溫度對中型指環蟲產卵和孵化的影響
2013-07-24 11:29:44張效平李文祥艾桃山吳山功鄒王桂堂
水生生物學報 2013年3期
張效平 李文祥 艾桃山 吳山功鄒 紅 王桂堂
(1.中國科學院水生生物研究所, 武漢 430072; 2.中國科學院大學, 北京 100049;3.武漢中博水產生物技術有限公司, 武漢 430207)
隨著魚類養殖密度的提高和養殖環境的惡化,單殖吸蟲病的發生日益頻繁和嚴重, 給水產養殖造成較大的經濟損失[1,2]。單殖吸蟲一般寄生于魚類的鰓和體表, 以其后吸器的幾丁質結構插入魚類組織,破壞鰓及皮膚[3], 可刺激鰓細胞分泌過多黏液, 引起鰓絲黏連和壞死, 影響魚類呼吸[4],也可破壞魚類體表組織, 引起細菌、真菌和病毒的繼發性感染, 導致魚苗甚至成魚的大量死亡[5]。
中型指環蟲(Dactylogyrus intermedius)是寄生于鯽(Carassius auratus)、鯉(Cyprinus carpio)鰓部一種較常見的單殖吸蟲, 分布廣泛, 在國內外均有發現和報道[6—8]。近年來, 國內外的一些學者研究了中草藥提取物對中型指環蟲的殺滅效果[9—11], 但是還沒有中型指環蟲生活史方面的研究報道。由于單殖吸蟲生活史的每一階段都極大的受制于溫度, 其產卵、孵化和纖毛幼蟲的生長發育速度與溫度密切相關[12,13]。本實驗將以金魚(Carassius auratus)鰓部的中型指環蟲為研究對象, 研究在離體條件下溫度對中型指環蟲產卵和孵化的影響。這將為研究有效的生態控制技術和藥物防治方法提供理論依據。
1 材料與方法
1.1 實驗魚
實驗魚購自武漢市武昌區八一路花鳥市場, 將金魚養在盛有曝氣自來水的水族箱中。
1.2 中型指環蟲的收集
用MS-222將金魚麻醉后, 剪下鰓片, 放在載玻片上, 在解剖鏡下進行觀察。當發現指環蟲時, 迅速將其剝離下來, 立即放到盛有少量曝氣自來水的 24孔板中, 并對指環蟲進行清洗。在倒置顯微鏡下觀察指環蟲的形態特征, 如果指環蟲蟲體的體長在0.24—0.38 mm, 中央大鉤較短, 鉤尖平直, 其基部的背葉和腹葉分叉明顯, 背葉較短, 聯接棒單一,交配管呈斜管狀, 基部較寬, 且帶有基座[14], 可鑒定為中型指環蟲, 并將收集到的蟲體用來產卵和孵化實驗。
1.3 中型指環蟲的產卵
將收集的中型指環蟲放到盛有曝氣自來水的24孔板中, 一孔一蟲, 分別置于 10、22、30、35℃的生化培養箱和4℃的冰箱內, 前5h內每隔1h取出觀察一次, 之后每2h觀察一次, 記錄每條指環蟲產卵的數量, 直至蟲體死亡或連續 12h沒有產卵時實驗終止。
1.4 蟲卵的孵化
收集到中型指環蟲產下的蟲卵, 放到盛有曝氣自來水的24孔板中, 分別置于10、22、30、35℃的生化培養箱和 4℃的冰箱內, 在眼點出現之前每隔12h取出觀察一次, 當出現眼點后每 2h觀察一次,記錄蟲卵的變化情況, 直到所有蟲卵都孵化或變黑死亡, 實驗終止。
1.5 數據處理
在各溫度下產卵數量的顯著性檢驗采用單因素的方差分析(ANOVA), 孵化率采用卡方檢驗, 統計分析用軟件SPSS 13.0。
2 結果
2.1 中型指環蟲的產卵
產卵過程 在實驗中觀察到, 蟲卵形成很快,當將中型指環蟲移入盛有曝氣自來水的 24孔板后,第一枚卵在1min之內就可排出。第一枚卵排出后大約 2min, 另一枚卵在后面接著開始形成, 在顯微鏡下可以清楚地看到卵粒的形成和增大的過程。在蟲卵形成過程中, 蟲體做波浪狀的收縮和舒張運動,蟲卵越來越大, 大約6、7min后蟲卵完全成熟, 排出體外。但是后面蟲卵的形成時間和產卵過程越來越長。
卵的形態 在所觀察的正常卵中(能夠孵化出纖毛幼蟲的卵), 除了小部分為圓形, 大部分為梨形, 長約 30 μm; 蟲卵顏色較深, 后端有一個卵柄;蟲體最后產出的幾枚蟲卵, 形狀一般不規則, 顏色較淺, 近透明, 大小不一, 一般小于正常卵, 而且卵殼較薄, 容易破裂, 這些非正常卵不能孵化出幼蟲(圖 1)。

圖1 中型指環蟲在離體條件下所產的正常卵(左)與非正常卵(右)Fig.1 Normal (left) and abnormal (right) egg of Dactylogurus intermedius
