和均勻度指數灣內、外特征分布顯著,灣外明顯高于灣內,而套子灣灣內、外差異卻不明顯;冬季(12月),套子灣多樣性指數和均勻度指數分布特征是東部高于西部,而四十里灣的低值區位于灣東部。
2.3 浮游植物生物量的時空變化特征
2.3.1 周年變化特征
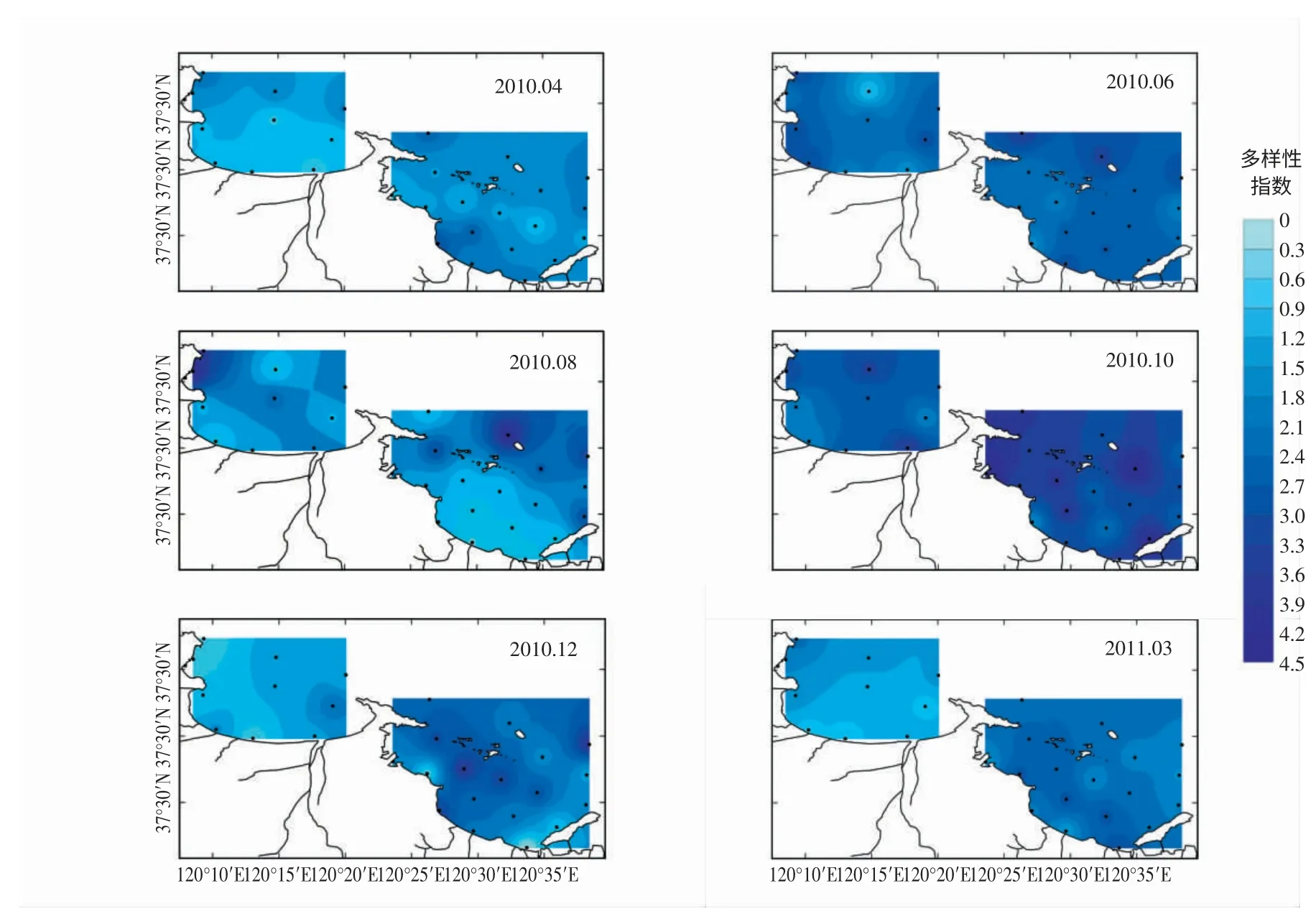
圖2 浮游植物多樣性指數(H′) 的周年變化和平面分布特征
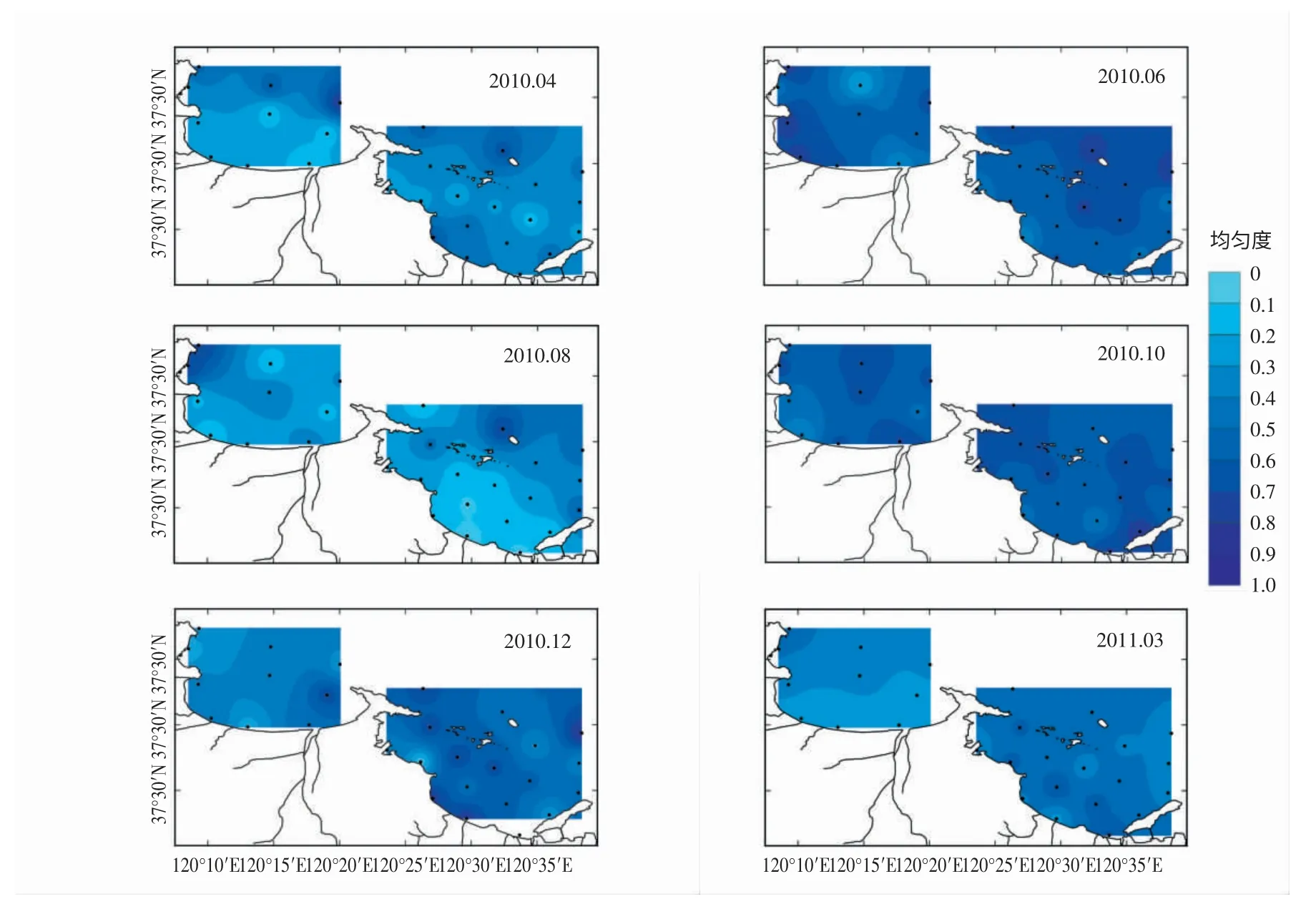
圖3 浮游植物均勻度指數(J) 的周年變化和平面分布特征
與大多數溫帶海域的研究結果一致(俞建鑾等,1993;李廣樓等,2006;楊世民等,2009;李超倫等,2010),煙臺近海浮游植物的細胞豐度具有明顯的季節變化特征,周年變化模式為雙峰型(圖4)。夏末(8月) 細胞豐度達到最高峰,細胞豐度在各站位的平均值是156.7×104cells/L;春季(3,4月) 為次高峰,細胞豐度在各站位的平均值為25.6×104cells/L;冬季(12月) 細胞豐度最低,各站位數量平均值僅為0.6×104cells/L。相關性分析表明(表2),4月浮游植物細胞豐度與SST、SRP及DRSi 的組合相關性最大且顯著,這可能與4月所受到的硅磷限制有關;6月與DIN 相關關系顯著,6月煙臺近海3 種營養鹽的濃度都極低,浮游植物與DIN 顯著相關可能說明了6月溶解性無機氮對細胞豐度的影響程度高于活性磷酸鹽和活性硅酸鹽;8月DIN、SRP 及N:P 的環境變量組合相關性顯著,6月低氮磷的狀態延續到8月,但活性硅酸鹽得到了及時的補充,這可能是細胞豐度與DIN 和SRP 顯著相關的原因。
