不同糖源及糖水平對大菱鲆糖代謝酶活性的影響
2013-07-24 11:29:34苗惠君苗淑彥陳塵悅李張文兵麥康森
水生生物學報 2013年3期
關鍵詞:水平
聶 琴 苗惠君 苗淑彥 陳塵悅李 靜 張文兵 麥康森
(1.中國海洋大學水產動物營養與飼料農業部重點實驗室, 海水養殖教育部重點實驗室, 青島 266003;2.中國石油大學(華東)生物技術與生物工程中心, 青島 266580)
水產飼料中適當提高糖水平可以起到節約蛋白質、降低飼料成本、減少氨和氮的排泄、減輕水體污染的作用[1,2]。與大多數哺乳動物是以葡萄糖為主要的能量來源不同, 魚類對飼料中糖的利用率較差,特別是一些肉食性魚類, 飼喂富含糖的飼料后會出現持久的高血糖現象[3]。雖然研究已證明魚體內存在利用糖的一系列酶和代謝途徑[4], 但魚類對糖利用能力低下的機制尚不完全清楚。糖分子結構是影響魚類對飼料中糖的利用的因素之一[1,5—7], 一般而言, 魚類對多糖和寡糖的利用要好于單糖和二糖[8—14]。可能原因是單糖易于消化吸收, 促使血糖含量迅速升高, 而魚體內的糖代謝酶活力尚未充分增加, 導致魚體對糖的分解和轉化滯后, 降低了魚體對糖的利用[15]。但在大鱗大麻哈魚(Onchorhynchus tshawytscha)、虹鱒(Oncorhynchus mykiss)、白鱘(Acipenser transmontanus)、草魚(Ctenopharyngodon idella)和金頭鯛(Sparus aurata)中的研究結果表明,這些魚類對葡萄糖或麥芽糖的利用能力好于糊精或淀粉[16—21]。不同魚類對飼料中糖的利用能力也存在差異, 研究證明金頭鯛攝食添加 20%糊化淀粉的飼料后, 其生長不受影響; 當淀粉水平進一步增加時, 則生長減緩、攝食率下降[22]。在飼料中添加10%或 20%的生淀粉或糊化淀粉時, 歐洲海鱸(Dicentrarchus labrax)的生長和飼料利用率與不含糖對照組均無顯著差異[23]。在印度野鯪(Labeo rohita)幼魚飼料中添加 43%的淀粉(生淀粉∶糊化淀粉 = 80∶20)時, 仍然有顯著促進生長和提高蛋白質利用率的效果[24]。一般認為, 草食性和雜食性魚類比肉食性魚類對糖的利用能力強, 淡水、溫水性魚類比海水、冷水性魚類利用糖的能力強, 海水、冷水性魚類飼料的適宜糖水平應低于20%[3]。
大菱鲆(Scophthalmus maximusL.)屬鲆科(Bothidae), 菱鲆屬(Scophthalmus), 原產于歐洲北海、波羅的海、地中海等海域, 屬冷水性魚類。我國從1992年引進至今, 已形成一定規模的大菱鲆養殖, 尤其是在北方的養殖, 創造了巨大的經濟效益和社會效益[25]。大菱鲆攝食習性為肉食性。迄今, 關于大菱鲆配合飼料中糖適宜添加量的報道較少, 且由于實驗用大菱鲆的生長階段、實驗方法的不同,結果存在差異。李曉寧等[26]研究表明大菱鲆幼魚[(4.30 ± 0.01) g]飼料中糖的適宜含量為15.0%或更低, 而中魚[(89.57 ± 0.32) g]和大魚[(208.30 ± 0.29)g]階段飼料中糖含量可達25.5%。但馬愛軍等[6]認為大菱鲆幼魚(27 g)對糖的需求量僅為4%。上述研究都未從糖代謝酶的角度進一步探討大菱鲆的糖代謝,也沒有系統研究糖源和糖水平的交互作用對大菱鲆糖代謝的影響。本研究擬通過在飼料中添加不同糖源及糖水平, 分析兩者對大菱鲆糖代謝酶活性的影響, 為探討大菱鲆幼魚對不同糖源的利用差異, 為確定其飼料糖的適宜添加量提供基礎數據。
1 材料與方法
1.1 飼料配方與制作
本研究設計為3 × 4的雙因素實驗, 即3種糖源(葡萄糖、蔗糖和糊精)和 4個糖水平(0、5%、15%和28%)。以白魚粉、酪蛋白、明膠為蛋白源, 魚油、大豆卵磷脂為主要脂肪源配制等氮等能的實驗飼料。其中, 一種為不添加糖的飼料(Diet 1), 其他飼料組分別添加5%、15%、28%的葡萄糖(Diet 2、Diet 3、Diet 4)、蔗糖(Diet 5、Diet 6、Diet 7)、糊精(Diet 8、Diet 9、Diet 10), 共10種實驗飼料。飼料原料先粉碎, 并過80目篩。將粉碎好的原料按飼料配方逐級放大混合, 再加入大豆卵磷脂和魚油, 手工搓油,并在高效混合機(0.5-V型, 上海天祥健臺制藥機械有限公司)中混勻后, 加入30%左右的蒸餾水揉成軟面團, 用制粒機(EL-260, 山東威海友誼機械廠)擠壓出直徑為1.5 mm的顆粒飼料。飼料在恒溫50℃下干燥 12h, 烘干后存于?20℃的冰柜中待用。飼料配方及成分分析(表1)。
1.2 養殖過程與管理
養殖實驗在中國海洋大學鰲山衛實驗基地流水式養殖系統中進行, 實驗用大菱鲆為青島膠南市古鎮營村養殖廠人工培育的同一批苗種。在正式實驗前, 用商品飼料(青島七好生物科技有限公司)飽食馴養 2周, 以適應養殖環境。饑餓 24h后分組, 平均初始體重為(8.06 ± 0.08) g, 每個玻璃鋼桶(500 L)放養28尾大菱鲆, 每個飼料處理設 3個重復。養殖實驗持續9周, 每天07: 00和18: 00分別進行1次飽食投喂。實驗所用海水經沉淀、沙濾后進入養殖系統, 實驗期間水溫(19 ± 1), pH℃為 7.7 ± 0.1, 鹽度(25.2 ± 1.0)‰, 溶氧含量 ≥ 7.0 mg/L。
