吳茱萸次堿改善溶血性磷脂酰膽堿誘導的內皮細胞縫隙連接細胞間通訊功能障礙
2013-12-08 06:56:12余艷榮彭維杰況海斌宋培源
中國藥理學通報 2013年11期
關鍵詞:效應
劉 勇,余艷榮,彭維杰,況海斌,徐 宏,宋培源,羅 丹
縫隙連接(gap junction,GJ)是由相鄰細胞膜上的縫隙連接蛋白構成的細胞間親水性通道。相鄰細胞通過GJ進行信息和物質能量的交流,即縫隙連接細胞間通訊(gap junction inter-cellular communication,GJIC),在維持細胞新陳代謝、調控增殖和分化等過程中起重要作用。目前發現縫隙連接蛋白(connexin,Cx)大約有20多種,其中血管內皮細胞表達Cx37、Cx40和 Cx43[1]。近年來的大量研究提示,血管壁Cx的類型、數目和空間分布的改變所致的GJIC功能障礙參與動脈粥樣硬化(atherosclerosis,AS)發生發展[2-3]。其中,Cx37 和 Cx40 被認為具有抗AS作用,在Apoe-/-小鼠模型,敲除Cx37和Cx40基因可加速高膽固醇飲食誘發的AS斑塊形成[4-5]。抗AS藥物他汀類可恢復人類一級血管中Cx37的表達,提示改善GJIC可能作為他汀類藥物的非降脂抗AS作用機制[6]。
吳茱萸次堿(rutaecarpine,Rut)為傳統中藥吳茱萸的主要活性成分,具有廣泛的心血管保護效應[7]。前期大量實驗證據均表明,Rut的主要心血管效應(如對豚鼠心房的正性肌力和正性頻率作用,舒血管和降壓作用,以及心肌保護作用等等)均與激活辣椒素受體(transient receptor potential vanilloid 1,TRPV1)有關[8]。TRPV1屬瞬時感受器電位離子通道(TRP)超家族成員,廣泛分布于感覺神經元末梢,我們已確證人臍靜脈內皮細胞(HUVECs)膜表面也存在TRPV1[9]。國外學者發現[10],Rut可抑制AS因子LIGHT誘導的單核細胞遷移,提示其具有潛在的抗AS效應。已知血管內皮損傷是AS血管病變的始動環節,保護內皮是抗AS的重要策略。我們前期研究發現,Rut明顯降低Ox-LDL的主要活性成分溶血性磷脂酰膽堿(lysopbosphatidylcholine,LPC)誘導的內皮細胞凋亡,預先給予TRPV1競爭性拮抗劑CAPZ可以取消其效應,提示Rut通過激活TRPV1發揮內皮保護效應[11]。本實驗在LPC誘導的內皮損傷模型進一步確定Rut的內皮保護作用,觀察其對內皮Cx37、Cx40和GJIC的影響,并探討Rut對GJ的調控機制是否涉及TRPV1途徑。
1 材料與方法
1.1 藥品與試劑 人臍靜脈內皮細胞株HUVEC-12(北京大學醫學院腫瘤研究所提供,源自ATCC細胞庫),人單核細胞株THP-1(中科院上海生化細胞所提供,源自ATCC細胞庫);吳茱萸次堿(純度>99%,上海君拓生物技術有限公司),LPC、Lucifer Yellow(熒光黃染料)、CAPZ(Sigma,USA),SYBR@Premix EX TaqTM(寶生物工程有限公司),TransZol細胞裂解液,RT-PCR Super Mix(北京全式金生物技術有限公司),兔抗人 Cx37和兔抗人 Cx40抗體(ab83788 和 ab38580,Abcam),Goat anti-rabbit IgG-(H+L)/HRP(北京中杉金橋有限公司),NO檢測試劑盒(南京建成生物工程研究所),活性氧(ROS)檢測試劑盒(江蘇碧云天生物技術有限公司)。
