系統性紅斑狼瘡分子病因研究進展①
2013-11-27 11:15:42蘭州大學第二醫院腎內科蘭州730030
中國免疫學雜志 2013年6期
關鍵詞:研究
曹 平 趙 玉 王 靜 (蘭州大學第二醫院腎內科,蘭州730030)
系統性紅斑狼瘡(Systemic lupus erythematosus,SLE)是一種多系統損傷的自身免疫性疾病,以T、B細胞異常活化、補體激活、免疫復合物清除障礙、沉積、自身抗體的產生及免疫調節異常為特點[1,2]。世界流行病學調查為1.4% ~21.9%,發病率為(7.4~159.4)/10萬人,可造成多系統的損害,其中狼瘡腎炎(Lupus nephritis,LN)是最嚴重的并發癥之一,累積發病率亞洲人群最高,約為55%,非洲人群51%,其他各洲相對低,這些病人中23%約10年后發展為終末期腎病[3]。目前SLE病因、發病機理十分復雜,而且不同個體其發病機理不全相同[4]。近年越來越多的研究表明,機體的遺傳因素在SLE的易感性發病方面處于主導地位,且是多基因協調控制的,其發生、發展與否有賴于環境因素的“扳機”及促進作用[5]。目前已知多種遺傳基因與人類SLE易感性有關聯,其中FEN1基因的單核苷酸多態性與SLE有關[5],FEN1基因異常可造成細胞凋亡機制障礙,誘發自身抗體形成最終導致疾病的發生發展[6]。另外,全基因組掃描證實,在人類染色體上,存在著50多個與SLE相關的區段,在SLE家族中呈現不同程度的連鎖,提示其上可能存在著免疫相關的候選基因,包括人類白細胞抗原(HLA)Ⅰ類基因區 HLA-A、B、C、E、F、G、H、J和 X;Ⅱ類基因包括 HLA-DR、HLA-DQ、HLA·DP、TAP、LMP、DM;Ⅲ類基因包括編碼補體成分基因(C2、C4A、C4B和Factor B)、細胞因子基因[腫瘤壞死因子(TNF)、α、β基因]、淋巴毒素(LTA、LTB)、熱休克蛋白HSP70、T 細胞受體 (TCR)基因、TRIM、IL-4R、EGR1、KLRC1基因,人群16q12區(58.46 cM)OAZ基因和凋亡相關基因等[3,7-12],現就近幾年報道SLE的可能的分子病因及發病機理作一概述。
1 FcγRII和 FcγⅢR(1q23) 與 SLE
FcγR家族屬于免疫球蛋白超家族,其中FcγRIIa、FcγIIRb、FcγRⅢa 和 FcγRⅢb 存在基因多態性,被認為可能與SLE發病過程中免疫復合物清除有關,影響炎癥反應,導致個體對自身免疫性疾病有不同易感性,該基因多態性與SLE發病的關系在不同的種群中研究結果不一致,Karassa等通過Meta分析得出FcγRIIa-R131是SLE發病的重要危險因素,但不是狼瘡性腎炎的易感基因,且不同種族間沒有差異性分布;FcγRⅢa-158F與LN有關,但與SLE無關[13]。Chu等對日本、泰國、中國人群也作了FcγR基因多態性SLE發病關系的Meta分析卻得出FcγRIIa-R131與SLE和LN都無關的結論,而FcγRⅢa-158F與SLE有關,與LN發病無關[14]。這些研究提示FcγR基因確實是某些種群SLE和LN的易感基因,且種群間存在差別。為了更進一步的研究FcγR基因多態性在SLE發病中的作用,應進行多種人群采取更大規模的SLE遺傳學和FcγR各亞型功能的研究。