溫度對中型指環蟲產卵的影響 在 4℃條件下, 大部分中型指環蟲直到死亡也不產卵。只有少數已懷有發育完全的卵粒的中型指環蟲會產一枚卵。在此溫度條件下, 中型指環蟲成蟲最長存活時間為9d。
在 10—35℃溫度范圍內, 平均產卵量隨著溫度的升高而增加, 其平均產卵量分別為 3.30、4.10、4.13和 4.24枚/蟲(圖 2), 統計結果顯示: 在 35℃條件下的產卵量明顯高于10℃(P< 0.05), 其他溫度條件下的平均產卵量沒有顯著性差異; 中型指環蟲的產卵速率隨著溫度的升高而加快, 產卵維持的時間分別為4d、23h、15h和11h。
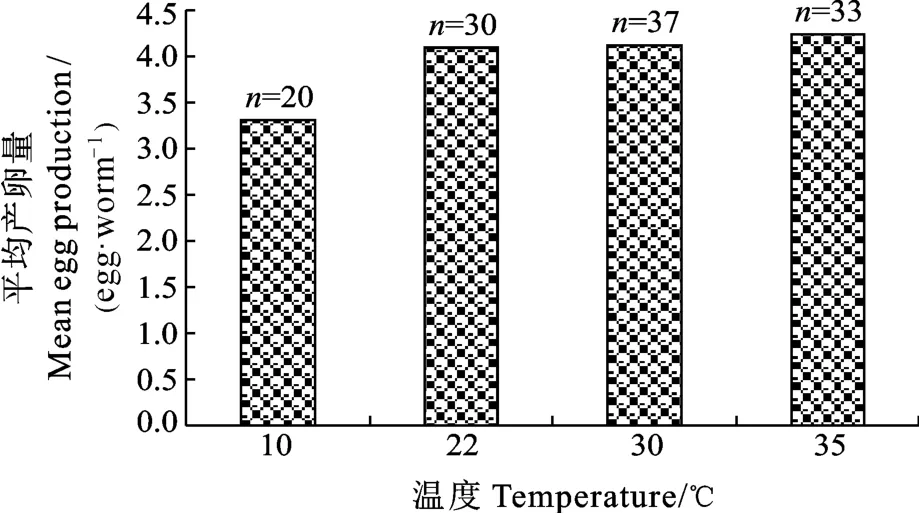
圖2 不同溫度條件下中型指環蟲的平均產卵量(n代表中型指環蟲的數量)Fig.2 Mean egg production of Dactylogyrus intermedius at 10℃,22℃, 30℃ and 35℃ (n represents the number of D.intermedius examined)
2.2 中型指環蟲蟲卵的孵化

圖3 中型指環蟲蟲卵在孵化過程中的變化Fig.3 Stages of Dactylogyrus intermedius in egg hatching
蟲卵的孵化過程 剛排出的蟲卵為灰黑色, 當蟲卵在 22℃孵育 24h后, 開始出現黑色眼點(圖 3a)。在孵育2d后, 纖毛幼蟲的輪廓明顯, 還可看到纖毛幼蟲后吸盤上的小鉤(圖 3b)。有時可見蟲體在卵殼內蠕動, 以頭部頂卵殼上的卵蓋。大約在孵化 96h后, 大部分幼蟲頂破卵蓋而成功鉆出, 留下一個空殼(圖 3c)。有些幼蟲在蠕動的過程中, 頭部偏離了卵蓋的位置, 無法從卵蓋處鉆出, 而是從中間或后端頂開一個洞而鉆出。還有一部分幼蟲在鉆出的過程中, 只有頭部鉆出卵殼而身體后端卻被卡在卵殼內, 造成死亡。沒有鉆出卵殼的纖毛幼蟲則死亡, 慢慢變黑。
纖毛幼蟲的形態 剛孵化出的纖毛幼蟲呈圓筒狀, 兩端稍尖, 兩對眼點大而清晰; 后吸盤具有若干對小錨鉤, 且小錨鉤在不停地做伸縮運動, 大錨鉤收縮在體內; 纖毛幼蟲具有三環纖毛, 分別分布在前部、中部和尾部(圖4), 纖毛不停地在水中擺動, 纖毛幼蟲有時靜止在某一水層, 有時又非常活躍, 時而急速前行, 時而做“8”字形運動, 并不時地以頭器與孔壁接觸。
溫度對中型指環蟲孵化的影響 在 4℃條件下, 中型指環蟲蟲卵不能孵化, 但是將蟲卵放在 4℃條件下保存一周后, 再取出放在室溫(約 27℃)下, 蟲卵能成功孵化,且孵化率達到60%。
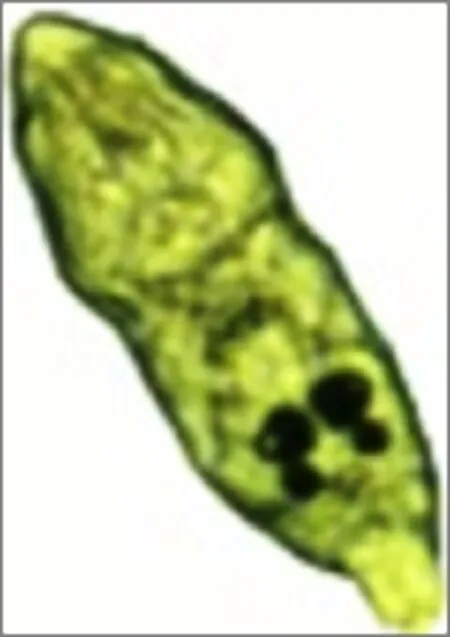
圖4 剛孵化的纖毛幼蟲的形態Fig.4 Morphology of newly hatched oncomiracidia of Dactylogyrus intermedius
在 22℃條件下, 中型指環蟲的孵化率最高,為 72.7%, 其次是 10℃,而在30℃和35℃條件下的孵化率最低, 約 50%(圖5)。卡方檢驗結果顯示 4種溫度條件下的孵化率無顯著差異(P>0.05)。隨著溫度的升高,蟲卵的孵化速度明顯增加, 即孵化的時間逐漸減少, 在 10℃下, 中型指環蟲卵最快需要 24d才能孵化, 在 22℃下, 蟲卵在第2d即開始出現眼點, 3d后開始孵化, 而當溫度升至30℃和 35℃時, 蟲卵的平均孵化時間也分別降至42h和26h。纖毛幼蟲最長存活時間隨著溫度的升高而減少, 在 4個溫度條件下的最長存活時間分別為4d、3d、56h 和 34h。
3 討論
3.1 離體條件下卵的形態和產卵速率以及溫度對產卵速率的影響