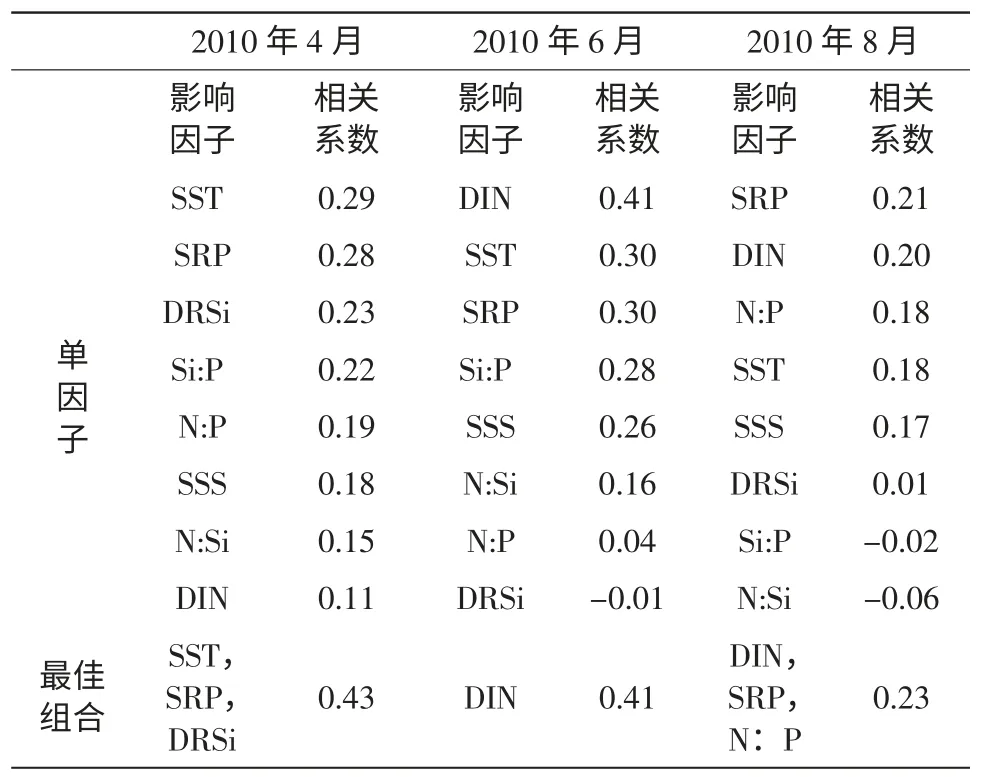
表2 煙臺近海浮游植物豐度與環境因子間的相關性
浮游植物群落結構組成亦呈現出明顯的季節特征(圖5,6)。夏季6月份出現硅藻-甲藻-褐胞藻的聯合優勢特征,在其他調查季節硅藻占據絕對優勢,細胞豐度占到總豐度的80.6 %~99.8 % (圖5)。此外,浮游植物優勢物種的組成具有明顯的季節演替現象(圖6a-f)。冬季(12月) 浮游植物的優勢種單一,主要以具槽帕拉藻(Paralia.sulcata)為主;春季(3,4月) 優勢種以廣溫性的海鏈藻(Thalassiosira sp.)、具槽帕拉藻(P. sulcata) 和角毛藻(Chaetoceros sp.) 為主;夏初(6月) 優勢度較低,形成以錐狀斯克里普藻(Scrippsiella trochoidea)、裸甲藻(Gymnodinium sp.)、赤潮異彎藻(Heterosigma akashiwo)、具槽帕拉藻(P. sulcata)和雙刺原多甲藻(Protoperidinium bipes) 為主的硅藻-甲藻-褐胞藻聯合優勢;夏末(8月) 和秋季(10月) 浮游植物群落結構延續了夏初的特征,出現硅藻-褐胞藻聯合優勢,包括柔弱擬菱形藻(Pseudo-nitzschia delicatissima)、具槽帕拉藻(P.sulcata)、赤潮異彎藻(H.akashiwo)、尖刺擬菱形藻(Pseudo-nitzschia pungens)、海鏈藻(Thalassiosira sp.)、圓篩藻(Coscinodiscus sp.)、中肋骨條藻(Skeletonema costatum)。