2 補體缺陷與SLE
在SLE病人中,約50% ~75%病人存在低補體血癥,很多患者的腎組織及皮膚組織亦可見大量的補體C1、C3和C4沉積[15]。研究發現,補體基因遺傳性缺陷可致的低補體血癥可能與SLE發病有關,尤其是經典途徑中早期補體成份(C1~C4)缺失或無功能時容易出現狼瘡樣癥狀,補體基因遺傳性缺陷可能導致補體缺乏溶血活性、膜攻擊復合物(MAC)缺陷、免疫復合物清除障礙、凋亡細胞清除障礙[15]。肝臟合成補體能力下降、嚴重的腎臟病造成補體丟失等因素也可與SLE補體異常有關。單補體成分C3、C4及總補體(CH50)活性在疾病活動期均可降低,其中任何一種補體成分缺陷均可使CH50降低,補體水平下降作為SLE活動性指標之一[16,17]。臨床上發現部分C3缺陷患者伴有膜增殖性腎小球腎炎、血尿或蛋白尿等表現,認為C3缺陷與一種稱為C3腎炎因子的物質有關,現已確定C3腎炎因子為一種抗C3bBb復合物上新抗原的特異性IgG抗體,它起到穩定C3bBb活性的作用[17]。在臨床上CH50測定是最簡單的補體活性篩選方法。Lyon等報道了1例C2缺陷年輕患者:初診表現為面部皮疹和頸部淋巴結腫大而無狼瘡和其他自身免疫病的證據,2年后檢測到抗核抗體和抗Sm抗體,最終診斷為SLE[18]。Jonsson等研究也證實了C2缺陷與SLE相關[19]。血漿C4是由C4A和C4B基因編碼的蛋白產物,C4A和C4B的功能不同,等位基因C4AQ0和SLE的相關證據強于C4BQ0與SLE,目前大量研究表明C4AQ0與SLE人群顯著相關,此等位基因大部分存在于DR3-B8單倍型[20]。總之,補體缺陷與SLE發病及疾病活動性關系密切,可在一定程度上反映病情的變化,CH50、C3和C4水平及其變化可以作為判斷SLE疾病活動性的重要指標,可為SLE患者生物靶向治療提供線索。
3 FEN1與SLE
FEN1基因位于人類染色體11q12,長1 144 bp,含有一個外顯子,編碼380個氨基酸,構成FEN1核酸酶的一級結構,是一種結構特異性多功能核酸酶[21],在DNA的多條代謝途徑、維護染色體的穩定及細胞凋亡過程中發揮著重要作用[22,23]。Li Zheng等建立FEN1基因E160D點突變小鼠模型,造成FEN1約90%以上的 EXO和GEN酶活性喪失,在FEN1基因E160D點突變小鼠腎臟及肺組織中,TUNEL-陽性的核酸產物計數較野生型小鼠顯著增高,表明FEN1基因E160D點突變可導致小鼠凋亡DNA產物增多。進一步分析發現,野生型小鼠成纖維細胞50%的凋亡細胞被轉運所需要的時間比FEN1基因E160D突變小鼠50%的凋亡細胞轉運時間少約24小時,后者對凋亡DNA產物的清除延遲,從而推測FEN1基因E160D點突變影響FEN1核酸酶功能,致使細胞凋亡過程中核小體DNA鏈不能有效分解,當大量的凋亡小體在機體組織中滯留時,作為具有免疫原性的靶抗原,構成危險信號對免疫系統造成威脅[24]。