洞頭海域網采浮游植物的月際變化
2013-12-16 08:19:08朱旭宇曾江寧江志兵廖一波陳全震
生態學報 2013年11期
關鍵詞:優勢
朱旭宇,黃 偉,曾江寧,江志兵,杜 萍,廖一波,張 鵬,陳全震,*
(1.國家海洋局第二海洋研究所,國家海洋局海洋生態系統與生物地球化學重點實驗室,杭州 310012;2.國家海洋局南通海洋環境監測中心站,南通 226005;3.浙江省海洋水產養殖研究所,溫州 325000)
浮游植物是海洋生態系統中最主要的初級生產者,其在海洋生態系統的能量流動和物質循環中起至關重要的作用。水動力[1]、溫度[2]、鹽度[3]、營養鹽[4-5]、競爭[6]和攝食[7]等對浮游植物群落具有重要的調控作用,而浮游植物又可通過食物鏈最終影響生態系統的穩定,具有較好的環境指示意義。
洞頭列島(121°10′E,27°50′N)位于甌江口外約50 km的浙江省東南海域。因其受臺灣暖流、閩浙沿岸流和甌江徑流的影響[8-10],營養鹽輸入豐富,是浙江沿海主要漁場之一[11]。近20年來,浙江沿岸區域和本島經濟的快速發展,生活污水、水產養殖、農業施肥和工業生產等人類活動造成氮、磷營養鹽入海通量急劇增加,最終導致該海域頻繁爆發東海原甲藻(Prorocentrum donghaiense)和微小原甲藻(P.minimum)等有害赤潮[12]。因此,研究洞頭海域浮游植物群落對環境變化的響應具有重要科學意義,但目前鮮有該海域浮游植物及其環境因子影響的報道[13-14]。本研究根據在洞頭島鄰近海域進行的周年(月際)連續監測結果,并結合歷史資料,分析了浮游植物群落的物種組成、優勢種和多樣性指數與主要環境因子的關系。研究結果可為深入了解環境變化對該海域浮游植物群落的影響提供基礎資料。
1 研究區域和方法
1.1 研究區域
洞頭海區是我國強潮區之一,潮汐屬正規半日潮,該海區潮流主要為往復流[15],因此,可以認為該海區水體混合較均勻。于2010年8月至翌年7月每月中旬對洞頭海域進行一次理化因子與浮游植物的現場調查,共設7個平行采樣點(圖1)。
1.2 樣品采集與分析
按照《海洋調查規范》[16],浮游植物樣品用淺III浮游生物網(網口直徑37 cm,網口面積0.1 m2,網目為77 μm)自底至表作垂直拖網采集,并用Hydro-bios流量計記錄濾水量。樣品用中性甲醛固定,經濃縮后用Nikon E200顯微鏡觀察、鑒定和計數。同時采集表層(0.5 m)水樣,黑暗冷藏條件下帶回實驗室,硝酸鹽、亞硝酸鹽、銨鹽、活性磷酸鹽和硅酸鹽的分析方法按《海洋監測規范》[17]進行。其中采用鋅-鎘還原法采用重氮-偶氮法,采用次溴酸鈉氧化法,采用硅鉬黃法采用磷鉬藍法。
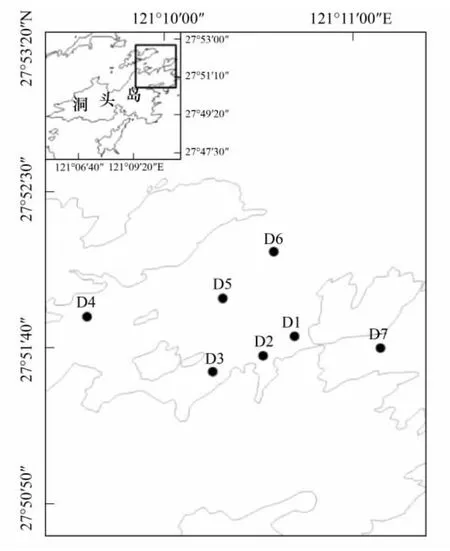
圖1 洞頭海域調查站位Fig.1 Sample stations in Dongtou Island
1.3 數據處理
采用以下公式[18-19]分別計算:

式中,S為樣品中的種類總數,N為樣品中的總個體數,ni為樣品中第i種的個體數,fi為該種在各樣品中出現的頻率,a、b分別為A、B月份所具有的物種數,c為兩月共有的物種數。