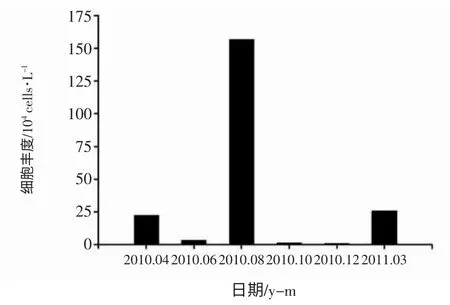
圖4 浮游植物細胞豐度的周年變化
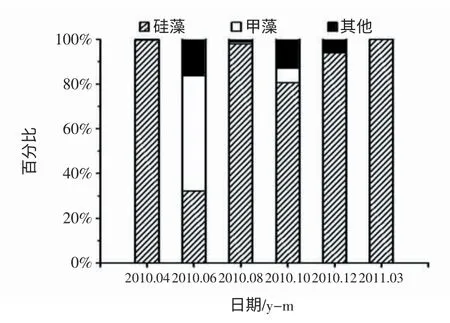
圖5 浮游植物物種組成周年變化
2.3.2 平面分布特征
研究表明,煙臺近海浮游植物生物量的平面分布特征隨著季節的變化而呈現出不同的特征,在浮游植物生物量出現高峰的春季(3,4月) 和夏季(8月),灣內、外平面分布特征顯著,而在生物量低峰期的秋季(10月) 和冬季(12月),灣內、外平面分布特征不顯著(圖7)。以往的研究表明,在近岸海灣內,浮游植物的平面分布可以顯著受到海灣流場與營養鹽濃度分布的影響,其中風海流在近岸表層流場的形成中具有重要作用(管秉賢,1962;張瑞安等,1990; 孫洪亮等,2001; 李占海等,2003;萬修全等,2004)。煙臺近海的溫帶氣候特征具有夏季多東南風、冬季多西北風的特點,而四十里灣與套子灣的位置春季(3,4月)受西北風對流場向岸堆積影響,并且春季冰雪、河流融化后,陸源營養鹽大量注入近岸,故浮游植物生物量的高值區多出現在灣內。在夏季,降雨增多,河流徑流量增大,陸源營養鹽易隨雨水及徑流流入灣內,使得浮游植物生物量高值區多出現在灣內。
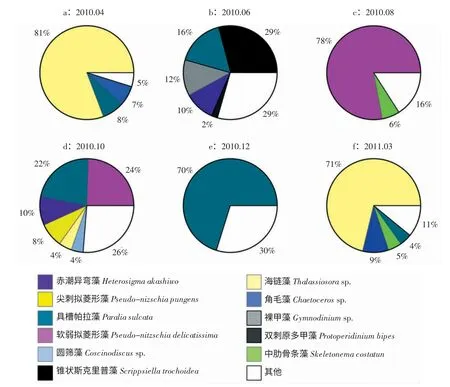
圖6 煙臺近海優勢種組成的周年變化
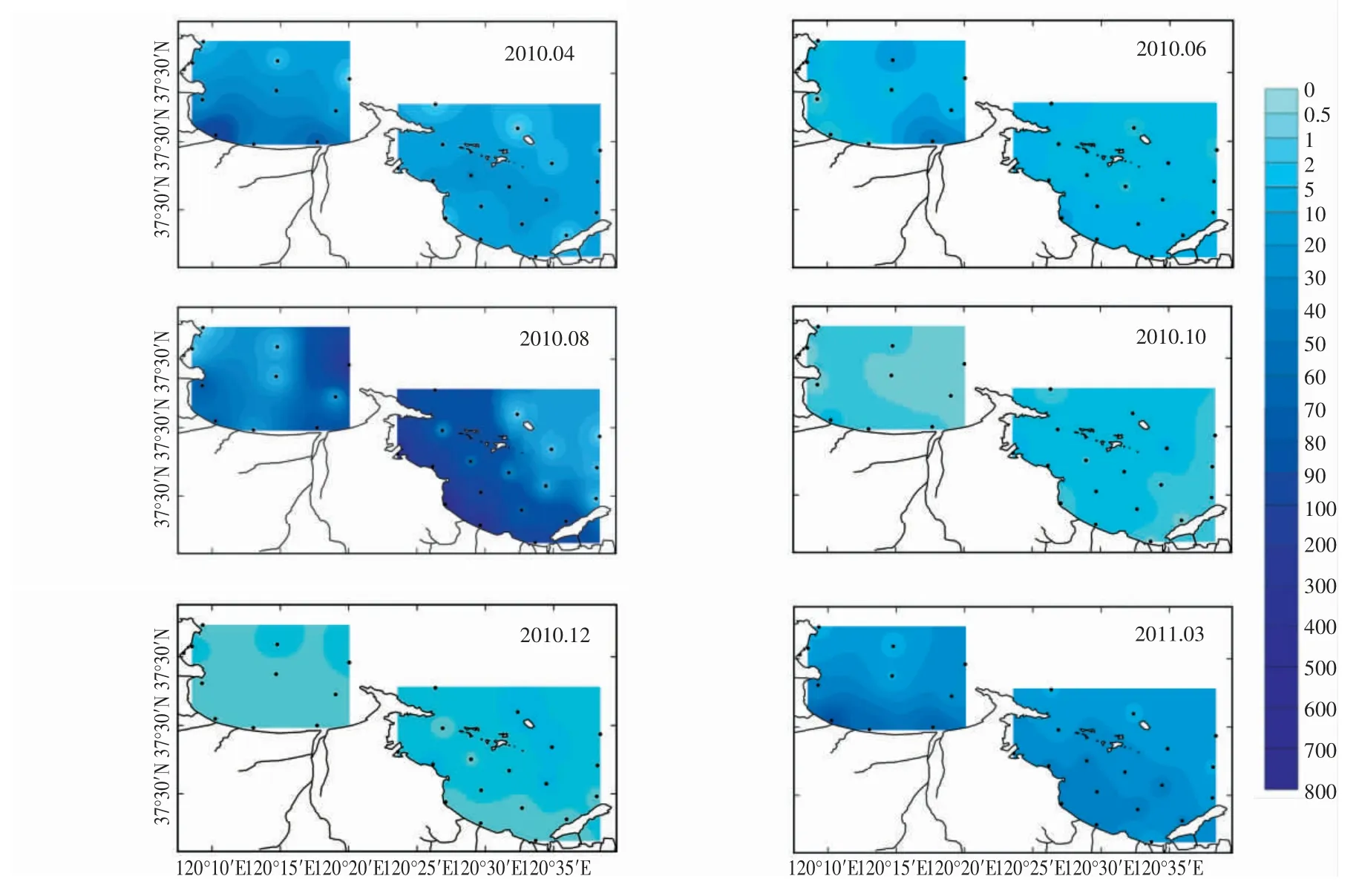
圖7 浮游植物細胞豐度的周年變化和平面分布特征