馮玲等對43例LN患者和26例健康者的外周血標本,采用全血基因組DNA柱式試劑盒提取DNA,直接PCR方法擴增FEN1基因片段,擴增后PCR產物應用基因測序方法檢測FEN1基因序列,并對測序結果與基因數據庫中FEN1基因進行比較,搜索可能的突變位點,結果發現LN患者正向測序中存在946位堿基C缺失突變(P=0.046),為該基因可能在LN的發病機制中的作用提供了一定的線索[25]。而我們初步研究中顯示該基因外顯子位點61563302、61563303堿基GT變為TC。我們認為FEN1在細胞周期或DNA損傷修復過程中會發生不同的翻譯后修飾,這些翻譯后修飾會抑制或促進它與不同蛋白之間的相互作用或者觸發其降解。因此,我們推測FEN1在DNA復制或修復過程中所起的作用是受一系列的翻譯后修飾調控的,翻譯后修飾的改變會使FEN1功能失常,引起DNA復制和修復缺陷并最終導致疾病的發生,為此下一步我們繼續應運基因操作技術就SLE患者該基因的多態性、表達產物等與疾病發生發展做更一步探討。
4 細胞凋亡缺陷與SLE
細胞凋亡是一種相關基因嚴格調控的細胞生理性死亡,它是許多生物進程的一個重要組成部分。一般認為,有序的細胞凋亡能阻止凋亡細胞的繼發性壞死和酶的釋放,從而抵制炎癥和免疫應答的發生,如果凋亡細胞清除障礙不能被及時清除,導致凋亡物質“進出”平衡失調,造成凋亡物質的累積,使細胞膜的完整性受損,細胞膜破裂,釋放出大量修飾過的細胞核和細胞質物質,成為維持自身免疫應答的反復性的刺激原,不斷攻擊已經致敏的免疫系統,破壞其對自身抗原的中樞和外周耐受,出現組織器官受損(圖1),進而引發炎癥和免疫應答,促使SLE的發生、發展[26]。SLE患者細胞凋亡異常及凋亡產物清除障礙導致核小體過度釋放,核小體與抗核小體抗體(AnuA)形成的免疫復合物沉積于腎小球上,AnuA在腎小球基底膜沉積是以固有細胞凋亡產生的核小體作為腎小球基底膜內的原位抗原或循環中核小體先與GBM相結合作為種植抗原,然后再在原位結合AnuA形成免疫復合物,這種沉積主要發生在上皮下,與膜性腎小球腎炎的發生發展有關,AnuA可能還參與Ⅰ型膠原的形成,從而加速腎小球的纖維化進程,另外LN患者體內免疫相關細胞,如淋巴細胞凋亡的增加,出現免疫系統紊亂,造成核小體過度釋放,血循環中出現的DNA以寡核小體形式存在,抗核小體抗體與抗DNA抗體有交叉反應,這兩者在LN的發生發展中都有著重要的作用[3,27,28]。因此,目前 AnuA 作為SLE患者腎臟損害的一項早期監測指標,將有助于早期干預治療,提高預后。目前已知促進凋亡的基因有Fas及Fas配體(Fas-L)基因、p53基因、c-myc基因等;抑制凋亡的基因有bc1-2基因、bc1-xL基因、Pim-1基因等,促進基因與抑制基因的相互平衡作用決定著細胞凋亡與否,其中起關鍵作用的是Fas基因、Fas-L基因及bc1-2基因。
Fas/Fas-L基因突變可導致細胞凋亡活化途徑受到抑制,使正常的免疫下調失效,自身反應性B細胞在體內積累增多,并產生致病性抗體,在SLE發病和病情進展中的重要作用已得到公認。Fas基因突變動物模型lpr/lpr小鼠在6月齡時發生SLE,而Fas-L基因突變gld/gld小鼠,也可以發生類似的疾病[29]。