文中除種類數外的其他數據以平均值±標準差(Mean±S.D.)表示。用CANOCO軟件分析浮游植物與環境因子的關系,為簡化分析數據,避免種類檢出的偶然性,本文用于排序的物種為每月出現的優勢種。物種矩陣和環境矩陣的各項參數(除pH值外)都經過log(x+1)轉換,將物種數據進行去趨勢對應分析(DCA)后,發現軸長>4,故選擇典范對應分析(CCA)進行物種-環境分析[20],繪圖采用SigmaPlot 9.0軟件進行。
2 結果
2.1 理化因子
2.1.1 鹽度、溫度和pH
調查期間鹽度和水溫月變化較明顯(圖2)。鹽度在7月最高,為30.3,12月最低,為26.8;從季節來看,夏季(30.1)>春季(28.3)>秋季(28.0)>冬季(26.8)。水溫變化趨勢與鹽度類似,8月最高,為29.9℃,2月最低,為7.0℃;從季節來看,夏季(28.2℃)>秋季(21.1℃)>春季(15.9℃)>冬季(9.9℃)。pH全年變化不大,從季節來看,夏季(8.19)>冬季(8.16)>春季(8.14)>秋季(8.13)。
2.1.2 營養鹽
調查期間磷酸鹽濃度全年變化幅度較大(圖3),12月濃度最高,為1.43 μmol/L,5—9月較低,最低值出現在8 月,僅 0.58 μmol/L;從季節來看,冬季(1.31 μmol/L)>秋季(1.07 μmol/L)>春季(1.00 μmol/L)>夏季(0.61 μmol/L)。硅酸鹽濃度在大多月份較高,其中10月最高,為55.05 μmol/L,8月最低,為20.57 μmol/L(圖3b);從季節來看,冬季(42.06 μmol/L)>秋季(41.18 μmol/L)>春季(33.22 μmol/L)>夏季(24.81 μmol/L)。亞硝酸鹽和銨鹽濃度全年均較低(圖3),亞硝酸鹽夏季(1.73 μmol/L)>春季(1.38 μmol/L)>秋季(0.58 μmol/L)>冬季(0.14 μmol/L),銨鹽夏季(2.26 μmol/L)>秋季(2.19 μmol/L)>春季(0.90 μmol/L)>冬季(0.87 μmol/L)。硝酸鹽濃度在本海域較高(圖3),僅在7—9月稍低,8月最低,為11.32 μmol/L;從季節來看,冬季(88.81 μmol/L)>春季(86.23 μmol/L)>秋季(55.69 μmol/L)>夏季(27.77 μmol/L)。溶解無機氮(DIN)變化趨勢與硝酸鹽一致(圖3),8月最低,為15.02 μmol/L;從季節來看,冬季(89.82 μmol/L)>春季(88.51 μmol/L)>秋季(58.46 μmol/L)>夏季(31.76 μmol/L)。
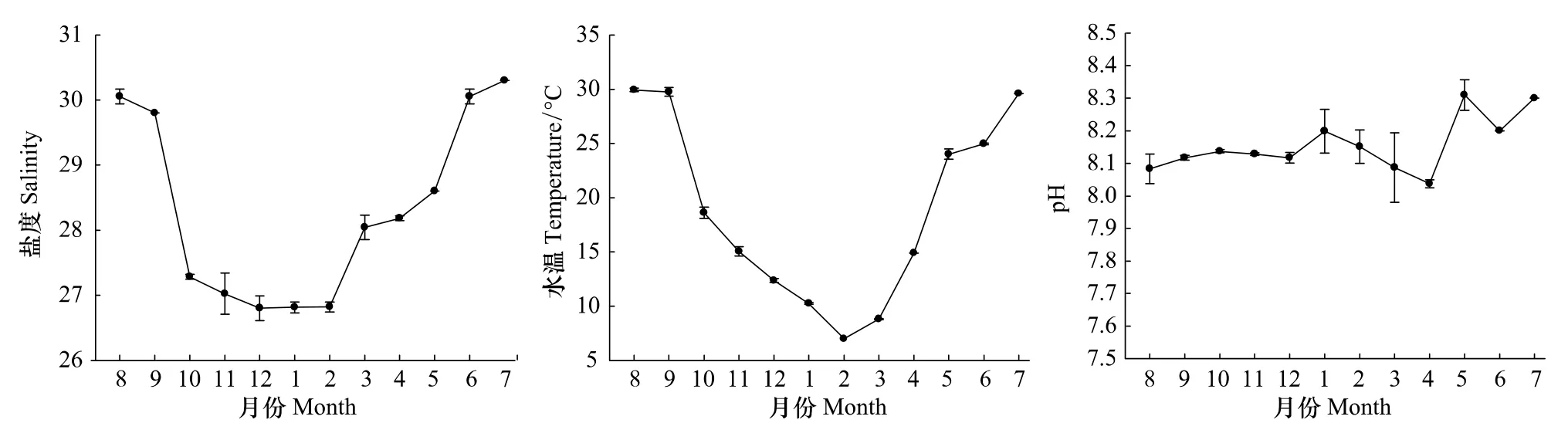
圖2 2010—2011年洞頭海域表層海水鹽度、溫度和pH月變化(平均值±標準差)Fig.2 Monthly variation of salinity,temperature and pH of surface waters in Dongtou Island during 2010—2011(Mean±S.D.)
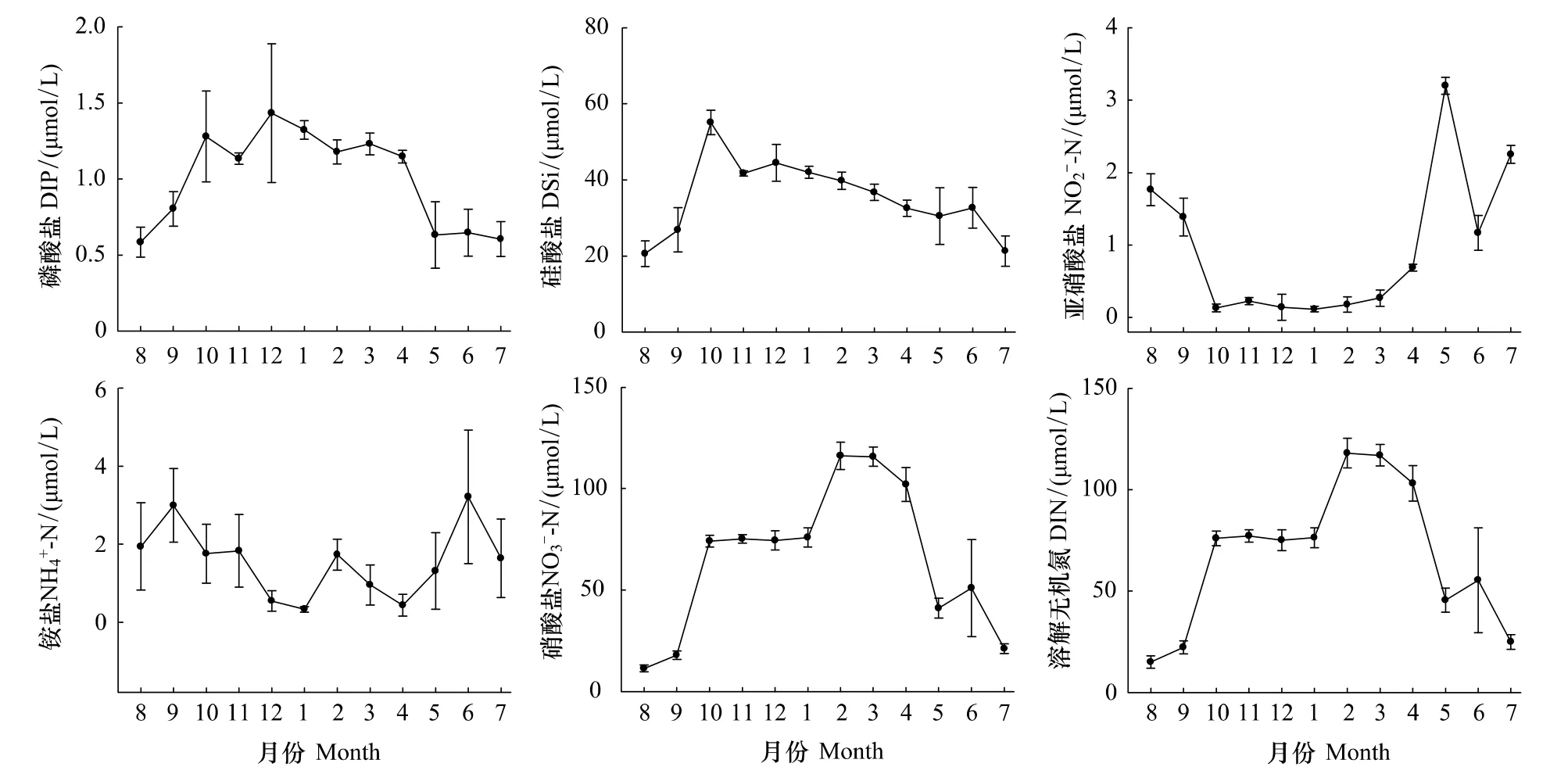
圖3 2010—2011年洞頭海域表層海水營養鹽濃度月變化Fig.3 Monthly variation of nutrient concentration of surface waters in Dongtou Island during 2010—2011