1.3 樣品收集與分析
在實驗結束時, 實驗魚饑餓 24h, 每桶隨機選取 3尾, 解剖分離出肝臟, 液氮速凍后保存樣品于?80℃冰箱用于酶活性分析。
飼料常規成分測定: 水分采用恒溫干燥法(105), ℃粗蛋白用凱氏定氮法(N × 6.25), 粗脂肪用索氏抽提法, 灰分用馬弗爐灰化法(550), ℃總能用氧彈儀(Parr 6100, USA)測定。實驗飼料中糖含量以飼料中可利用糖含量計量, 采用蒽酮比色法測定[27]。
大菱鲆肝臟己糖激酶(HK)和丙酮酸激酶(PK)采用試劑盒測定(南京建成生物工程研究所)。葡萄糖激酶(GK)、磷酸果糖激酶(PFK)、磷酸烯醇式丙酮酸羧激酶(PEPCK)、1, 6 - 二磷酸果糖酶(FBPase)的活性采用Polakof,et al.[28]方法: 取0.5 g肝臟, 在4℃下, 加入10倍緩沖液勻漿, 緩沖液參照文獻 [28]配制。勻漿液于10000 r/min離心30min后, 取上清, 在30℃、用酶標儀(Thermo Scientific Multiskan Spectrum, USA)340 nm波長下完成測定。肝臟勻漿上清液蛋白質含量采用 Bradford法測定, 以牛血清蛋白為標準蛋白[29]。酶的活性表示為: 在 30℃條件下,每毫克蛋白每分鐘水解 1 μmol 底物為 1個活力單位U。
1.4 計算及統計方法
數據用平均值±標準誤(X±SE)表示, 采用SPSS 17.0 分析軟件對實驗數據進行雙因素方差分析(Two-way ANOVA)。當差異顯著(P< 0.05)時, 用Duncan 檢驗進行多重比較分析[30]。
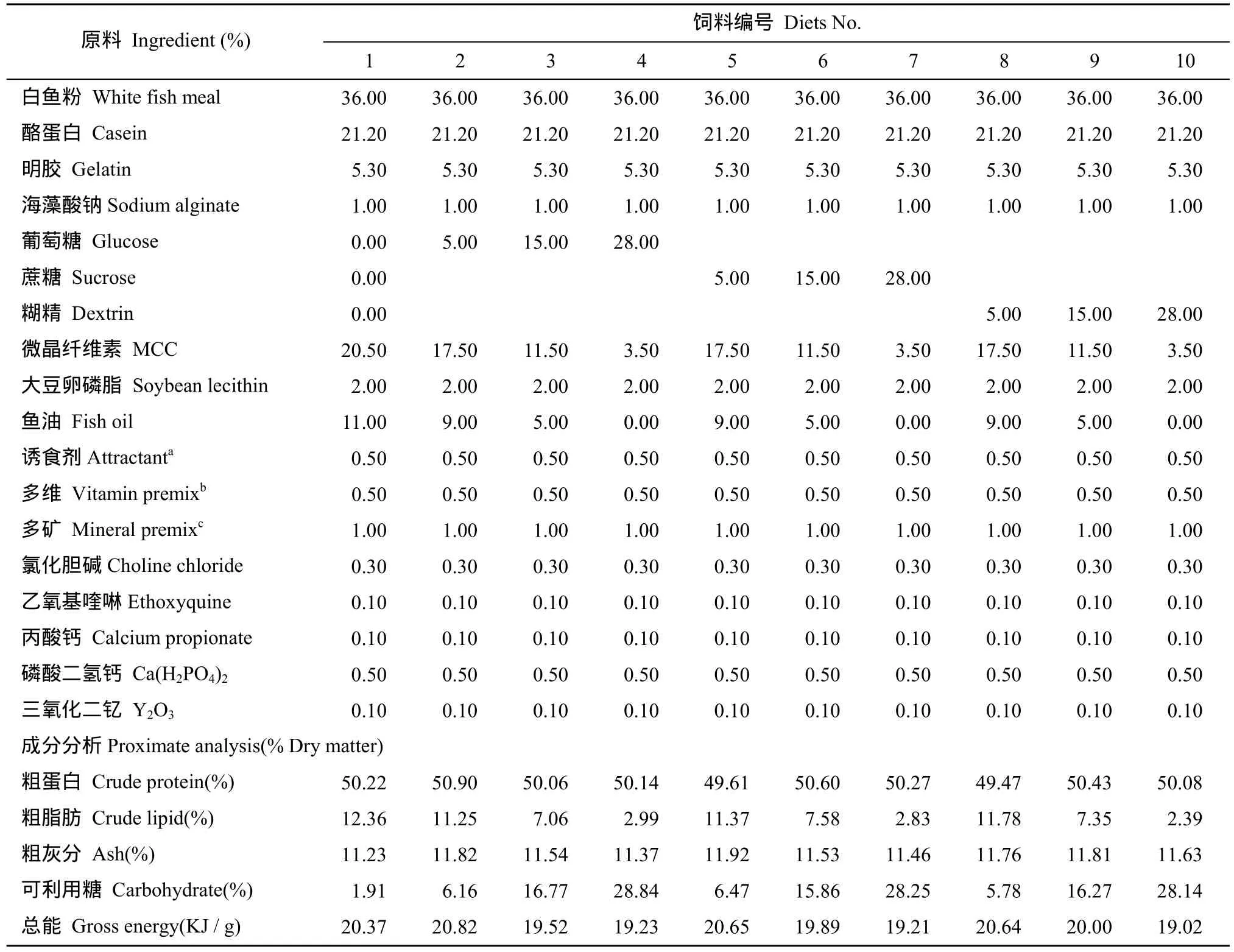
表1 實驗飼料配方及營養組成Tab.1 Formulation and proximate composition of the experimental diets
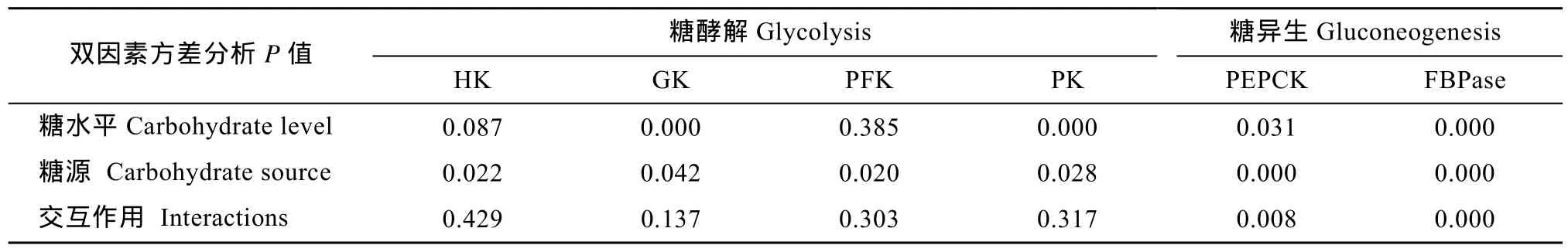
表2 不同糖源及糖水平對大菱鲆肝臟糖酵解和糖異生酶活性的影響Tab.2 Effects of dietary carbohydrate sources and levels on the activities of glycolytic and gluconeogenetic enzymes in the liver of turbot
2 結果
2.1 不同糖源及糖水平對大菱鲆肝臟糖酵解酶活性的影響
大菱鲆肝臟中HK活性不受飼料中糖水平的顯著影響(P> 0.05, 表2), 但不同糖源對HK活性的影響顯著(P< 0.05)。當飼料中糖添加量為28%時, 葡萄糖組HK活性顯著高于糊精組, 蔗糖組HK活性與葡糖糖組和糊精組之間均沒有顯著差異(圖 1A); 在其他糖水平下, 糖源對HK活性的影響均不顯著。
大菱鲆肝臟中GK活性受飼料中糖源及水平的顯著影響(P< 0.05), 但糖源和糖水平的交互作用不顯著(P> 0.05)。在葡萄糖組中, 和對照組相比, GK活性隨葡萄糖水平的升高呈增加趨勢, 當葡萄糖添加量為15%和28%時GK活性都顯著高于對照組(P<0.05)。在蔗糖組中, 蔗糖含量的增加促進GK活性,但僅在添加量為 28%時 GK活性顯著高于對照組(P<0.05)。在糊精組中, 糊精添加量為15%時GK活性顯著高于對照組和添加量為 5%組(P<0.05), 但與添加量為28%組無顯著差異(圖1B)。當飼料中糖添加量為 28%時, 葡萄糖組和蔗糖組 GK活性顯著高于糊精組(圖 1C); 在其他糖水平下, 糖源對 GK活性的影響均不顯著。
大菱鲆肝臟中PFK活性沒有受到飼料中糖水平的顯著影響(P>0.05), 但不同糖源對 PFK活性的影響顯著(P<0.05)。當飼料中糖添加量為28%時, 蔗糖組PFK活性顯著高于糊精組, 葡萄糖組PFK活性與糊精組和蔗糖組之間無差異(圖 1D); 在其他糖水平下, 糖源對PFK活性的影響均不顯著。
大菱鲆肝臟中 PK活性受飼料中糖源及糖水平的顯著影響(P<0.05), 但糖源和糖水平的交互作用不顯著(P>0.05)。在葡萄糖組中, 當葡萄糖添加量為15%時, PK活性為最高(40.44 ± 3.47) mU/mg且顯著高于對照組(P<0.05)。在糊精組中, PK活性呈先升高后降低趨勢, 當糊精添加量為5%時, PK活性顯著高于對照組和含糊精 28%組(P<0.05), 但與含糊精15%組無顯著差異。在蔗糖組中, PK活性不受蔗糖水平的影響(P>0.05)(圖 1E)。當飼料中糖添加量為15%時, PK活性在葡萄糖組顯著高于蔗糖組(P<0.05), 但與糊精組無差異(圖 1F); 在其他糖水平下,糖源對PK活性的影響均不顯著。
2.2 不同糖源及糖水平對大菱鲆肝臟糖異生酶活性的影響