bc1-2基因是細胞凋亡的抑制基因,其表達產物bc1-2蛋白是一種抑制多種細胞凋亡的蛋白,目前已有足夠的證據說明在SLE外周T細胞中存在bc1-2表達的增多,且在各亞群間無明顯差異,相反,B細胞則無此特性。與Fas基因相似,bc1-2基因異常也促進SLE的發生。首先,顯著增多的bc1-2蛋白一方面可使T細胞逃避細胞凋亡;另一方面它又促進T細胞增殖,導致反應性T細胞的大量產生及增殖。其次,bc1-2基因的突變也導致其表達的異常。動物實驗表明來自bc1-2基因突變鼠的T細胞對多種殺傷因素具有耐受性,這同樣可過度抑制T細胞的正常凋亡。另外bc1-2基因與c-myc基因的聯合表達也導致自反應性T細胞的出現及T細胞的過度增生[29]。最近我們收集了我省多家醫院的LN患者腎臟活檢組織標本40余例,對其中11例細胞凋亡檢測發現存在凋亡小體堆積,見圖2,其余標本正在實驗過程中。
總之,細胞凋亡研究是一個方興未艾的研究領域,是現代生物學的支柱學科之一。這方面的研究對包括SLE在內的多種疾病的發病機制提供了新的認識角度,為多種疾病的治療提供了新思路和新途徑。
5 微小干擾RNA與SLE
近年來,微小干擾(microRNA)作為一種非編碼RNA分子,已有證據表明其在先天免疫應答及炎癥因子的信號傳導過程中起著重要的負調節作用,SLE干擾素通路異常活化可能與某些miRNA異常表達有關,并且在SLE重要的致病通路中起重要作用,Toll樣受體7受到刺激后誘發漿細胞樣樹突狀細胞(plasmactoid dendritic cell pDC)內19miRNAs產生不同程度的表達,其中miR-155和miR-155*主要的產物通過相反作用調節pDC介導Ⅰ型干擾素的產生,miR-155*通過靶向IRAKM,促進Ⅰ型干擾素的產生,而miR-155通過靶向TAB2,抑制Ⅰ型干擾素的產生,且在不同階段發揮作用,此外,通過對miR-155和 miR-155*產生機制的研究,發現pDC自身分泌的Ⅰ型干擾素以及被激活的KHSRP蛋白可以在轉錄后水平反向調控miR-155和miR-155*的產生,這一結果解釋了來自于同一前體的miR-155*和miR-155卻能在不同的時間點被誘導的原因[30]。羅曉兵等研究小組采用候選基因測序的策略,選擇大樣本的病例-對照研究方法,在多個人群中發現miR-146a啟動子區域存在基因變異(rs57095329)與SLE顯著相關,攜帶疾病相關等位基因的個體,其miR-146a基因表達水平顯著低于對照組,miR-146a表達缺陷,影響SLE易感基因ETS1編碼表達的蛋白轉錄因子的結合能力,增加或者降低ETS1表達水平,導致miR-146a啟動子活性的改變,也證實ETS1與miR-146a相互間的影響作用,在SLE發病中發揮疊加效應,引起Ⅰ型干擾素通路過度活化,進而參與SLE疾病的發生發展[31]。這些研究例證了復雜性疾病中多個易感基因通過相互作用共同參與SLE疾病的發生,闡明了新型有效的調控Ⅰ型干擾素產生環節的措施,可能是SLE治療的潛在靶點。
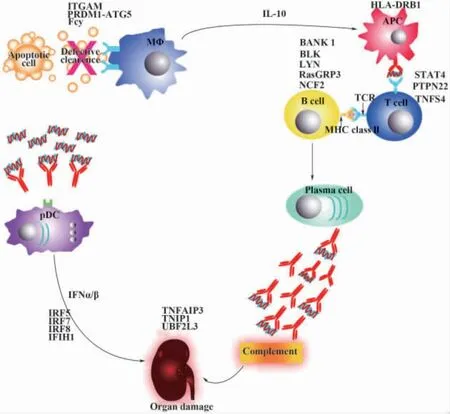
圖1 系統性紅斑狼瘡免疫系統損害Fig.1 The impaired immune system in patients with systemic lupus erythematosus(SLE)