此外,四十里灣和套子灣的浮游植物細胞豐度的平面分布特征也存在季節差異,如6月和8月,在套子灣,浮游植物生物量出現灣東側高于灣西側的平面分布特征,而四十里灣的這一特征并不顯著,浮游植物生物量高值區仍在灣內側邊緣(圖7)。這可能是由四十里灣和套子灣地理條件及人類活動強度差異所造成的。套子灣為開闊式海灣,夏季東南風形成的風海流影響營養鹽濃度的分布,進而影響浮游植物的平面分布;而四十里灣為半封閉海灣,由于崆峒島和芝罘島的阻隔作用,導致灣內、外水交換不如套子灣通暢,造成營養鹽易在灣內堆積的動力學特征,進而影響浮游植物的平面特征。另外,四十里灣與套子灣受人類活動影響的強度也有所不同,四十里灣受到港運、旅游、貝類養殖和污水排放的多重人類活動影響(李偉才 等,2006; 中國海灣志編纂委員會,1991; 趙衛紅等,2000),而套子灣則主要受旅游和海帶養殖的影響(劉風非,2008),兩灣的污染程度不同,這也可能是造成兩灣浮游植物空間分布特征不同的原因。
2.4 四十里灣和套子灣的浮游植物群落比較
四十里灣和套子灣各調查月及整個調查期的細胞豐度的T 檢驗結果表明,秋季(10月) 和冬季(12月),四十里灣與套子灣細胞豐度存在顯著差異(P<0.05),其他季節差異不顯著,整個調查期間,兩灣生物量并不存在顯著差異(P=0.44>0.05)。

圖8 浮游植物群落結構聚類分析結果
此外,四十里灣和套子灣浮游植物的群落結構在各季節均存在差異。例如,2010年10月,四十里灣的優勢種是柔弱擬菱形藻(P. delicatissima)(29%)、具槽帕拉藻(P.sulcata) (21%) 和尖刺擬菱形藻(P.pungens) (11%),而套子灣的優勢種為具槽帕拉藻(P.sulcata) (28%)、赤潮異彎藻(H. akashiwo)(26 %)和海鏈藻(Thalassiosira sp)(10%)。