在離體條件下, 中型指環蟲最后產出的幾枚卵在大小、形狀和顏色等特征上與正常卵差距明顯,而且不能孵化出幼蟲。卵的形成需要消耗蟲體的營養物質, 而在離體條件下, 隨著蟲卵不斷的形成和排出, 成蟲體內的營養物質逐漸被消耗, 由于得不到充足的營養補充, 最后形成的幾枚卵由于營養不足, 而出現異常。另外, 該指環蟲前幾枚卵的排出速率較快, 只需要幾分鐘, 而后面的產卵速率越來越慢,這應該與指環蟲在離體條件下的應激產卵有關[15]。在魚體上, 指環蟲的產卵一般分為兩種情況, 一種是產卵季節性很強, 只在某特定時期產卵, 由于事先積累大量卵, 因此產卵的速率很快; 另外一種情況是產卵季節性弱, 卵一經形成即產出, 產卵速率較慢、較穩定, 易受溫度影響[16]。在本實驗中, 成熟的中型指環蟲蟲體內只存在一枚蟲卵, 并沒有積累多枚卵, 產卵類型類似于后者。
指環蟲的產卵速率與溫度密切相關, 在10、22、30、35℃溫度下, 中型指環蟲的產卵維持的時間分別為4d、23h、15h和11h, 即產卵速率隨著溫度的升高而加快。當溫度從 5℃升高到 18℃時, 單殖吸蟲Discocotyle sagittata的產卵速率從 1.5枚/(蟲?d)上升到 12枚/(蟲?d)[17]; 當溫度從 10℃升到20℃時,單殖吸蟲Neoheterobothrium hirame的產卵速率也由 8.5 枚/(蟲?h)上升到 32.55 枚/(蟲?h)[18]。可見, 在一定溫度范圍內, 隨著溫度升高, 單殖吸蟲的產卵速率也隨著加快。
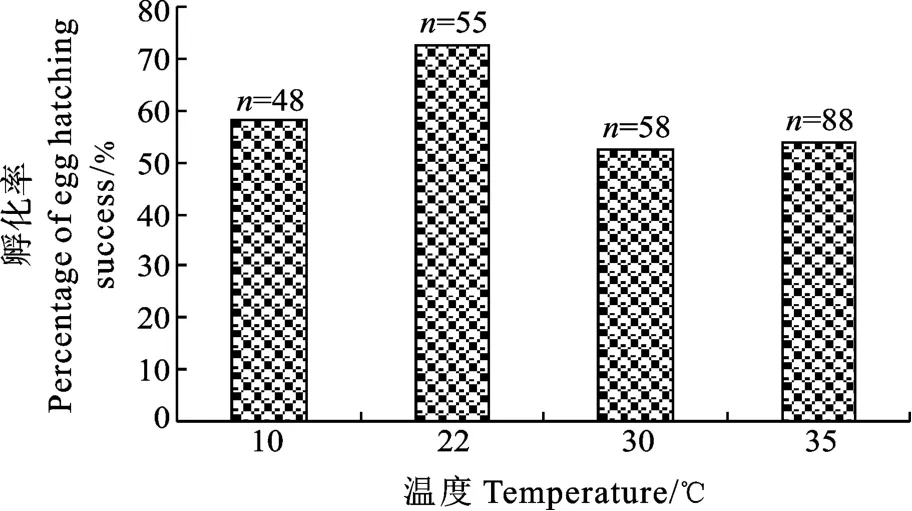
圖5 不同溫度條件下中型指環蟲蟲卵的孵化率(n代表蟲卵的數量)Fig.5 Hatching rate of D.intermedius at 10℃, 22℃, 30℃ and 35℃(n represents the number of eggs)
3.2 溫度對產卵量的影響
單殖吸蟲的產卵量也受到溫度的影響。在 4℃條件下, 中型指環蟲基本上不產卵, 當溫度從 10℃升到 35℃, 中型指環蟲的平均產卵量從 3.30枚/蟲上升至 4.24枚/蟲, 且 35℃條件下的平均產卵量顯著高于10℃的。秉偽指環蟲(Pseudodactylogyrus bini)在 24h內的平均產卵量在 30℃最高, 在 5℃條件下秉偽指環蟲基本上不產卵[19]。在 17.5℃條件下, 本尼登蟲(Benedenia seriolae)和軛聯蟲(Zeuxapta seriolae)的平均產量均達到最高, 分別為61.4枚/蟲和309.3枚/蟲[12]。在14℃條件下, 小鞘指環蟲(Dactylogyrus vaginalatus)的產卵量最大, 低于或高于此溫度, 產卵量均下降[20]。不同的單殖吸蟲種類, 其產卵量達到最大值的溫度各不相同, 與小鞘指環蟲、秉偽指環蟲、本尼登蟲和軛聯蟲相比, 中型指環蟲產卵量達到最大值的溫度更高。
3.3 溫度對孵化速率和孵化率的影響
同樣, 溫度對蟲卵的孵化速率和孵化率也都有較大的影響。在4℃條件下, 中型指環蟲蟲卵不能孵化, 在 10—35℃的溫度范圍內, 蟲卵的孵化速率不斷增加, 孵化率卻在 22℃最高。在 15℃、20℃和 23℃三個溫度條件下, 河鱸錨首吸蟲(Ancyrocephalus mogurndae)的孵化速率隨著溫度的升高而加快, 而孵化率卻在15℃時最高[21]。在13—21℃范圍內, 本尼登蟲蟲卵的孵化率無明顯差異, 均在 0.73—0.78;而軛聯蟲的蟲卵在13℃下的孵化率要明顯低于17.5℃和 21℃[12]。單殖吸蟲Bivagina tai[22]和Neoheterobothrium hirame[23]的蟲卵在高溫條件下(30℃)孵化率均下降。秉偽指環蟲的蟲卵在34℃條件下可以發育, 但是卻沒有發現成功孵化, 且在 37℃條件下其蟲卵的發育和孵化完全受到抑制[19]。可見, 單殖吸蟲的孵化速率達到最大值時的溫度往往高于孵化率達到最高值時的溫度, 也就是說, 孵化速率達到最大值時的溫度并不一定是最適的孵化溫度。在35℃的高溫條件下, 中型指環蟲的蟲卵還能成功孵化,孵化速率也是最快的, 但是孵化率卻降低, 說明此溫度已經對蟲卵孵化產生一定的抑制作用。另外,本研究還發現在4℃低溫保存7d后的蟲卵仍能再成功孵化, 且孵化率沒有明顯降低。但是低溫(5℃)處理Diplectanum aequans的蟲卵后, 在室溫(18℃)條件下的孵化率卻顯著降低[24]。可見, 溫度對不同種類單殖吸蟲的產卵、孵化等產生不同程度的影響。