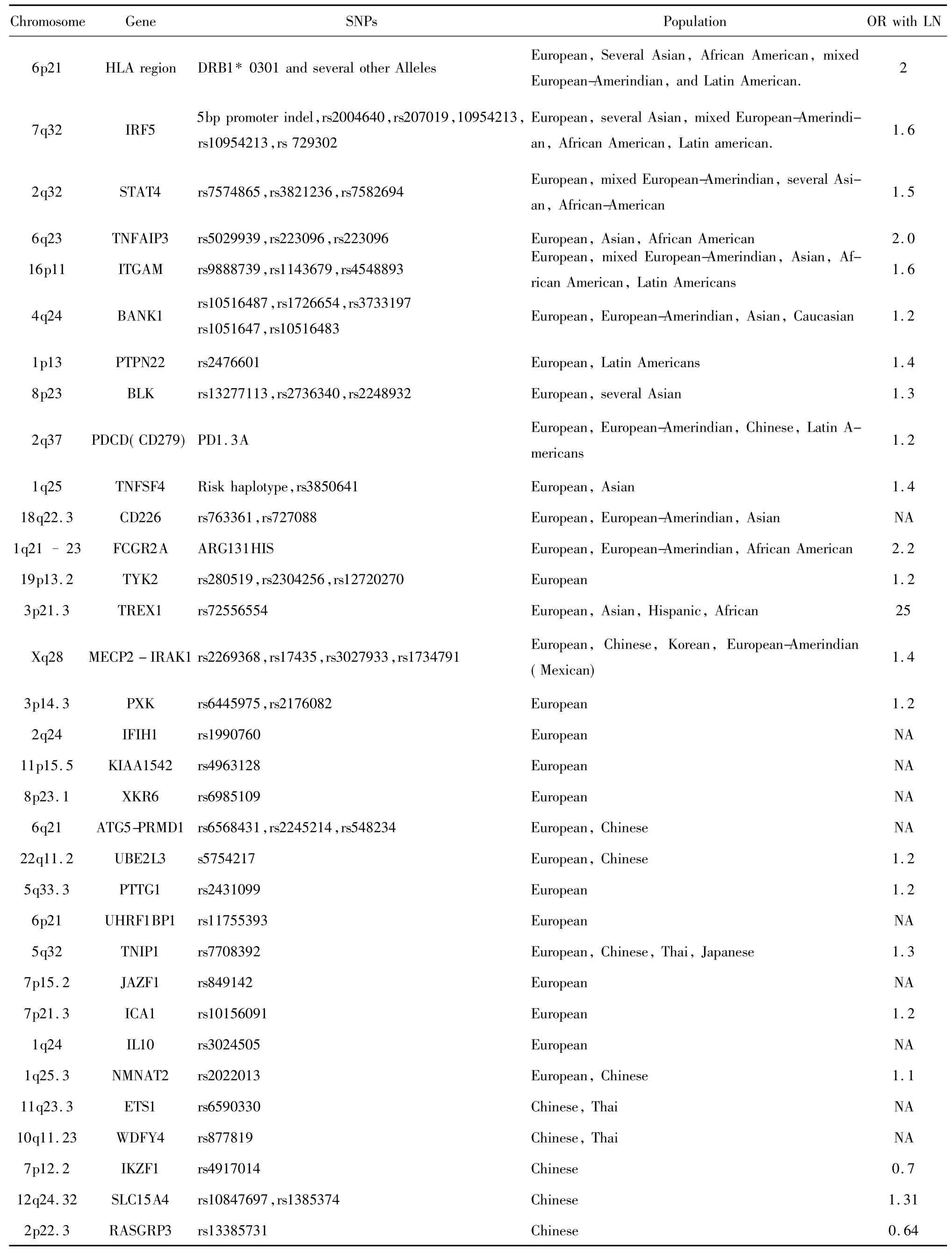
表1 SLE并發LN的易感基因Tab.1 Susceptibility genes in SLE associated with LN

圖2 LN患者腎臟組織細胞凋亡檢測圖Fig.2 LN kidney tissue apoptosis detection figure
6 狼瘡性腎炎易感基因
SLE基因學研究的發展促進了LN遺傳和易感基因的研究的持續發展,LN病因、發病機理十分復雜,而且不同個體其發病機理不全相同,和SLE一致亦存在種族差異性和遺傳異質性。以下列出近年來國內外研究已報道的不同基因多態性和LN的相關性[3],詳見表 1。
7 展望
隨著人類基因組計劃的完成,醫學基因組學研究以及計算機技術的快速發展,以當前的相關技術研究單基因遺傳病的致病基因已不再是難題。但單基因遺傳病的發病率通常比較低,嚴重危害人類健康和生活質量并困擾著科學研究策略的往往是一些多基因遺傳病,如SLE,對其易感基因的研究不僅對于發現易感人群、進行基因診斷有重要價值,而且對于探索疾病的發病機制和開辟新的治療途徑也有重大的意義。
目前SLE分子病因研究的困難在于:(1)多因素復雜性,十多個或者數十個甚至更多基因在不同環境下存在疊加交互作用。(2)遺傳異質性研究的疾病可能是存在不同遺傳背景表型集合。(3)所選擇的候選基因假陽性不一定和SLE疾病真實相關。(4)臨床疾病的診斷和分類的不確定性,因表型而被錯誤地作為患病的研究對象及未經隨訪證實的健康對照。(5)由于研究人群的遺傳結構不一致、樣本量不同、多重檢驗而導致的假陽性的結果會造成很多誤導。
近年來,有關SLE發病病因和診斷的研究雖然進展較大,但仍缺乏多中心、多對照的研究方法,各地科研側重點有所不同,難以從總體上把握其發病病因,尚有待于充分借鑒現代科學技術,加大基礎研究力度,力求從細胞、分子水平闡明SLE的病因,為臨床治療提供可靠的理論依據。糖皮質激素和免疫抑制劑聯合治療已經使SLE患者的預后及社會回歸率得到了很大的提高,但SLE仍然緩解率低下,部分重癥患者對激素及免疫抑制劑治療無效或療效欠佳,長期使用帶來的副作用也嚴重威脅了這些患者的健康,提高SLE的療效,改善患者的預后迫切需要我們對SLE發病機制的進行深入的理解,諸如抗CD20單抗(rituximab,利妥昔單抗)、抗CD5、抗IL-6、抗IL-10、干擾素等抗體,這些針對SLE發病機制中某一環節或影響其疾病進展的關鍵分子進行的選擇性靶向治療,儼然已經成為治療SLE的新方向,因此,繼續針對在SLE發病中起關鍵作用的遺傳流行病學、基因易感區域定位、克隆、轉錄、翻譯后的修飾等環節的深入研究,是最終提高SLE療效、控制病變進展、改善預后的根本途徑,基于此以生物技術為基礎的多種生物制劑的研發和應用將是控制SLE的有效措施。
1 Foster M H.T cells and B cells in lupus nephritis[J].In:Semin Nephrol,2007;27:47-58.