調查海域營養鹽比例月變化也較明顯(圖4)。N∶P除8和9月較低外,其他各月均較高,其中8月最低,為 25.7∶1,2 月最高,為 100.2∶1;從季節來看,春季(85.6∶1)>冬季(70.1∶1)>秋季(51.7∶1)>夏季(50.1∶1)。Si∶P范圍在28.3—50.4之間,其中4月最低,6月最高;從季節來看,夏季(40.3∶1)>秋季(37.7∶1)>春季(35.4∶1)>冬季(32.2∶1)。Si∶N變化范圍在0.3—1.4之間,其中8月最高,3和4月較低;從季節來看,夏季(0.9∶1)>秋季(0.8∶1)>冬季(0.5∶1)>春季(0.4∶1)(圖 4)。
2.2 浮游植物群落結構
2.2.1 種類組成
網采浮游植物共鑒定出187種,隸屬于硅藻(143種,占76.5%)、甲藻(40種,占21.4%)、金藻(2種,占1.1%)、藍藻(1種,占0.5%)和綠藻(1種,占0.5%)等5個門類。硅藻中角毛藻屬Chaetoceros的種類數最高,有26種;圓篩藻屬Coscinodiscus次之,有16種。甲藻中原多甲藻屬Protoperidinium的種類最多,有14種;角藻屬Ceratium次之,有10種。共發現72種赤潮種,占浮游植物種類數的38.5%,其中硅藻44種,甲藻25種,藍藻1種,金藻2種。各月浮游植物門類分布如圖5所示,9月種類數最高,有97種,6月最低,僅38種;從季節來看,秋季(78種)>夏季(72種)>冬季(69種)>春季(59種)。
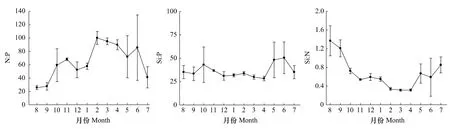
圖4 2010—2011年洞頭海域表層海水營養鹽比例月變化Fig.4 Monthly variation of nutrient ratios of surface waters in Dongtou Island during 2010—2011
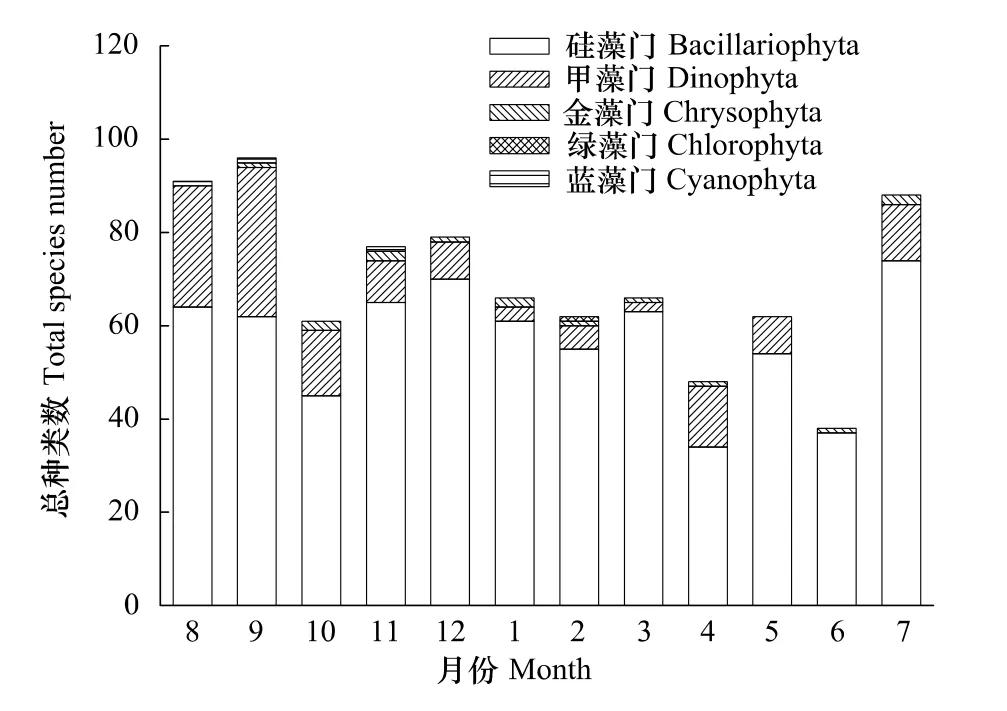
圖5 2010—2011年洞頭海域浮游植物種類組成月變化Fig.5 Monthly variation of phytoplankton species composition in Dongtou Island during 2010—2011
浮游植物種類在每相鄰兩月之間的相似性系數(JS)變幅為25.60—44.92%(表1),可以看出調查海域浮游植物月與月之間約50%以上的種類組成不相同,表明時間差異較明顯,浮游植物種類演替比較快。
2.2.2 生態類群
根據洞頭海域浮游植物的適溫范圍,可以將觀察到的浮游植物分為三類:(1)廣溫類群,該類群對溫度適應范圍較廣,是調查海域的優勢類群,代表性種類有尖刺偽菱形藻(Pseudo-nitzschia pungens)、翼鼻狀藻纖細變型(Proboscia alata f.gracillima)和尖葉原甲藻(Prorocentrum triestinum)等;(2)暖水類群,該類群適宜水溫較高的季節和水域,在調查海域夏、秋季出現較多,代表性種類有勞氏角毛藻(C.lorenzianus)、透明輻桿藻(Bacteriastrum hyalinum)和瓊氏圓篩藻(C.jonesianus)等;(3)暖溫類群,該類群在本海域出現頻率和豐度均較低,代表性種類有柔弱幾內亞藻(Guinardia delicatula)、短孢角毛藻(C.brevis)、扭鏈角毛藻(C.tortissimus)、筆尖形根管藻(Rhizosolenia styliformis)和皇冠角毛藻(C.diadema)等。