通過本實驗的結果, 可以初步得出, 當水溫為22℃時, 中型指環蟲的產卵數量和纖毛幼蟲的存活時間都比較高, 且孵化率最高, 表明在該溫度條件下, 中型指環蟲種群增長迅速, 容易引起指環蟲病的暴發和流行。
[1] Buchmann K, Slotved H C, Dana D.Gill parasites fromCyprinus carpioin Indonesia [J].Aquaculture, 1995, 129:437—439
[2] Dove A, Ernst I.Concurrent invaders-four exotic species of Monogenea now established on exotic freshwater fishes in Australia [J].International Journal for Parasitology, 1998,28: 1755—1764
[3] Jalali B, Barzegar M.Dactylogyrids (Dactylogyridae:Monogenea) on common carp (Cyprinus carpioL.) in freshwaters of Iran and description of the pathogenicity ofD.sahuensis[J].Journal of Agriculture,Science and Technology, 2005, 7: 9—16
[4] Rahanandeh M, Jalali B, Sharifpour I,et al.Survey on Dactylogyrusis in Caspian Frisian roach (Rutilus fissi ktum)caused byDactylogyrus fissi[J].Global Veterinaria, 2010,4(5): 515—518
[5] Bandilla M, Valtonen E T, Suomalainen L R,et al.A link between ectoparasite infection and susceptibility to bacterial disease in rainbow trout [J].International Journal for Parasitology, 2006, 36: 987—991
[6] Kritsky D C, Heckmann R.Species ofDactylogyrus (Monogenoidea: Dactylogyridae) andTrichodina mutabilis(Ciliata)infesting koi carp,Cyprinus carpio, during mass mortality at a commercial rearing facility in Utah, U.S.A [J].Comparative Parasitology, 2002, 69(2): 217—218
[7] Grupcheva G I, Nedeva I L.Parasite fauna of the crucian carp (Carassius auratus gibelioBloch.) in the Zrebchevo reservoir (Bulgaria) [J].Acta Zoologica Bulgarica, 2000,51(1): 115—122
[8] Wang K Y, Yao L, Xie J B,et al.Effect of the active monomer fromRadix angelicae pubescentison killingDactylogyrus intermediusand its active component identification[J].Acta Hydrobiologica Sinica, 2012, 36(1): 93—101 [汪開毓, 姚璐, 謝嘉賓, 等.獨活活性單體對中型指環蟲的殺滅作用及其成分鑒定.水生生物學報, 2012, 36(1): 93—101]