2 Goodnow C C,Sprent J,de St Groth B F et al.Cellular and genetic mechanisms of self tolerance and autoimmunity[J].Nature,2005;435(7042):590-597.
3 Alberto de Zubiria Salgado,Catalina Herrera-Diaz.Lupus nephritis:an overview of recent findings[J].Autoimmune Dis,2012;2012:849684.
4 Crispin J C,Liossis S N,Kis-Toth K et al.Pathogenesis of human systemic lupus erythematosus:recent advances[J].Trends Mol Med,2010;16(2):47-57.
5 Kim I,Hur Nw Fau-Shin H D,Park B L et al.Associations of DNase IV polymorphisms with autoantibodies in patients with[J].Rheumatology(Oxford),2008;47(7):996-999.
6 Zheng L,Dai H F,Zhou M et al.Fen1 mutations result in autoimmunity,chronic inflammation and cancers[J].Nat Med,2007;13(7):812-819.
7 Corporaal S,Bijl M F,Kallenberg C G.Familial occurrence of autoimmune diseases and autoantibodies in a Caucasian[J].Clin Rheumatol,2002;21(2):108-113.
8 Wong M,Tsao B P.Current topics in human SLE genetics[J].Springer Semin Immunopathol,2006;28(2):97-107.
9 Harley J B,Kelly J A,Kaufman K M.Unraveling the genetics of systemic lupus erythematosus[J].Springer Semin Immunopathol,2006;28(2):119-130.
10 Morel L.Genetics of human lupus nephritis[J].Semin Nephrol,2007;27(1):2-11.
11 Rizzo R,Hviid Tv Govoni M,Padovan M et al.HLA-G genotype and HLA-G expression in systemic lupus erythematosus:HLA-G as a putative susceptibility gene in systemic lupus erthematosus[J].Tissue Antigens,2008;71(6):520-529.