飼料中糖源及糖水平均顯著影響大菱鲆肝臟中PEPCK的活性(P<0.05), 且交互作用顯著(P<0.05)。在糊精組中, PEPCK呈先顯著降低后升高再降低的波動趨勢, 對照組(28.23 ± 1.17) mU/mg和含糊精15%組(23.53 ± 3.81) mU/mg的PEPCK顯著高于含糊精5%組(11.75 ± 1.13) mU/mg(P<0.05), 同時對照組的PEPCK活性也顯著高于含糊精28%組(16.82 ±2.35) mU/mg(P<0.05), 而含糊精 15%和 28%組PEPCK活性無顯著差異; 飼料葡萄糖或蔗糖水平都不影響 PEPCK的活性(圖 2A)。當飼料中糖添加量為5%時, 葡萄糖組和蔗糖組PEPCK活性顯著高于糊精組(P<0.05); 當飼料中糖添加量為28%時, 蔗糖組 PEPCK活性顯著高于葡萄糖組和糊精組(P<0.05); 在其他糖水平下, 糖源對PEPCK活性無顯著影響(圖 2B)。
飼料中糖源及糖水平均顯著影響大菱鲆肝臟中FBPase的活性(P<0.05), 且交互作用顯著(P<0.05)。在葡萄糖組中, FBPase活性先增加后降低, 當葡萄糖添加量為5%時, FBPase活性顯著高于其他組(P<0.05); 在糊精組中, 對照組的FBPase活性顯著高于其他 3組(P<0.05); 在蔗糖組中, 蔗糖水平不影響FBPase活性(圖2C)。當飼料中糖添加量為5%時, 葡萄糖組的 FBPase活性顯著高于蔗糖和糊精組(P<0.05); 當飼料中糖添加量為 15%時, 葡萄糖組的FBPase活性顯著高于糊精組(P<0.05), 但與蔗糖組無差異; 在其他糖水平下, 糖源對FBPase活性無顯著影響(圖2D)。
3 討論
糖酵解與糖異生在生理功能上分別是糖的分解與糖的合成, 前者能在魚體攝入過多糖分時降解血糖, 避免血糖高造成的生理負擔; 后者則是在魚體血糖降低時合成葡萄糖, 維持血糖內穩態, 魚體需要二者的共同調節才能正常地分解和轉化糖分[31]。與哺乳動物糖代謝機制類似, 調節魚類糖酵解的關鍵酶為己糖激酶(HK)、葡萄糖激酶(GK)、磷酸果糖激酶(PFK)、丙酮酸激酶(PK), 調節糖異生的關鍵酶為磷酸烯醇式丙酮酸羧激酶(PEPCK)、1, 6-二磷酸果糖酶(FBPase)、葡萄糖-6-磷酸酶(G6Pase)。飼料添加不同糖源及水平對大菱鲆 G6Pase活性的影響將在后續實驗中展開研究。
3.1 不同糖源及糖水平對大菱鲆糖酵解酶活性的影響
在歐洲海鱸、虹鱒、鯉魚(Cyprinus carpio)、草魚和金頭鯛等魚類中已證實了 GK的存在, 且在肝臟中活性最高, 并隨飼料糖水平的增加而顯著升高[8,19,23,32,33]。本研究也發現, 飼料中葡萄糖、蔗糖、糊精添加量與大菱鲆肝臟 GK活性呈正相關, 支持上述觀點。但當飼料葡萄糖或糊精添加量從15%增加至28%時, GK值不再增加或有降低趨勢, 推測可能是大菱鲆肝臟 GK對糖的適應能力有限, 添加15%的葡萄糖或糊精已促進GK活性達到飽和狀態。歐洲海鱸的 GK活性隨飼料淀粉含量的變化亦有相似的趨勢[34]。不同糖源對魚類GK活性的誘導能力也存在差異, 與分子量較大的糖類相比, 分子量較小的糖類更能促進魚體內GK活性的增高。Enes,et al.[7]研究發現, 在飼料中添加 20%的葡萄糖、麥芽糖、糊精、淀粉后金頭鯛體內GK活性在葡萄糖組和麥芽糖組最高。飼料添加葡萄糖對歐洲海鱸 GK活性的誘導作用較淀粉明顯[8]。在本研究中, 糖添加水平為28%時, 葡萄糖組、蔗糖組對大菱鲆體內GK的誘導作用顯著高于糊精組, 支持上述觀點。
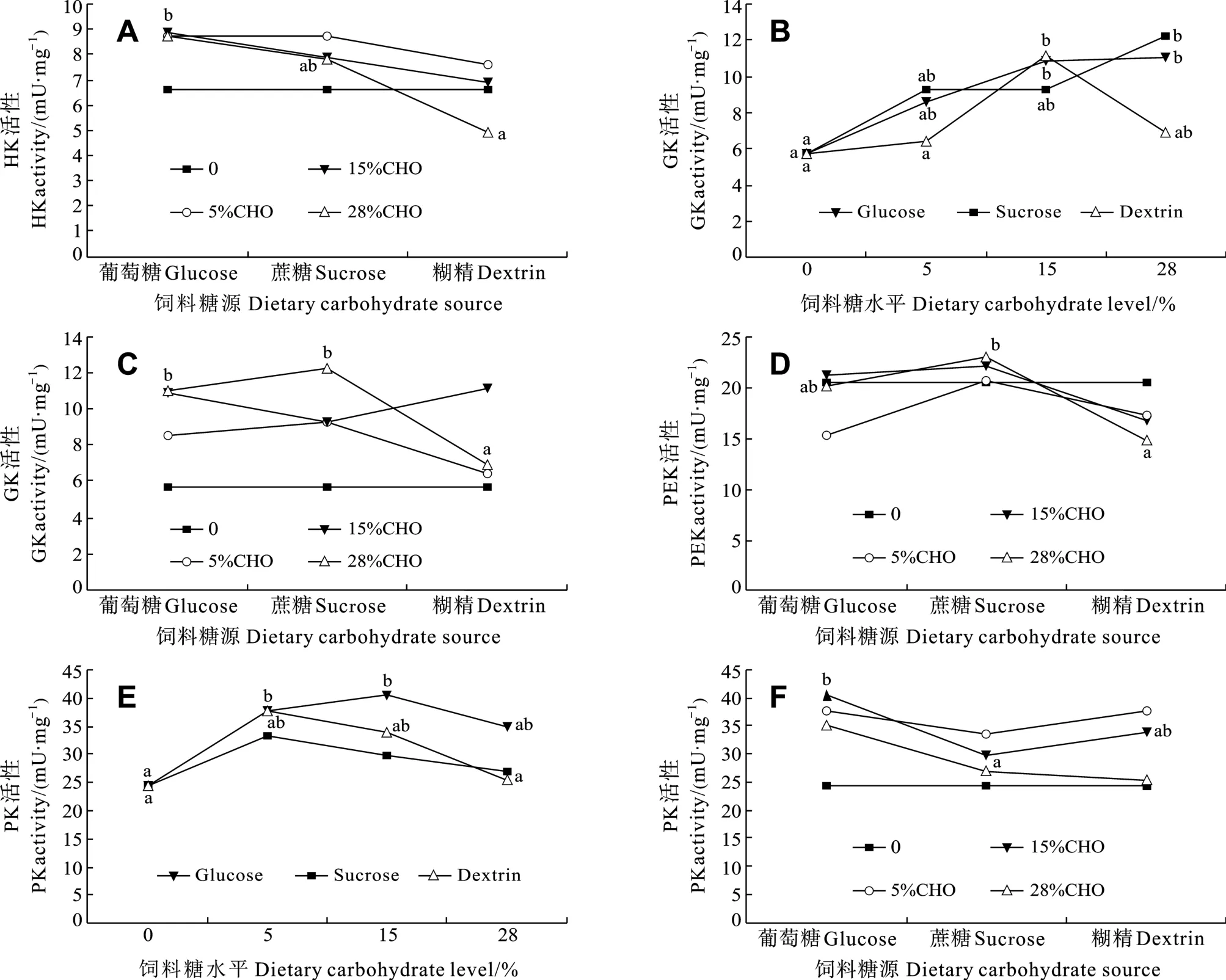
圖1 飼料中不同糖源及糖水平對大菱鲆肝臟中己糖激酶(HK)、葡萄糖激酶(GK)、磷酸果糖激酶(PFK)和丙酮酸激酶(PK)活性的影響Fig.1 Effects of dietary carbohydrate sources and levels on activities of hexokinase (HK), glucokinase (GK), 6-phosphofructokinase-1(PFK) and pyruvate kinase (PK) in the liver of turbot
有關 PK的研究表明, 高糖飼料能誘導魚類肝臟 PK的活性增加[1,2,23,35]。飼料中糖源為葡萄糖或糊精時, 大菱鲆體內 PK活性隨葡萄糖或糊精水平增加而升高, 當葡萄糖添加量為 15%或糊精水平為5%時, PK活性呈最大值并顯著高于對照組。但隨葡萄糖或糊精水平的進一步增加, PK出現了類似GK在高糖情況下(添加量為 28%)活性降低的變化, 這也與Meton,et al.[1]研究不同水平淀粉對金頭鯛PK活性的影響時變化趨勢一致。同時, 大菱鲆肝臟PK活性也受飼料中糖源的顯著影響, 當糖添加量為15%時, 葡萄糖組PK活性顯著高于蔗糖組, 但與糊精組無顯著差異。Enes,et al.發現, 金頭鯛PK活性不受飼料添加20%葡萄糖、麥芽糖或糊精的影響[6],但歐洲海鱸攝食含20%糖的飼料后PK活性在葡萄糖組顯著高于淀粉組[8]。產生這種差異的原因是否是由于物種的不同或實驗條件差異, 導致對 PK活性的影響不同, 有待于進一步證實。