表1 2010—2011年洞頭海域浮游植物種類組成月相似性系數(JS,%)Table 1 Variation of species similarity index of phytoplankton in Dongtou Island during 2010—2011
2.2.3 細胞豐度
調查海域浮游植物細胞豐度時間變化較明顯(圖6),細胞豐度在7—9月較高,其它各月均維持在相對低的水平;從季節來看,夏季(128.83×106個/m3)>秋季(13.19×106個/m3)>冬季(10.81×106個/m3)>春季(3.98×106個/m3)。本海域硅藻細胞豐度全年變化趨勢與總細胞豐度趨勢一致,3月最低,為2.32×106個/m3,7—9月出現高值(圖6)。甲藻細胞豐度維持在較低水平,全年均值變化范圍為0—3.97×106個/m3,1月甲藻細胞豐度最高,此時尖葉原甲藻占優勢。金藻幾乎全年都出現,但細胞豐度較低,在104—105個/m3水平之間。藍藻全年細胞豐度也較低,在8月達最高值,而綠藻僅在個別月份出現。可以看出,硅藻作為浮游植物的主體,對浮游植物豐度的變化起支配作用。
2.2.4 優勢種
調查期間各月網采浮游植物優勢種不同(表2)。其中,1月優勢種最多,有7種,11月最少,僅1種,為骨條藻(Skeletonema spp.)。除6—8月外的其他各月,骨條藻均為主要優勢種。具槽帕拉藻(Paralia sulcata)在6月占主要優勢,優勢度(Y)為0.401。擬旋鏈角毛藻(C.pseudocurvisetus)在溫度較高的7和8月占一定優勢,且在7月Y高達0.659,細胞豐度為159.22×106個/m3。柔弱偽菱形藻(P.delicatissima)和尖刺偽菱形藻在8月占主要優勢,其Y分別為0.497和0.118。此外,優勢種多數為硅藻,甲藻較少,僅3種,分別為雙刺原多甲藻(P.bipes)、尖葉原甲藻和透鏡翼藻(Diplopsalis lenticula)。束毛藻(Trichodesmium spp.)在8和9月占一定優勢。

圖6 2010—2011洞頭海域浮游植物總豐度和各門類細胞豐度的月變化Fig.6 Monthly variation of total cells abundance and cell abundance of every phyla in Dongtou Island during 2010—2011

表2 2010—2011洞頭海域浮游植物優勢種組成及優勢度Table 2 Dominant phytoplankton species and their dominance in Dongtou Island during 2010—2011
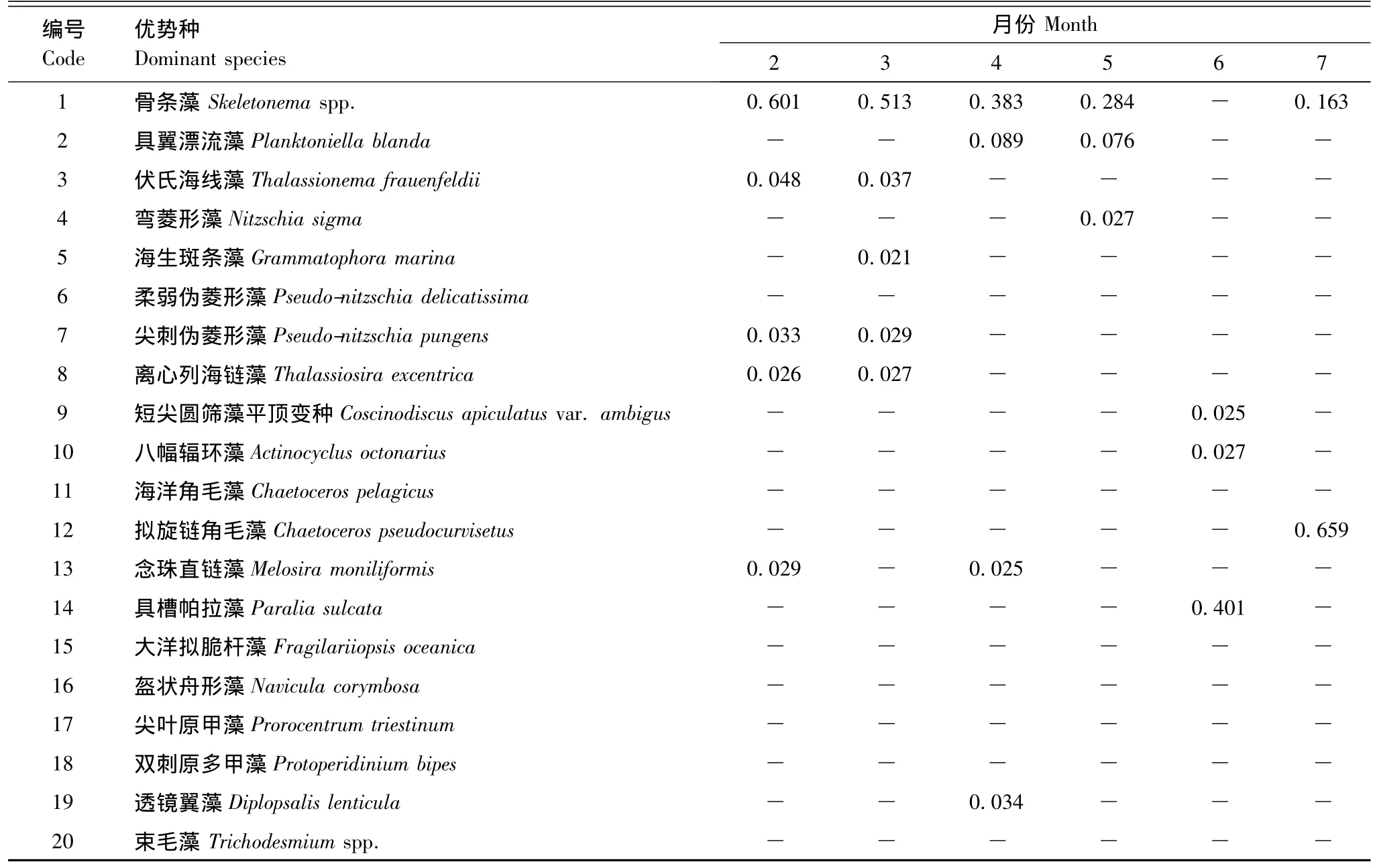
續表
2.2.5 群落多樣性指數
調查海域浮游植物群落多樣性指數年均值的變化如圖7所示。Shannon-Wiener指數(H′)和均勻度指數(J)周年變化趨勢大體一致。H′值9月最低,僅為0.73,4月最高,為2.10。J值9月最低,僅為0.22,4月最高,為0.74。從季節來看,H′值春季(2.08)>冬季(1.79)>夏季(1.73)>秋季(1.20),J值春季(0.70)>冬季(0.56)>夏季(0.52)>秋季(0.38)。