1.2 儀器 ABI 7500熒光定量 PCR儀(ABI,USA),流式細胞儀(FCM)(Becton Dickinson,USA),熒光顯微鏡(Olympus,日本),酶聯免疫檢測儀(BIO-RAD,USA)。
1.3 實驗分組 HUVEC-12解凍復蘇后,取3~8代細胞進行實驗。將細胞稀釋至1×108·L-1后接種至96孔板或6孔板中,待細胞80%融合時換成含1%小牛血清的DMEM培養基培養24 h,使細胞同步化,然后分別加入不同的處理因素:①Control組:培養基中不加任何處理因素;②LPC損傷組:加入LPC(終濃度為10 mg·L-1)孵育內皮細胞24 h;③-⑤+Rut(高、中、低)保護組:分別加入終濃度為 1.0、0.3、0.1 μmol·L-1的吳茱萸次堿孵育內皮細胞10 min后,加入LPC繼續孵育24 h;⑥+CAPZ+Rut(H)組:用 TRPV1受體阻斷劑 CAPZ(10 μmol·L-1)預先處理細胞10 min后,加入吳茱萸次堿(1.0 μmol·L-1)孵育內皮細胞 10 min,然后加入LPC繼續孵育24 h。
1.4 Real-time PCR 檢測 Cx37、Cx40 mRNA 的表達(SYBR Green染料法)Trizol提取內皮細胞總RNA,按RT-PCR試劑盒方法逆轉錄。所用引物和擴增產物如下:Cx37(150bp):5'-GGCACCTATGTCGCCAGTG-3'(上游)和5'-GCGAGAGACAAAGCAGTCCA-3'(下游);Cx40(137bp):5'-TCCTGGAGGAAGTACACAAGC-3'(上游)和5'-ATCACACCGGAAATCAGCCTG-3'(下游);GAPDH(102bp):5'-AAGGTGAAGGTCGGAGTCAAC-3'(上游)和 5'-GGGGTCATTGATGGCAACAATA-3'(下游)。每 20 μl PCR 反應體系中含:cDNA 2 μl,SYBR Premix Ex Taq(2 × )10 μl,上下游引物各 0.4 μl,ROX Reference DyeⅡ(50 × )0.4 μl,RNase-free Water 6.8 μl。擴增條件為:預變性95℃ 30 s,變性95℃,5 s,退火60℃ 34 s,40 循環。以 2-△△Ct法進行相對定量。
1.5 Western blot檢測 Cx37、Cx40 的蛋白水平提取細胞蛋白,經BCA法蛋白定量,聚丙烯酰胺凝膠電泳,半干轉至PVDF膜,5%脫脂奶粉室溫封閉2 h,加入一抗(1 ∶1 000稀釋),4℃孵育過夜,TBST室溫漂洗3次后加入二抗(1∶5 000),漂洗后將膜放入裝有ECL混合液的封閉袋中置于X線暗盒中,壓片曝光1~20 min,顯影、定影。將X線片掃描,使用Quantity One分析軟件進行灰度分析,以β-actin的密度為內參計算相對密度,結果表示為實驗組相對于正常對照組相對密度的倍數差異。
1.6 劃痕負載試驗測定 GJIC功能 Lucifer Yellow(熒光黃)染料不能通過正常細胞細胞膜,只能在劃傷瞬間進入被劃傷細胞內,并通過縫隙連接傳遞給相鄰正常細胞,故熒光黃在細胞間傳遞的范圍可反映GJIC功能。待細胞生長融合后,吸除細胞培養液,加入1 ml 0.5 g·L-1的LY熒光染料,在培養皿細胞表層劃3道劃痕,室溫孵育3 min后吸出染料,PBS漂洗3次,熒光顯微鏡下觀察染料在劃痕垂直方向上的遷移范圍。