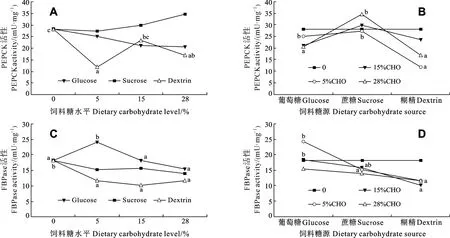
圖2 飼料中不同糖源及水平對大菱鲆中肝臟中磷酸烯醇式丙酮酸羧激酶(PEPCK)和1,6-二磷酸果糖酶(FBPase)活性的影響Fig.2 Effects of dietary carbohydrate sources and levels on activities of phosphoenolpyruvate carboxykinase (PEPCK) and fructose-1,6-bisphosphatase (FBPase) in the liver of turbot
在大菱鲆肝臟糖酵解途徑中, HK和PFK不受飼料中糖水平的顯著影響, 即HK和PFK活性不受飼料糖含量的誘導, 這分別與在歐洲海鱸[8,23,34]、金頭鯛[22]、虹鱒[36]和南方鲇(Silurus meridionalisChen)[31]等中的研究結果一致。相比之下, GK和PK受糖源和糖水平的顯著影響, 顯示GK和PK是決定大菱鲆肝臟糖酵解能力的兩個重要因素。在飼料糖水平低于 15%時, 大菱鲆的糖酵解能力隨之出現適應性的增強, 這也合理解釋了大菱鲆肝糖原含量隨飼料糖水平增加而增加這一結果(未發表數據)。但飼料糖含量繼續增加至 28%時, 大菱鲆肝臟的糖酵解能力沒有繼續增強, 反而有下降的趨勢。在研究糖源和糖水平對大菱鲆生長的影響時也發現, 飼料含較低水平的糖時不影響大菱鲆的生長, 但高水平的糖(添加量為 28%)導致大菱鲆特定生長率顯著降低(未發表數據)。這表明了大菱鲆對飼料糖水平的適應能力有限, 飼料添加 15%的糖可能已達到了大菱鲆機體代謝利用糖的最大限度。比較3種糖源對大菱鲆糖酵解的促進作用, 葡萄糖的促進作用最強,糊精次之。
3.2 不同糖源和糖水平對大菱鲆糖異生酶活性的影響
本研究發現, 在大菱鲆的糖異生途徑中,PEPCK和FBPase兩種酶都受飼料糖源及水平的顯著影響, 且交互作用顯著。當飼料添加葡萄糖或蔗糖時, PEPCK活性不受影響。在糊精組中, 隨飼料糊精水平的增加, PEPCK活性雖有波動變化, 但整體呈下降的趨勢。戈賢平等[37]鲌對翹嘴紅 (Eryghroculter ilishaeformis)攝食添加不同水平淀粉的飼料后 12h取樣分析表明, 中糖組(14.45%)、高糖組(23.98%)比無糖組顯著降低了 PEPCK活性, 與本研究結果一致。但Kirchner,et al.[38]研究認為虹鱒攝食含20%糊精的飼料后 PEPCK的活性和基因表達量與無糖對照組之間均無顯著差異。同時, 本研究還發現在葡萄糖組中, FBPase活性僅隨飼料中添加低水平葡萄糖(5%)的誘導而升高, 但葡萄糖水平進一步增加時, FBPase活性與不含糖的對照組之間無顯著差異。在糊精組中, FBPase活性則受到飼料添加糊精的顯著抑制。Panserat,et al.[39]研究發現金頭鯛攝食含20%糖的飼料后, FBPase基因表達水平顯著低于攝食不含糖的飼料, 即飼料含較高水平的糖時顯著抑制了FBPase的基因表達。但Kirchner,et al.[38]研究認為虹鱒的 FBPase活性和基因表達量在攝食含 20%糊精和不含糖的飼料之間無顯著差異, 并認為虹鱒體內的葡萄糖穩態失衡可能與糖異生酶活性或基因表達不受飼料糖含量的調節有關。在本研究中大菱鲆肝臟糖異生關鍵酶PEPCK和FBPase活性受飼料添加糊精的抑制, 這一結果說明了其體內可能存在較為完善的糖異生調節機制。但同時, 本研究所采用的實驗飼料為等氮等能設計, 因此在飼料糖水平增加時脂肪含量隨之降低。Panserat,et al.[40]報道含高脂肪(25%)的飼料顯著促進了虹鱒肝臟G6Pase基因表達和酶活性, 但對PEPCK和FBPase基因表達并無影響。這表明飼料中脂肪含量的變化在一定程度上也影響了魚體的糖異生能力, 這是否也是本研究中PEPCK和FBPase活性變化的一個影響因素, 有待于進一步研究。
綜上所述, 飼料添加 15%的葡萄糖或糊精時,可以有效促進大菱鲆肝臟糖酵解酶的活性, 同時低水平的葡萄糖促進糖異生酶活性; 糊精在促進糖酵解的同時對糖異生作用存在一定程度的抑制。在本實驗條件下, 較添加其他糖源或糖水平, 飼料添加15%的糊精更有利于維系大菱鲆機體內葡萄糖的動態平衡。
[1] Meton I, Mediavilla D, Caseras A,et al.Effect of diet composition and ration size on key enzyme activities of glycolysis-gluconeogenesis, the pentose phosphate pathway and amino acid metabolism in liver of gilthead sea bream (Sparus aurata) [J].British Journal of Nutrition, 1999, 82:223—232