經聚類分析,28 個調查站位的浮游植物群落按40%~60%的群落相似性劃分為4-7 個聚類組(圖8)。浮游植物物種組成存在明顯的團塊分布特征,除秋季(10月) 外,其他季節套子灣和四十里灣大部分站位處于不同聚類組。為了驗證兩灣浮游植物群落結構差異的顯著性,將四十里灣和套子灣各月浮游植物數據進行ANOSIM 檢驗,并利用SIMPER 分析找出對四十里灣和套子灣浮游植物群落結構差異性重要貢獻的物種。結果表明:早春(3月) 四十里灣和套子灣浮游植物群落結構具有顯著差異(global R=0.867,P=0.1%<0.05),兩灣的造成差異的物種有海鏈藻(Thalassiosira sp.)、角毛藻(Chaetoceros sp.)、丹麥細柱藻(Leptocylindrus danicus)、諾氏海鏈藻(Thalassiosira nordenskioeldii) 等;夏初(6月) 兩灣浮游植物群落雖存在重疊但可以清楚分開(global R=0.618,P=0.1 %<0.05;其他月份兩灣浮游植物差異不明顯(global R<0.5)。
3 結論
根據對煙臺近海2010年4月到2011年3月6個航次28 個站位浮游植物群落特征的調查結果分析發現:
(1) 煙臺近海浮游植物群落結構呈現出明顯的溫帶海域區系特征:組成主要以硅藻和甲藻為主,其中硅藻無論在種類還是細胞數量上都占絕對優勢。浮游植物生態類型以溫帶近岸種和廣布種為主。多樣性指數(H′) 和均勻度指數(J) 呈現明顯的季節變化,6月和10月最高,8月最低。生物量季節變化呈雙峰型,最高峰出現在夏季(8月),次高峰出現在春季(3,4月)。優勢種周年演替明顯,夏季(6月),浮游植物主要以甲藻-褐胞藻-硅藻聯合占優為主,在其他調查季節,硅藻占據絕對優勢地位。