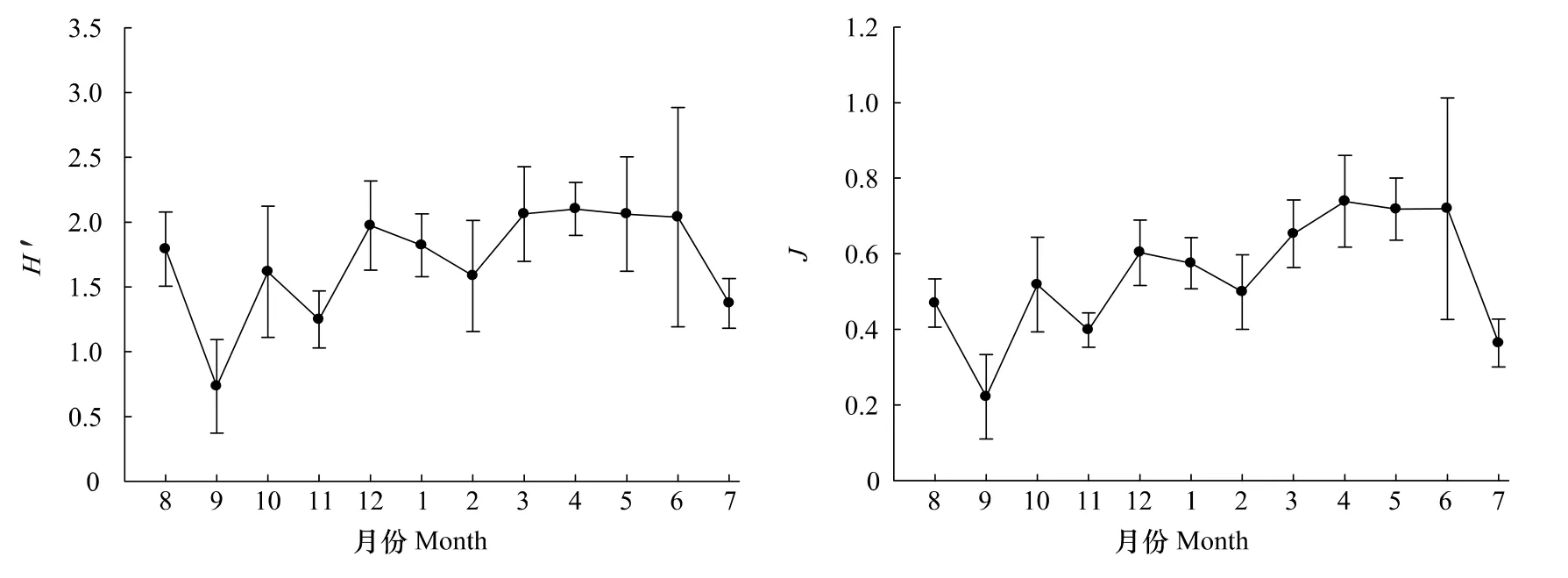
圖7 2010—2011洞頭海域浮游植物Shannon-Wiener指數(H′)和均勻度指數(J)月變化Fig.7 Monthly variation of H′and J of phytoplankton in Dongtou Island during 2010—2011
2.3 浮游植物群落與理化因子的關系
選擇各月優勢種(20種)用于CCA分析,浮游植物代碼見表2。在CCA排序圖中,箭頭表示環境因子,箭頭連線的長短表示物種分布與環境因子相關性的大小。蒙特卡羅檢驗(Monte Carlo test)表明,第1軸和全部軸P<0.05,故CCA分析的排序結果是可信的。CCA分析中的9個環境因子可解釋浮游植物群落總變量的90.9%。第1軸和第2軸的特征值分別為0.605和0.293,并各自解釋了35.7%和17.3%的物種變量。第1軸和第2軸的物種—環境相關系數分別為0.998和0.997,表明這9個環境因子與20種主要浮游植物種類相關性較好。營養鹽結構、鹽度和溫度是影響浮游植物群落的主要環境因子(圖8)。組Ⅰ (暖水高鹽種)與Si∶N、溫度和鹽度密切正相關,而與營養鹽濃度和N∶P呈負相關;組Ⅱ (近岸廣溫種)與組I剛好相反;組Ⅲ(附生種或底棲種)和具槽帕拉藻(14)對環境變化的耐受性較強,故與溫度、鹽度及營養鹽濃度和結構等基本無相關性;而骨條藻(1)屬相對廣溫廣鹽種,在該海域除夏季外均為優勢種,故位于坐標原點附近,受環境因子的影響較小。
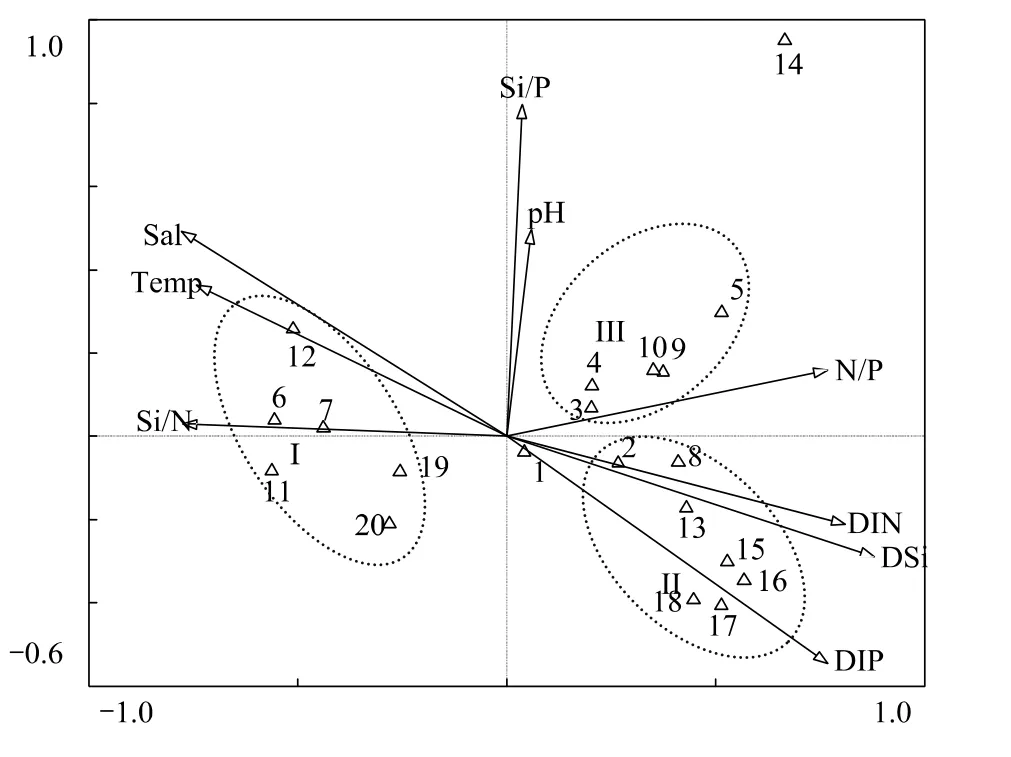
圖8 主要浮游植物種類與環境因子間的CCA排序圖Fig.8 CCA ordination of main phytoplankton species with environmental variables
3 討論
3.1 浮游植物群落與水團的關系
洞頭海域主要受臺灣暖流、閩浙沿岸流和甌江北口淡水徑流的影響。夏、秋季臺灣暖流影響較大[8-9],調查海域外海(鹽度29.8—30.1)暖水性種類增多,如海洋角管藻(Cerataulina pelagic)、遠距角毛藻(C.distans)和擬旋鏈角毛藻等硅藻類,及透鏡翼藻和馬西里亞角藻(C.massiliense)等甲藻類。其中,6月浮游植物種類數較低,這可能是由于采樣前發生過裸甲藻(Gymnodinium sp.)赤潮,導致海區種類數降低;9月浮游植物種類數高達97種,此時甲藻種類最多(32種)。冬、春季,受閩浙沿岸流與淡水徑流影響[8,10],調查海域既有相對低鹽(鹽度26.8—28.6)種類(瓊氏圓篩藻、布氏雙尾藻Ditylum brightwellii和尖葉原甲藻等),也有淡水種類和半咸水種類(二角盤星藻具刺變種Pediastrum duplex var.echinatum和針桿藻Synedra sp.等)。