[2] Fernández F, Anna G M, Córdoba M,et al.Effects of diets with distinct protein-to-carbohydrate ratios on nutrient digestibility, growth performance, body composition and liver intermediary enzyme activities in gilthead sea bream(Sparus aurataL.)fingerlings [J].Journal of Experimental Marine Biology and Ecology, 2007, 343: 1—10
[3] Wilson R P.Utilization of dietary carbohydrate by fish [J].Aquaculture, 1994, 124: 67—80
[4] Dabrowski K, Guderley H.Intermediary metabolism [A].In:Halver J E, Hardy R W (Eds.), Fish Nutrition, 3rd edn [C].Academic Press, London.2002, 309—365
[5] Aikins K F, Hung S S O, Hughes S G,et al.Effects of feeding a high level of D-glucose on liver function in juvenile white sturgeon(Acipenser transmontanus) [J].Fish Physiology and Biochemistry, 1993, 12(4): 317—325
[6] Ma A J, Chen S Q, Lei J L,et al.Influences of the main energy matter in feed on the growth of young turbot,Scophthalmus maximus[J].Oceanologia Etlimnologia Sinica,2001, 32(5): 527—533 [馬愛軍, 陳四清, 雷霽霖, 等.飼料中主要能量物質對大菱鲆幼魚生長的影響.海洋與湖沼, 2001, 32(5): 527—533]
[7] Enes P, Peres H, Couto A,et al.Growth performance and metabolic utilization of diets including starch, dextrin, maltose or glucose as carbohydrate source by gilthead sea bream(Sparus aurata) juveniles [J].Fish Physiology and Biochemistry, 2010, 36: 903—910
[8] Enes P, Panserat S, Kaushik S,et al.Rapid metabolic adaptation in European sea bass (Dicentrarchus labrax) juveniles fed different carbohydrate sources after heat shock stress [J].Comparative Biochemistry and Physiology,Part A, 2006,145: 73—81
[9] Lin J H, Shiau S Y.Hepatic enzyme adaptation to different dietary carbohydrates in juvenile tilapiaOreochromis niloticus × O.aureus[J].Fish Physiology and Biochemistry, 1995,14(2): 165—170
[10] Shiau S Y, Lin Y H.Utilization of glucose and starch by the grouperEpinephelus malabaricusat 23°C [J].Fisheries Science, 2002, 68: 991—995
[11] Shikata T, Iwanaga S, Shimeno S.Effects of dietary glucose,fructose, and galactose on hepatopancreatic enzyme activities and body composition in carp [J].Fisheries Science, 1994,60(5): 613—617