(2) 煙臺近海浮游植物空間分布特征隨著季節變化呈現出不同的特征。春季和夏季,灣外細胞豐度普遍低于灣內;秋季和冬季,灣內、外平面分布特征不顯著。海灣流場與營養鹽濃度分布可能是造成灣內和灣外差異的主要原因。相關性分析表明,4月,煙臺近海SST、SRP 和DRSi 組合最能解釋浮游植物的分布特征;6月,DIN 與細胞豐度顯著相關;8月,DIN、SRP、N:P 比的組合是影響浮游植物豐度的最佳解釋變量組合。四十里灣與套子灣浮游植物細胞豐度的空間分布特征也有所差異,地理條件和人類活動強度的不同是造成兩灣細胞豐度空間特征差異的主要原因。秋季(10月) 和冬季(12月) 四十里灣與套子灣細胞豐度差異顯著,但整個調查期間兩灣細胞豐度差異不顯著。春初(3月),四十里灣與套子灣浮游植物群落結構差異顯著,其他調查季節兩灣浮游植物并無顯著差異。
Clarke K R,1993. Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology,18(1):117-143.
Clarke K R,Gorley R N,2006. PRIMER v6:User Manual/Tutorial.Plymouth,PRIMER-E Ltd.
Pielou E C,1969.An introduction to mathematical ecology.New York:Wiley-Interscience.