[9] Wang G X, Zhou Z, Cheng C,et al.Osthol and isopimpinellin fromFructus cnidiifor the control ofDactylogyrus intermediusinCarassius auratus[J].Veterinary Parasitology,2008, 158: 144—151
[10] Liu Y T, Wang F, Wang G X,et al.In vivoanthelmintic activity of crude extracts ofRadix angelicae pubescentis,Fructus bruceae,Caulis spatholobi,Semen aesculi, andSemen pharbitidisagainstDactylogyrus intermedius(Monogenea) in goldfish (Carassius auratus) [J].Parasitology Research, 2010, 106: 1233—1239
[11] Wang G X, Zhao Y K, Shen Y H,et al.Study on the activity 25 natural plant extracts killing the Dactylogyrus [J].Journal of Northwest University(Natural Science Edition), 2011,41(1): 73—76 [王高學, 趙云奎, 申燁華, 等.25種植物提取物殺滅魚類指環蟲活性研究.西北大學學報, 2011,41(1): 73—76]
[12] Tubbs L A, Poortenaar C W, Sewell M A,et al.Effects of temperature on fecundityin vitro, egg hatching and reproductive development ofBenedenia seriolaeandZeuxapta seriolae(Monogenea) parasitic on yellowtail kingfishSeriola lalandi[J].International Journal for Parasitology, 2005, 35:315—327
[13] Lackenby J A, Chambers C B, Ernst I,et al.Effect of water temperature on reproductive development ofBenedenia seriolae(Monogenea: Capsalidae) from seriola lalandi in Australia [J].Diseases of Aquatic Organisms,2007, 74: 235—242
[14] Chen C L.An Illustrated Guide to the Fish Disease and Causative Pathogenic Fauna and Flora in the Hupei Province[M].Beijing: Science Press.1973, 118 [陳啟鎏.湖北省魚病病原區系圖志.北京: 科學出版社.1973, 118]
[15] Liu X D, Chen Q, Ke L,et al.Morphological observation on life cycle and embryonic development ofPseudodactylogyrus bini[J].Acta Parasitologica Et Medica Entomologica Sinica, 2008, 15(2): 75—77 [劉曉東, 陳強, 柯翎,等.秉偽指環蟲生活史形態觀察及發育條件研究.寄生蟲與醫學昆蟲學報, 2008, 15(2): 75—77]