[12] Shiau S Y, Peng C Y.Protein-sparing effect by carbohydrates in diets for tilapia,Oreochromis niloticus×O.aureus[J].Aquaculture, 1993, 117: 327—334
[13] Lee S M, Kim K D, Lall S P.Utilization of glucose, maltose,dextrin and cellulose by juvenile flounder (Paralichthys olivaceus) [J].Aquaculture, 2003, 221: 427—438
[14] Tan Q, Xie S Q, Zhu X,et al.Effect of dietary carbohydrate sources on growth performance and utilization for gibel carp(Carassius auratus gibelio) and Chinese longsnout catfish(Leiocassis longirostrisGünther) [J].Aquaculture Nutrition,2006, 12: 61—70
[15] Panserat S, Capilla E, Gutierrez J,et al.Glucokinase is highly induced and glucose-6-phosphatase poorly repressed in liver of rainbow trout (Oncorhynchus mykiss) by a single meal with glucose [J].Comparative Biochemistry and Physiology,Part B, 2001, 128: 275—283
[16] Bergot F.Carbohydrate in rainbow trout diets: effects of the level and source of carbohydrate and the number of meals on growth and body composition [J].Aquaculture, 1979, 18:157—167
[17] Buhler D R, Halver J E.Nutrition of salmonid fishes [J].The Journal of Nutrition, 1961, 74: 307—318
[18] Hung S S O, Storebakken T.Carbohydrate Utilization by Rainbow Trout Is Affected by Feeding Strategy [J].Journal of Nutrition, 1994, 124: 223—230
[19] Enes P, Panserat S, Kaushik S,et al.Hepatic glucokinase and glucose-6-phosphatase responses to dietary glucose and starch in gilthead sea bream (Sparus aurata) juveniles reared at two temperatures [J].Comparative Biochemistry and Physiology,Part A, 2008, 149: 80—86
[20] Hung S S O, Fynn A K, Lutes P,et al.Ability of juvenile white sturgeon (Acipenser transmontanus) to different carbohydrate sources [J].Journal of Nutrition, 1989, 119:727— 733
[21] Tian L X, Liu Y J, Hung S S O.Utilization of glucose and corn starch by juvenile grass carp [J].North American Journal of Aquaculture, 2004, 66: 141—145
[22] Couto A, Enes P, Peres H,et al.Effect of water temperature and dietary starch on growth and metabolic utilization of diets in gilthead sea bream (Sparus aurata) juveniles [J].Comparative Biochemistry and Physiology,Part A,2008,151: 45—50