Shannon C,Weaver W,1949.The mathematical theory of communication.Chicago:University of Illinois.
Uterm?hl H,1958. Zur Vervollkommnung der quantitativen Phytoplankton Methodik. Mitteilungen der Internationalen Vereinigung für theoretische und angewandte Limnologie,(9):1-38.
遲守峰,2008.煙臺市四十里灣海域赤潮成因分析及預防對策.齊魯漁業,25(9):55-57.
戴明,李純厚,張漢華,等,2007.海南島以南海域浮游植物群落特征研究.生物多樣性,15(1):23-30.
管秉賢,1962.有關我國近海海流研究的若干問題.海洋與湖沼,4(3-4):121-141.郭皓,2004.中國近海赤潮生物圖譜.北京:海洋出版社.郭玉潔,錢樹本,2003.中國海藻志.北京:科學出版社.
紀靈,王榮純,劉昌文,等,2003.煙臺海洋傾倒區環境監測及對比評價.海洋通報,22(2):53-59.
蔣金杰,劉東艷,邸寶平,等,2011.煙臺四十里灣浮游植物群落的季節變化及其對環境的指示意義.海洋學報,33(6):151-164.
李超倫,張永山,孫松,等,2010.桑溝灣浮游植物種類組成、數量分布及其季節變化.漁業科學進展,31(4):1-8.
李廣樓,陳碧娟,崔毅,等,2006.萊州灣浮游植物的生態特征.中國水產科學,13(2):292-299.
李偉才,孫軍,宋書群,等,2006.煙臺港和鄰近錨地及其入境船舶壓艙水中的浮游植物.海洋湖沼通報,(4):70-77.
李偉新,朱仲嘉,劉鳳賢,1982.海藻學概論.上海:上海科學技術出版社.
李占海,柯賢坤,王倩,等,2003.瓊州海峽水沙輸運特征研究.地理研究,22(2):151-159.
林永水,周近明,何建宗,2001.赤潮生物.北京:科學出版社.
劉風非,2008. 套子灣海洋環境現狀淺析. 海洋開發與管理,25(6):96-98.
劉旭,劉艷,趙瑞亮,等,2010.煙臺海洋傾倒區生物群落結構現狀及動態變化分析.海洋通報,29(4):396-401.
寧璇璇,紀靈,王剛,等,2011.煙臺套子灣海域秋季浮游植物群落的初步研究.海洋通報,30(4):425-429.
錢樹本,劉東艷,孫軍,2005.海藻學.青島:中國海洋大學出版社.
山東省海洋與漁業廳,2010.2009年山東省海洋環境質量公報.
孫翠慈,王友紹,孫松,2006.大亞灣浮游植物群落特征.生態學報,26(12):3948-3958.
孫洪亮,黃衛民,2001.廣西近海潮汐和海流的觀測分析與數值研究——Ⅱ.數值研究.黃渤海海洋,19(4):12-21.
萬修全,鮑獻文,吳德星,等,2004.渤海夏季潮致-風生-熱鹽環流的數值診斷計算.海洋與湖沼,35(1):41-47.
王云龍,袁騏,沈新強,2005.長江口及鄰近水域春季浮游植物的生態特征.中國水產科學,12(3):300-306.
吳玉霖,周成旭,張永山,等,2001.煙臺四十里灣海域紅色裸甲藻赤潮發展過程及其成因.海洋與湖沼,32(2):159-167.
楊世民,董樹剛,2006.中國海域常見浮游硅藻圖譜.青島:中國海洋大學出版社.
楊世民,董樹剛,竇明武,等,2009.2004-2005年青島前海定點54 周次浮游植物群落結構特征.海洋科學進展,27(4):523-536.
俞建鑾,李瑞香,1993.渤海、黃海浮游植物生態的研究.黃渤海海洋,11(3):52-59.
張瑞安,董以芝,1990.煙臺排污海域的自然環境條件分析和污染物輸運路經研究.海岸工程,9(2):35-44.
趙衛紅,焦念志,趙增霞,2000.煙臺四十里灣養殖水域營養鹽的分布及動態變化.海洋科學,24(4):31-34.
中國海灣志編纂委員會,1991.中國海灣志-第三分冊:山東半島北部和東部海灣.北京:海洋出版社.