浮游植物細胞豐度大部分月份處于較低水平,但在7和8月急劇增加,這分別與擬旋鏈角毛藻和偽菱形藻的大量生長有關。擬旋鏈角毛藻為暖水種類,有研究表明角毛藻適宜高溫條件下生長[22]。王艷等[23]研究也發現,擬旋鏈角毛藻在湛江海域的夏季大量繁殖,細胞豐度高達2289.87×106個/m3,是當地較重要的赤潮種。偽菱形藻也是重要的赤潮種類,且部分種類如P.multiseries可產生多莫酸(DA)毒素[24]。在本海區發現的偽菱形藻主要為尖刺偽菱形藻和柔弱偽菱形藻,該藻是典型的廣溫種,但在調查海區只在8月豐度較高,為128.16×106個/m3,此時海區溫度為29.9℃,鹽度為30.1,N∶P范圍為20.8—29.4。這與陳菊芳等[25]對大亞灣偽菱形藻的研究成果相似,他們認為大亞灣高密度的偽菱形藻出現的溫度、鹽度和N∶P范圍分別為25.0—30.0℃、28.9—31.3和6.2—32.9。另外,盔狀舟形藻(Navicula corymbosa)等附生性藻類在溫度較低的冬、春季豐度也較高,這主要由于本海域羊棲菜(Sargasum fusiforme)養殖的原因。
浮游植物多樣性可以表示群落的穩定性,當多樣性較高時,即優勢種種類較多,無優勢性特別突出的種類時,該群落結構是穩定的[26]。秋季H′值和J值均較低(圖7),分別為1.20和0.38,這是由于此時優勢種較單一,骨條藻占絕對優勢(表2);其他季節H′值和J值均較高。這表明本海域浮游植物群落結構尚屬穩定。
3.2 營養鹽對浮游植物的影響
CCA結果表明營養鹽也是影響本海域浮游植物群落的主要因素。Redfield[27]曾提出,浮游植物生長較合適的 Si∶N ∶P 值為16∶16∶1,當 N ∶P>16∶1 時,屬于磷限制,N ∶P<16∶1 時,屬于氮限制;當 Si∶N>1∶1 時,屬于氮限制,Si∶N<1∶1時,屬于硅限制。本海域全年N∶P均較高(>25∶1),因此浮游植物的生長受磷限制。一般而言,甲藻如原甲藻、鰭藻Dinophysis等對磷的需求較高[28],因此,磷限制了本海域甲藻的生長。從圖8也可以看出,甲藻優勢種豐度與磷酸鹽濃度密切正相關,1月磷酸鹽濃度較高(1.30 μmol/L),此時甲藻豐度較高,當磷酸鹽濃度降低時,甲藻豐度降低。柔弱偽菱形藻和尖刺偽菱形藻與Si∶N密切正相關,當Si∶N升高時,其豐度升高,而Hlaili等[28]認為偽菱形藻對氮的需求較高,氮濃度升高對偽菱形藻生長有利。這可能是因為偽菱形藻大量增殖消耗了水體中的氮,導致Si∶N升高,從而呈現出偽菱形藻豐度與Si∶N密切正相關的現象。擬旋鏈角毛藻在溫度較高而營養鹽濃度較低的夏季有較高的豐度,故它與溫度密切正相關,與磷酸鹽、硅酸鹽和硝酸鹽密切負相關。一些廣布種如具槽帕拉藻等生態幅較大,故其對環境因子的變化不敏感,受環境影響較小。
近30年來,洞頭海域浮游植物優勢種構成變化較明顯(表3)。圓篩藻在20世紀80年代初是較常見的優勢種,常在秋、冬季占主要優勢,主要種類為有棘圓篩藻(C.spinosus);進入90年代后,圓篩藻仍然為浮游植物群落的主要組成部分,但主要種類為瓊氏圓篩藻和虹彩圓篩藻(C.oculus-iridis),而且1990年春季其優勢地位被夜光藻(Noctiluca scientillans)取代;至今,本海域圓篩藻種類雖較多(16種),但豐度較低,已不再成為優勢種群。骨條藻在20世紀80和90年代細胞豐度較低,僅在個別季節占優勢,而在本研究調查期間,已成為本海域最重要的赤潮種。此外,甲藻優勢種也有較大變化,角藻在80年代占一定優勢,主要為梭角藻(C.fusus)和叉狀角藻(C.furca),隨后優勢度有所下降。本次調查期間,角藻種類雖也較多(10種),但不再占優勢,近幾年,本海域甲藻赤潮主要由東海原甲藻引起[12]。