1.7 MTT水平檢測內皮細胞活力 將內皮細胞消化后制成單細胞懸液,以每孔103~104個細胞接種于96孔培養板中,每孔體積200 μl,同時設置調零孔(只加入等體積的無細胞培養液)。培養結束后,每孔加入 MTT 溶液(5 g·L-1)20 μl,37℃繼續孵育4 h,終止培養,小心吸棄孔內培養上清液。每孔加入150 μl DMSO,振蕩10 min,使結晶物充分溶解。用酶標儀測定490 nm處各孔的吸光度值。細胞活力(OD)=樣品吸光度值-調零孔吸光度值。
1.8 細胞培養液中NO水平的測定 吸取細胞培養液,2 000×g離心5 min,留取上清液,按試劑盒操作說明檢測。硝酸還原酶法測定細胞培養液中NO的含量,NO(μmol·L-1)=樣品消光值 ÷ 標準管消光值×100。
1.9 細胞ROS生成 (流式探針法)DCFH-DA本身沒有熒光,進入細胞后可被胞內的酯酶水解生成DCFH。細胞內的 ROS可進一步氧化無熒光的DCFH生成有熒光的DCF,檢測DCF的熒光強度可以反映細胞內ROS水平。制備單細胞懸液,洗滌后以 500 μl培養基重懸,加入 10 μmol·L-1DCFHDA熒光探針,37℃避光孵育20 min,洗滌細胞后用300 μl PBS重懸細胞,流式細胞儀檢測熒光強度。
1.10 內皮-單核細胞黏附率的檢測 單核細胞THP-1稀釋至109·L-1,加入6孔板的內皮細胞中(每孔1 ml),37℃孵育30 min后用PBS洗滌3遍,去除未黏附細胞,鏡下計數黏附的單核細胞數。倒置高倍顯微鏡下每孔隨機選擇6個視野,計數黏附的單核細胞數目。
2 結果
2.1 吳茱萸次堿對LPC損傷內皮細胞中Cx37和Cx40 mRNA水平的影響 Real-time PCR結果顯示(Fig 1),LPC孵育內皮細胞后,Cx37和Cx40 mRNA的水平均明顯下降,不同濃度的吳茱萸次堿可不同程度地上調兩者水平,預先給予TRPV1受體阻斷劑CAPZ(10 μmol·L-1)可取消吳茱萸次堿的這一效應(P<0.01)。
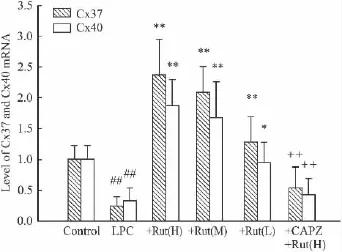
Fig 1 Effect of Rut on Cx37 and Cx40 mRNA expression in LPC treated HUVECs(Cx37 and Cx40 mRNA was detected by Real-time PCR)(ˉ ± s,n=3)
2.2 吳茱萸次堿對LPC損傷內皮細胞中Cx37和Cx40蛋白水平表達的影響 Western blot結果如Fig 2所示,LPC明顯抑制內皮細胞Cx37和Cx40的蛋白表達,吳茱萸次堿則可恢復兩者的表達水平,該效應可被CAPZ所阻斷(P<0.01)。
2.3 吳茱萸次堿對LPC誘導GJIC功能障礙的影響 熒光黃染料沿劃痕處的垂直傳遞范圍可反映GJIC功能,如Fig 3所示,LPC明顯抑制熒光黃染料沿劃痕處的垂直傳遞,吳茱萸次堿(1.0 μmol·L-1)能在一定程度上恢復LPC所致的GJIC功能障礙,該效應可被CAPZ所阻斷。

Fig 2 Effect of Rut on level of Cx37 and Cx40 proteinin LPC treated HUVECs(ˉ±s,n=3)
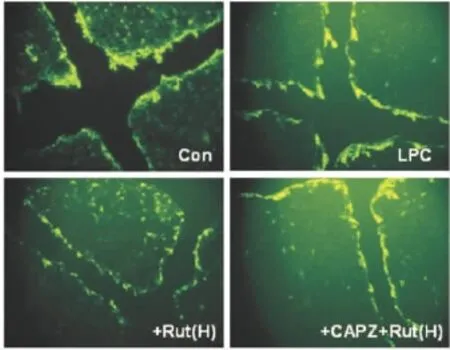
Fig 3 Effect of Rut on function of GJICin LPC treated HUVECs