[23] Enes P, Panserat S, Kaushik S.Effect of normal and waxy maize starch on growth, food utilization and hepatic glucose metabolism in European sea bass (Dicentrarchus labrax) juveniles [J].Comparative Biochemistry and Physiology,Part A, 2006, 143: 89 —96
[24] Kumar V, Sahu N P, Pal A K,et al.Modulation of key enzymes of glycolysis, gluconeogenesis, amino acid catabolism,and TCA cycle of the tropical freshwater fi shLabeo rohitafed gelatinized and non-gelatinized starch diet [J].Fish Physiology and Biochemistry, 2010, 36: 491—499
[25] Hanson R W, Reshef L.Regulation of phosphoenolpyruvate carboxykinase (GTP) gene expression [J].Annual Review of Biochemistry, 1997, 66: 581—611
[26] Li X N.Effects of dietary carbohydrate levels on growth performance, physiological status and body composition of turbot (Scophthalmus maximusL) and Japanese flounder(Paralichthys olivaceus) [D].Thesis for Master of Science.Ocean University of China, Qingdao.2011 [李曉寧.飼料糖水平對大菱鲆和牙鲆生長、生理狀態參數及體組成的影響.碩士學位論文, 中國海洋大學.青島.2011]
[27] He J H.Analyse and Check of Feedstuff [M].Beijing: China Agricultural Press.2005, 47 [賀建華.飼料分析與檢測.北京: 中國農業出版社.2005, 47]
[28] Polakof S, Míguez J M, Soengas J L.Dietary carbohydrates induce changes in glucosensing capacity and food intake of rainbow trout [J].American Journal of Physiology Regulatory Integrative and Comparative Physiology, 2008, 295:478—489
[29] Bradford M.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle dye binding [J].Analytical Biochemistry, 1976, 72: 248—254
[30] Hamid N K A, Mahayat M, Hashim R.Utilization of different carbohydrate sources and starch forms by bagrid catfish(Mystus nemurus) (Cuv&Val) [J].Aquaculture Nutrition,2011, 17: 10—18
[31] Lin X Z, Luo Y P, Xie X J.Effects of dietary carbohydrate level on glycolytic enzymes and serum glucose concentrations in the juvenile southern catfish,Silurus meridionalisChen after feeding [J].Acta Hydrobiologica Sinica, 2006,30(3): 304—310 [林小植, 羅毅平, 謝小軍.飼料碳水化合物水平對南方鲇幼魚餐后糖酵解酶活性及血糖濃度的影響.水生生物學報, 2006, 30(3): 304—310]
[32] Panserat S, Medale F, Blin C,et al.Hepatic glucokinase is induced by dietary carbohydrates in rainbow trout, gilthead sea bream, and common carp [J].American Journal of Physiology Regulatory Integrative and Comparative Physiology, 2000, 278: 1164—1170