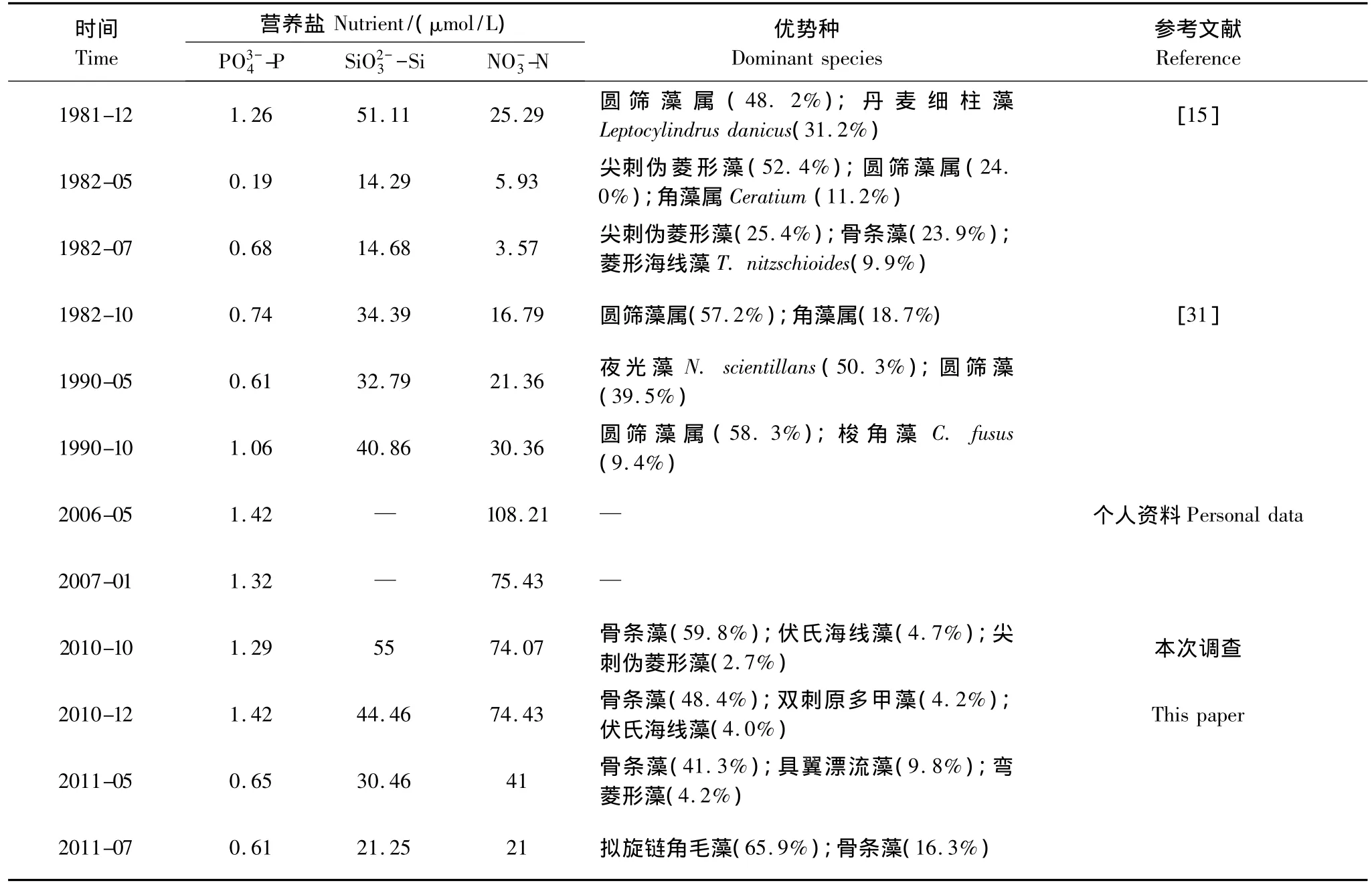
表3 不同調查年份洞頭海域營養鹽數據和網采浮游植物的群落參數Table 3 Nutrient concentration and community parameters of net-phytoplankton at different years in Dongtou Island
上述改變與洞頭海域環境因子的改變有關,如Zhou等[29]的研究認為,近50年來營養鹽結構(濃度及比例)的改變是長江口海域浮游植物豐度升高和優勢種組成改變的主要原因。從表3中可以看出,近30年來,磷酸鹽和硅酸鹽濃度變化不大,而硝酸鹽濃度急劇增加,80年代初期,其平均濃度僅為12.89 μmol/L,在本次調查中,其平均濃度高達52.63 μmol/L。同期,浮游植物優勢種也由圓篩藻向骨條藻等演替,Wang等[30]的研究也認為,營養鹽濃度升高會導致優勢種由大型硅藻(如根管藻等)向鏈狀硅藻(中肋骨條藻和偽菱形藻等)演替。另外,富營養化也是本海域東海原甲藻頻繁旺發的重要原因之一[12,29]。
致謝:感謝浙江省海洋水產養殖所謝起浪、柴雪良、李尚魯、林少珍、吳洪喜、王鐵桿、李磊和張立寧等的幫助。感謝冉莉華老師對英文摘要進行潤色。
[1] Zhou M J,Yan T,Zou J Z.Preliminary analysis of the characteristics of red tide areas in Changjiang River estuary and its adjacent sea.Chinese Journal of Applied Ecology,2003,14(7):1031-1038.
[2] Lassen M K,Nielsen K D,Richardson K,Garde K,Schlüter L.The effects of temperature increases on a temperate phytoplankton community——a mesocosm climate change scenario.Journal of Experimental Marine Biology and Ecology,2010,383(1):79-88.
[3] Alkawri A A S,Ramaiah N.Spatio-temporal variability of dinoflagellate assemblages in different salinity regimes in the west coast of India.Harmful Algae,2010,9(2):153-162.
[4] Sun J,Liu D Y,Yang S M,Guo J,Qian S B.The preliminary study on phytoplankton community structure in the central Bohai Sea and the Bohai strait and its adjacent area.Oceanologia et Limnologia Sinica,2002,33(3):461-471.