2.4 吳茱萸次堿對LPC誘導的內皮細胞活力的影響 如Tab 1所示,LPC明顯降低內皮細胞的活力,吳茱萸次堿(1.0、0.3、0.1 μmol·L-1)能劑量依賴性地抑制LPC所致的內皮細胞活力下降,預先給予CAPZ可取消這種保護作用(P<0.01)。
2.5 吳茱萸次堿對LPC誘導的內皮細胞NO和ROS生成的影響 如Tab 1所示,LPC可抑制內皮保護因子NO的釋放,促進氧化應激產物ROS的生成(P<0.01)。不同劑量的吳茱萸次堿能劑量依賴性地促進NO生成,降低ROS的水平。這些內皮保護效應均可被CAPZ所阻斷(P<0.01)。
2.6 吳茱萸次堿對LPC誘導內皮-單核細胞黏附的影響 如Tab 1所示,正常內皮細胞可黏附少量單核細胞(108±5)個,LPC孵育內皮細胞可明顯增加單核細胞黏附數量(271±8)個,不同濃度的吳茱萸次堿(1.0、0.3、0.1 μmol·L-1)均能明顯抑制內皮-單核細胞黏附(P<0.01),該效應均可被CAPZ所取消(P<0.01)。
Tab 1 Effect of Rut on endothelial cell damage induced by LPC(ˉ±s,n=6)
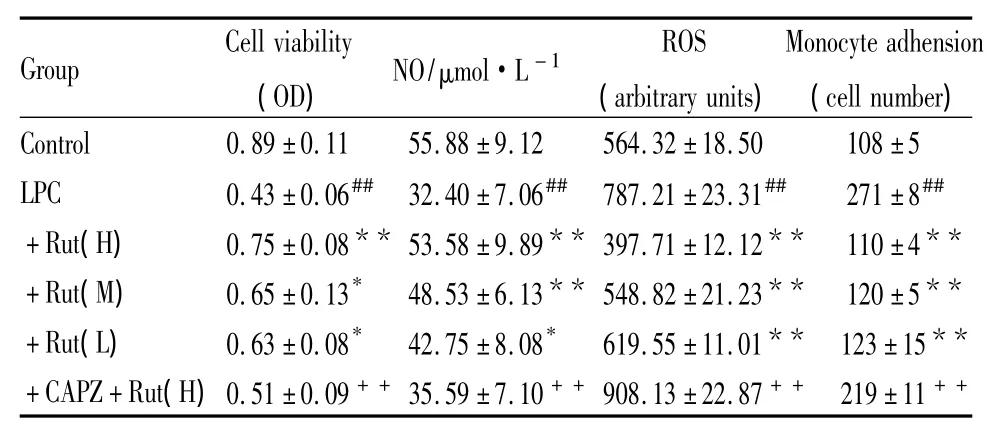
Tab 1 Effect of Rut on endothelial cell damage induced by LPC(ˉ±s,n=6)
##P<0.01 vs control;*P<0.05,**P<0.01 vs LPC;++P<0.01 vs+Rut(H).
Group Cell viability(OD)NO/μmol·L -1 ROS(arbitrary units)Monocyte adhension(cell number)Control 0.89 ±0.11 55.88 ±9.12 564.32 ±18.50 108±5 LPC 0.43 ±0.06## 32.40 ±7.06## 787.21 ±23.31## 271±8##+Rut(H) 0.75±0.08** 53.58±9.89** 397.71±12.12** 110±4**+Rut(M) 0.65±0.13* 48.53±6.13** 548.82±21.23** 120±5**+Rut(L) 0.63 ±0.08* 42.75 ±8.08* 619.55 ±11.01** 123±15**+CAPZ+Rut(H)0.51 ±0.09++35.59 ±7.10++908.13 ±22.87++ 219±11++
3 討論
吳茱萸次堿是蕓香科植物吳茱萸的干燥將近成熟果實中的一種吲哚喹唑啉類生物堿,具有廣泛的心血管效應。最近有研究報道,吳茱萸次堿可抑制單核細胞遷移,提示具有抗AS效應[10]。血管內皮結構和功能障礙是AS血管病變的始動因素,進而循環中單核細胞與損傷內皮黏附,遷移至內膜并最終發展為泡沫細胞,是AS斑塊進展的關鍵環節。保護血管內皮,抑制單核細胞黏附是防治AS斑塊進展的重要策略。高脂血癥、脂質代謝失常是AS的重要病因,其中氧化性低密度脂蛋白(Ox-LDL)在AS發生發展中起重要作用。LPC是Ox-LDL的主要活性成分,可誘導細胞膜脂質過氧化而損傷內皮細胞,并促進單核細胞黏附和泡沫細胞形成。因此,本實驗采用LPC孵育內皮細胞建立模擬AS內皮損傷模型。我們前期在培養的HUVEC發現,吳茱萸次堿可抑制LPC誘導的內皮細胞凋亡和TNF-α的釋放[11]。本實驗進一步證實,吳茱萸次堿對LPC誘導的內皮損傷具有明顯的保護效應,表現為升高內皮細胞活力,促進內皮保護因子NO生成,抑制ROS生成和單核細胞黏附。