[33] Zhang S L, Ai Q H, Xu W,et al.Effects of dietary carbohydrate to lipid ratio on growth, feed utilization, plasma glucose and glycolytic enzyme activities of juvenile darkbarbel catfish,Pelteobagrus vachelli[J].Acta Hydrobiologica Sinica, 2012, 36(3): 466-473 [張世亮, 艾慶輝, 徐瑋, 等.飼料中糖/脂肪比例對瓦氏黃顙魚生長、飼料利用、血糖水平和肝臟糖酵解酶活力的影響.水生生物學報, 2012,36(3): 466-473]
[34] Moreira I S, Peres H, Couto A,et al.Temperature and dietary carbohydrate level effects on performance and metabolic utilisation of diets in European sea bass (Dicentrarchus labrax)juveniles [J].Aquaculture, 2008, 274: 153—160
[35] Borrebaek B, Christophersen B.Hepatic glucose phosphorylating activities in perch (Perca fluviatilis) after different dietary treatments [J].Comparative Biochemistry and Physiology,Part B, 2000, 125: 387—393
[36] Panserat S, Médale F, Brèque J E,et al.Lack of significant long-term effect of dietary carbohydrates on hepatic glucose-6-phosphatase expression in rainbow trout (Oncorhynchus mykiss) [J].The Journal of Nutritional Biochemistry, 2000,11: 22—29
[37] Ge X P.Effects of different carbohydrate and lipid levels in diets on carbohydrate metabolic enzymes in Topmouth Culter (Eryghroculter ilishaeformisbleeker) [D].Thesis for Doctor of Science.Nanjing Agricultural University, Nanjing.2006 [戈賢平.不 同糖、脂含量日糧對翹嘴紅鲌 相關糖代謝酶的調節研究.博士學位論文, 南京農業大學, 南京.2006]
[38] Kirchner S, Panserat S, Lim P L,et al.The role of hepatic,renal and intestinal gluconeogenic enzymes in glucose homeostasis of juvenile rainbow trout [J].Journal of Comparative Physiology B, 2008, 178: 429—438
[39] Panserat S, Plagnes-Juan E, Kaushik S.Gluconeogenic enzyme gene expression is decreased by dietary carbohydrates in common carp (Cyprinus carpio) and gilthead seabream(Sparus aurata) [J].Biochimica et Biophysica Acta, 2002,1579: 35—42
[40] Panserat S, Perrin A, Kaushik S.High dietary lipids induce liver glucose-6-phosphatase expression in rainbow trout(Oncorhynchus mykiss) [J].Biochemical and Molecular Action of Nutrients, 2002, 132(2): 137—141
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