12 Ramos P S,Brown E E,Kimberly R P et al.Genetic factors predisposing to systemic lupus erythematosus and lupus nephritis[J].Semin Nephrol,2010;30(2):164-176.
13 Brambila-Tapia Aj,Davalos-Rodriguez IP.Fcgamma receptor polymorphisms and systemic lupus erythematosus[J].Rev Invest Clin,2009;61(1):66-72.
14 Niederer H A,Clatworthy M R,Willcocks L C et al.FcgammaRIIB,FcgammaRIIIB,and systemic lupus erythematosus[J].Ann N Y Acad Sci,2010;1183:69-88.
15 Pettigrew H D,Teuber S S,Gershwin M E.Clinical significance of complement deficiencies[J].Ann N Y Acad Sci,2009;1173:108-123.
16 Sarma J V.Ward P A.The complement system.Cell Tissue Res,2011;343(1):227-235.
17 Wen L,Atkinson J P,Giclas P C.Clinical and laboratory evaluation of complement deficiency[J].J Allergy Clin Immunol,2004;113(4):585-593;quiz 594.
18 Lyon V B,Nocton J J,Drolet B A et al.Esterly NB:Necrotic facial papules in an adolescent:C2 deficiency with eventual development[J].Pediatr Dermatol,2003;20(4):318-322.
19 Jonsson G,Truedsson L,Sturfelt G et al.Hereditary C2 deficiency in Sweden:frequent occurrence of invasive infection[J].Medicine(Baltimore),2005;84(1):23-34.
20 Graham R,Ortmann W,Rodine P et al.Specific combinations of HLA-DR2 and DR3 class II haplotypes contribute graded risk for disease susceptibility and autoantibodies in human SLE[J].European J Human Genetics,2007;15(8):8.
21 Ayyagari R,Gomes X V,Gordenin D A et al.Okazaki fragment maturation in yeast.I.Distribution of functions between FEN1[J].J Biol Chem,2003;278(3):1618-1625.
22 Liu Y,Kao H I,Bambara R A.Flap endonuclease 1:a central component of DNA metabolism[J].Annu Rev Biochem,2004;73(7):589-615.
23 Parrish J Z,Yang C,Shen B et al.CRN-1,a caenorhabditis elegans FEN-1 homologue,cooperates with CPS-6/Endo.G to promote apoptotic DNA degradation [J].Embo J,2003;22(13):3451-3460.
24 Zheng L,Zhou M,Chai Q et al.Novel function of the flap endonuclease 1 complex in processing stalled DNA[J].EMBO Rep,2005;6(1):83-89.
25 馮 玲,王 靜,王儉勤.FEN1基因突變在狼瘡性腎炎發病機制中的作用[J].國際泌尿系統雜志,2010;30(5):625-628.
26 Guerra S G,Vyse T J,Cunninghame Graham D S.The genetics of lupus:a functional perspective[J].Arthritis Res Ther,2012;14(3):211.
27 Kalaaji M,Fenton K A,Mortensen E S et al.Glomerular apoptotic nucleosomes are central target structures for nephritogenic[J].Kidney Int,2007;71(7):664-672.
28 王 靜,王志平.FEN1基因突變在狼瘡腎炎凋亡核小體清除機制中的作用[J].中華腎臟病雜志,2011;27(7):538-540.
29 Paunovic V,Carter N A,Thalhamer T et al.Immune complex-mediated co-ligation of the BCR with FcgammaRIIB results in[J].J Autoimmun,2012;39(4):332-346.
30 Zhou H,Huang X,Cui H et al.miR-155 and its star-form partner miR-155* cooperatively regulate type I[J].Blood,2010;116(26):5885-5894.
31 Luo X,Yang W,Ye D Q et al.A functional variant in microRNA-146a promoter modulates its expression and[J].PLoS Genet,2011;7(6):e1002128.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19