近年來的大量研究提示,血管壁縫隙連接蛋白Cx的類型、數目和空間分布的改變所致的GJIC功能障礙參與AS發生發展[12]。目前對Cx和AS關系的研究主要集中在Cx37、Cx40和Cx43,前兩者可能有抗 AS效應,而后者有促 AS作用[2]。在Apoe-/-小鼠模型,敲除Cx37基因可加速高膽固醇飲食誘發的AS斑塊形成[4];選擇性敲除內皮Cx40則升高黏附分子VCAM-1的表達,促進白細胞黏附,明顯增加AS斑塊面積[5]。本實驗主要觀察吳茱萸次堿對有抗AS作用的內皮Cx37、Cx40的mRNA和蛋白的表達的影響。實驗結果發現:LPC損傷組與對照組相比,Cx37、Cx40的mRNA和蛋白水平均明顯下調,GJIC功能也明顯抑制。這和整體水平的研究結果一致,Kwak等[13]在小鼠和人AS斑塊中發現,隨粥樣斑塊進展,內皮逐漸失去Cx37和Cx40的表達。這些結果提示,Cx37和Cx40表達下調所導致的GJIC功能障礙可能是AS血管病變的重要機制之一。目前的研究表明,Cx37和Cx40不僅可調節eNOS的表達和活性,進而調節NO的生成而保護血管內皮[14],還可通過抑制單核細胞與內膜黏附而發揮抗AS作用。我們實驗發現,不同劑量的吳茱萸次堿(1.0、0.3、0.1 μmol·L-1)組與 LPC 損傷組相比,Cx37、Cx40的mRNA和蛋白表達均明顯上調,并明顯改善GJIC功能。這提示Rut的內皮保護和抑制單核細胞黏附作用可能與上調Cx37和Cx40表達而改善GJIC功能有關。預先給予TRPV1阻斷劑CAPZ可取消Rut(H)組的內皮保護作用及對Cx37、Cx40的水平、和GJIC功能的調節作用,表明Rut的內皮保護作用及改善LPC誘導的GJIC障礙均與激活TRPV1有關。
TRPV1為一種配體門控的非選擇性陽離子通道,可被辣椒素、高溫和氫離子等理化因素激活,導致2價陽離子(主要是Ca2+)內流。目前,關于TRPV1對GJIC的調控作用尚未見報道,我們推測可能與抗氧化途徑有關。氧化應激是AS發生發展的重要病理生理機制,其中來源于NADPH氧化酶的ROS起主要作用。細胞內ROS可通過氧化Cx或者改變其所在膜環境等誘發Cx構象變化,從而抑制GJIC。在單核細胞和心肌細胞的研究發現,吳茱萸次堿可抑制NADPH氧化酶活性,減少ROS生成,從而產生抑制單核細胞遷移和心肌保護作用[10]。有證據表明,TRPV1參與調節NADPH氧化酶活性和ROS的生成,其機制與調節胞內Ca2+水平有關[15]。本實驗發現,吳茱萸次堿可抑制LPC損傷內皮中ROS的生成,而CAPZ阻斷TRPV1可取消這一效應。我們推測,吳茱萸次堿激活TRPV1后,可能通過抑制NADPH氧化酶活性和ROS生成,進而改善GJIC。
綜上所述,本研究結果表明,吳茱萸次堿可減輕LPC誘導的內皮細胞損傷和單核細胞黏附,恢復內皮Cx37和Cx40的表達而改善GJIC,其機制與激活TRPV1有關。
[1] Brisset A C,Isakson B E,Kwak B R.Connexins in vascular physiology and pathology[J].Antioxid Redox Signal,2009,11(2):267-82.