[16] Bychowsky B E.Monogenetic Trematodes, Their Classification and Phylogenis [M].Moscow Leningrad: Academy of Sciences, U.S.S.R (in Russian).1957, 107
[17] Gannicott A M, Tinsley R C.Environmental effects on transmission ofDiscocotyle sagittata(Monogenea): egg production and development [J].Parasitology, 1998, 117:499—504
[18] Tsutsumi N, Mushiake K, Mori K,et al.Effects of water temperature on the egg laying of the monogeneanNeoheterobothrium hirame[J].Fish Pathology, 2002, 37: 41—43
[19] Buchmann K.Temperature-dependent reproduction and survival ofPseudodactylogyrus bini(Monogenea) on the European eel (Anguilla anguilla) [J].Parasitology Research,1988, 75: 162—164
[20] Yao W J.Studies on the characteristics of oviposition ofDactylogyrus vaginalatus[J].Journal of Huazhong Agriculture University, 2002, 21(1): 77—79 [姚衛建.小鞘指環蟲產卵特點的研究.華中農業大學學報, 2002, 21(1): 77—79]
[21] Xia X Q, Nie P, Yao W J.Effects of light, temperature and host mucus on the egg hatching ofAncyrocephalus mogurndae(Monogenea) [J].Acta Hydrobiologica Sinica, 1996,20(2): 195—196 [夏曉勤, 聶品, 姚衛建.光照、溫度及宿主黏液對河鱸錨首吸蟲蟲卵孵化的影響.水生生物學報,1996, 20(2): 195—196]
[22] Ogawa K.Development ofBivagina taiMonogenea: Microcotylidae.Nippon Suisan Gakkaishi-Bull [J].Fish Pathology,1988, 54(1): 61—64
[23] Yoshinaga T, Segawa I, Kamaishi T,et al.Effects of temperature, salinity and chlorine treatment on egg hatching of the monogeneanNeoheterobothrium hirameinfecting Japanese flounder [J].Fish Pathology, 2000, 35(2): 85—88
[24] Cecchini S, Saroglia M, Cognetti-Varriale A M,et al.Effect of low environmental temperature on embryonic development and egg hatching ofDiplectanum aequans(Monogenea,Diplectanidae) infecting European sea bass,Dicentrarchus labrax[J].Fish Pathology, 2001, 36(1): 33—34