表1 調查期間浮游植物的物種組成
附錄

續表1 調查期間浮游植物的物種組成

續表1 調查期間浮游植物的物種組成

續表1 調查期間浮游植物的物種組成
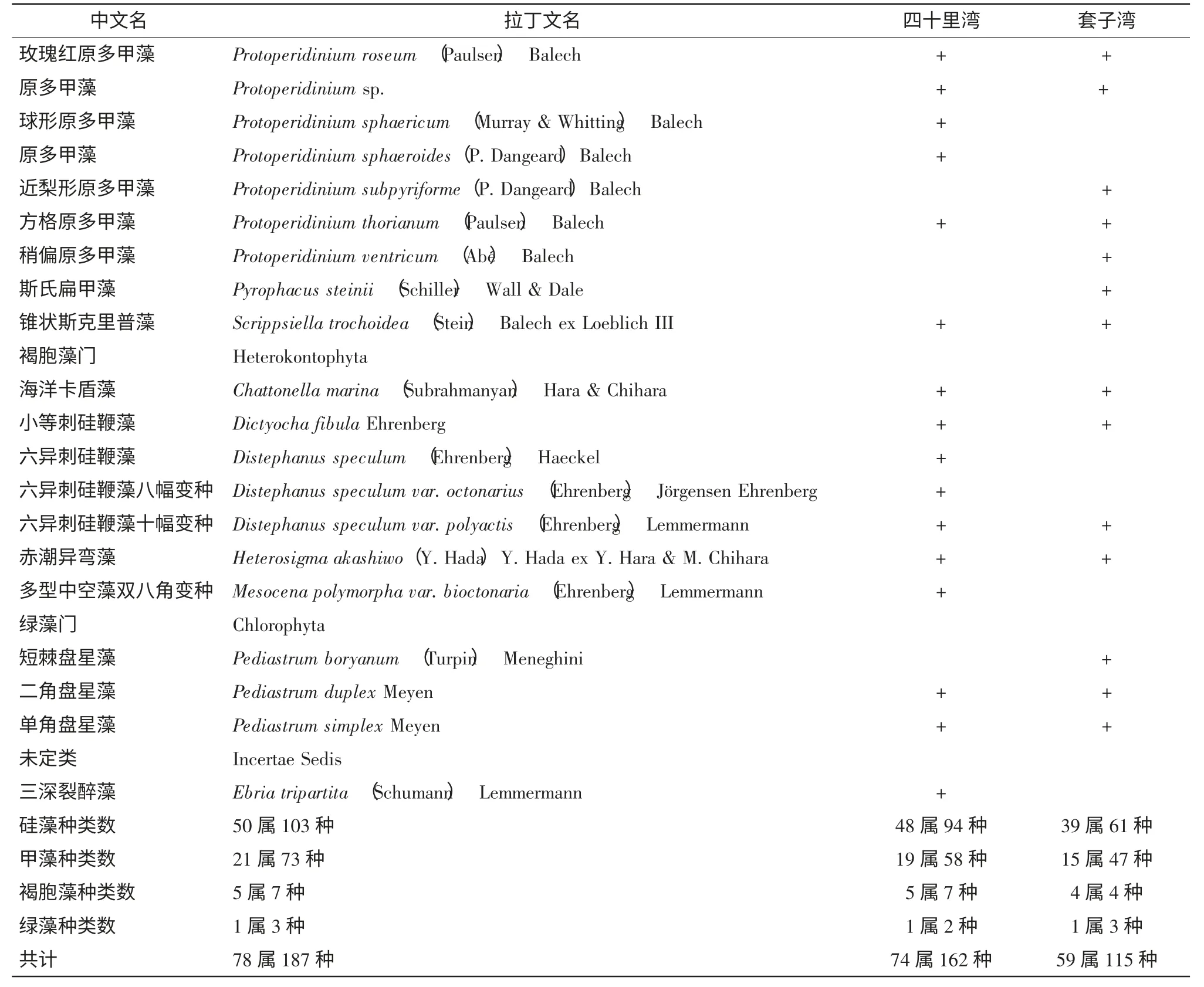
續表1 調查期間浮游植物的物種組成
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
音樂探索(2022年2期)2022-05-30 21:01:37
瘋狂英語·新策略(2019年10期)2019-12-13 08:43:28
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
當代陜西(2019年10期)2019-06-03 10:12:04
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
數學小靈通·3-4年級(2017年9期)2017-10-13 08:10:54
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