[2] Pfenniger A,Chanson M,Kwak B R.Connexins in atherosclerosis[J].Biochim Biophys Acta,2013,1828(1):157 -66.
[3] 陳 敏,蔣麗萍,洪 濤.縫隙連接蛋白在動脈粥樣硬化形成和防治中的作用[J].中國藥理學通報,2010,26(10):1271-4.
[3] Chen M,Jiang L P,Hong T.The role of gap junction connexin in the formation,precaution and treatment of atherosclerosis[J].Chin Pharmacol Bull,2010,26(10):1271 -4.
[4] Wong C W,Christen T,Roth I,et al.Connexin37 protects against atherosclerosis by regulating monocyte adhesion[J].Nat Med,2006,12(8):950 -4.
[5] Chadjichristos C E,Scheckenbach K E,van Veen T A,et al.Endothelial-specific deletion of connexin40 promotes atherosclerosis by increasing CD73-dependent leukocyte adhesion[J].Circulat,2010,121(1):123 -31.
[6] Pfenniger A,Wong C,Kwak B R,et al.Shear stress modulates the expression of the atheroprotective protein Cx37 in endothelial cells[J].J Mol Cell Cardiol,2012,53(2):299 -309.
[7] 胡長平,李元建.吳茱萸堿和吳茱萸次堿的藥理學研究進展[J].中國藥理學通報,2003,19(10):1084-7.
[7] Hu C P,Li Y J.Research progress in pharmacological actions of evodiamine and rutaecarpine[J].Chin Pharmacol Bull,2003,19(10):1084-7.
[8] Peng J,Li YJ.The vanilloid receptor TRPV1:role in cardiovascular and gastrointestinal protection [J].Eur J Pharmacol,2010,627(1-3):1-7.
[9] Luo D,Peng W J,Li Y J,et al.Transient receptor potential vanilloid 1-mediated expression and secretion of endothelial cell-derived calcitonin gene-related peptide[J].Regul Pept,2008,150(1-3):66-72.
[10] Heo S K,Yun H J,Yi H S,et al.Evodiamine and rutaecarpine inhibit migration by LIGHT via suppression of NADPH oxidase activation[J].J Cell Biochem,2009,107(1):123 -33.
[11]邱模昌,余艷榮,羅 丹,等.吳茱萸次堿對內皮細胞損傷的保護效應及機制[J].時珍國醫國藥,2013,24(3):580-2.
[11] Qiu M C,Yu Y R,Luo D,et al.Protective effect of rutaecarpine(RUT)on human umbilical vein endothelial cell(HUVEC)injury[J].Lishizhen Med Mat Med Res,2013,24(3):580 -2.
[12] Morel S,Burnier L,Kwak B R.Connexins participate in the initiation and progression of atherosclerosis[J].Semin Immunopathol,2009,31(1):49 -61.
[13] Kwak B R,Mulhaupt F,Veillard N,et al.Altered pattern of vascular connexin expression in atherosclerotic plaques[J].Arterioscler Thromb Vasc Biol,2002,22(2):225 -30.
[14] Looft-Wilson R C,Billaud M,Johnstone S R,et al.Interaction between nitric oxide signaling and gap junctions:Effects on vascular function[J].Biochim Biophys Acta,2012,1818(8):1895-902.
[15] Schilling T,Eder C.Importance of the non-selective cation channel TRPV1for microglial reactive oxygen species generation[J].J Neuroimmunol,2009,216(1 -2):118 -21.
猜你喜歡
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
科學大眾(2020年17期)2020-10-27 02:49:10
紅土地(2018年11期)2018-12-19 05:10:56
意林·全彩Color(2018年9期)2018-11-13 22:49:38
中學物理·高中(2016年12期)2017-04-22 11:53:03
中國衛生(2016年4期)2016-11-12 13:24:14
中國衛生(2014年4期)2014-12-06 05:57:14
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