[5] Lie A A Y,Wong C K,Lam J Y C,Liu J H,Yung Y K.Changes in the nutrient ratios and phytoplankton community after declines in nutrient concentrations in a semi-enclosed bay in Hong Kong.Marine Environmental Research,2011,71(3):178-188.
[6] Granéli E,Weberg M,Salomon P S.Harmful algal blooms of allelopathic microalgal species:the role of eutrophication.Harmful Algae,2008,8(1):94-102.
[7] Han X F,Wang R.The grazing impact and regulation effects of zooplankton on phytoplankton bloom.Marine Sciences,2001,25(10):31-33.
[8] Su J L,Yuan L Y.Hydrology in China offshore.Beijing:Ocean Press,2005:207-246.
[9] Gao Q,Xu Z L.Species composition and diversity of zooplankton in Oujiang River estuary in summer and autumn.Chinese Journal of Ecology,2009,28(10):2048-2055.
[10] Zhang F.Seasonal variation features of chlorophyll a content in Taiwan Strait.Journal of Oceanography in Taiwan Strait,2001,20(3):314-318.
[11] Wang Y N,Zhang Y P,Wang X H.Ecologic investigation of mollusca on the intertidal zone of Dongtou island,Zhejiang.Journal of Zhejiang College of Fisheries,1994,13(3):179-182.
[12] Yao W M,Zheng A R,Qiu J K.Eutrophication in Dongtou islands,Zhejiang and the relationships between eutrophication and red tide.Marine Environmental Science,2007,26(5):466-469.
[13] Chen L,Xu Z L,Yao W M,Zhang Z,Chen S,Cai S W,Bao X P.The distribution of nutrients,phytoplankton and zooplankton in spring of the Oujiang River estuary.Acta Ecologica Sinica,2009,29(3):1571-1577.
[14] Li Y,Li H,Lü S H,Jiang T J,Xiao Y P,Chen S.Species diversity and distribution of phytoplankton in Nanji islands national nature reserve.Acta Hydrobiologica Sinica,2010,34(3):618-628.
[15] The Committee of Investigation of Symposium for Coastal and Intertidal Resources in Zhejiang Province.The report of coastal and intertidal resources of Zhejiang Province.Beijing:Ocean Press,1988.
[16] Standardization Administration of China.The Specification for Marine Monitoring(GB/T 17378.4—2007).Beijing:China Standards Press,2007.
[17] Standardization Administration of China.The Specification for Oceanographic Survey(GB/T 12763.6—2007).Beijing:China Standards Press,2007.
[18] Pielou,E C.An introduction to mathematical ecology.New York:Wiley-Interscience,1969.
[19] Magurran A E.Ecological diversity and its measurement.New Jersey:Princeton University Press,1988:1-179.
[20] Lep? J,?milauer P.Multivariate analysis of ecological data using CANOCO.London:Cambridge University Press,2003:1-280.
[21] Sun J,Liu D Y.The preliminary notion on nomenclature of common phytoplankton in China seas waters.Oceanologia et Limnologia Sinica,2002,33(3):271-286.
[22] Wang Z H,Qi Y Z,Chen J F,Xu N.Population dynamics of Chaetoceros spp.and multivariate analysis of its relationship with environmental factors in Daya Bay,South China Sea.Acta Ecologica Sinica,2006,26(4):1096-1102.
[23] Wang Y,Nie R,Li Y,Lü S H.Species diversity and geographical distribution of Chaetoceros in Guangdong coastal waters.Advances in Marine Science,2010,28(3):342-352.
[24] Hagstr?m J A,Granéli E,Moreira M O P,Odebrecht C.Domoic acid production and elemental composition of two Pseudo-nitzschia multiseries strains,from the NW and SW Atlantic Ocean,growing in phosphorus-or nitrogen-limited chemostat cultures.Journal of Plankton Research,2011,33(2):297-308.
[25] Chen J F,Xu N,Wang Z H,Huang W J,Xie L C,Jiang T J,Qi Y Z.Dynamics of Pseudo-nitzschia spp.and environmental factors in Daya Bay,the south China Sea.Acta Scientiae Circumstantiae,2002,22(6):743-748.
[26] Aktan Y.Large-scale patterns in summer surface water phytoplankton(except picophytoplankton)in the Eastern Mediterranean.Estuarine,Coastal and Shelf Science,2011,91(4):551-558.
[27] Redfield A C.The biological control of chemical factors in the environment.American Science,1958,46(3):205-221.
[28] Hlaili A S,Chikhaoui M A,Grami B E,Mabrouk H H.Effects of N and P supply on phytoplankton in Bizerte Lagoon(western Mediterranean).Journal of Experimental Marine Biology and Ecology,2006,333(1):79-96.
[29] Zhou M J,Shen Z L,Yu R C.Responses of a coastal phytoplankton community to increased nutrient input from the Changjiang(Yangtze)River.Continental Shelf Research,2008,28(12):1483-1489.
[30] Wang Z H,Zhao J G,Zhang Y J,Cao Y.Phytoplankton community structure and environmental parameters in aquaculture areas of Daya Bay,South China Sea.Journal of Environmental Sciences,2009,21(9):1268-1275.
[31] The Second Institute of Oceanography.The report of marine biological resources of islands in Zhejiang Province.Hangzhou,1993.
參考文獻:
[1] 周名江,顏天,鄒景忠.長江口鄰近海域赤潮發生區基本特征初探.應用生態學報,2003,14(7):1031-1038.
[4] 孫軍,劉冬艷,楊世民,郭健,錢樹本.渤海中部和渤海海峽及鄰近海域浮游植物群落結構的初步研究.海洋與湖沼,2002,33(5):461-471.
[7] 韓希福,王榮.海洋浮游動物對浮游植物水華的攝食與控制作用.海洋科學,2001,25(10):31-33.
[8] 蘇紀蘭,袁業立.中國近海水文.北京:海洋出版社,2005:207-246.
[9] 高倩,徐兆禮.甌江口夏、秋季浮游動物種類組成及其多樣性.生態學雜志,2009,28(10):2048-2055.
[10] 張釩.臺灣海峽葉綠素a含量的季節變化特征.臺灣海峽,2001,20(3):314-318.
[11] 王一農,張永普,王旭華.浙江洞頭島潮間帶軟體動物的生態調查.浙江水產學院學報,1994,13(3):179-182.
[12] 姚煒民,鄭愛榕,邱進坤.浙江洞頭列島海域水體富營養化及其與赤潮的關系.海洋環境科學,2007,26(5):466-469.
[13] 陳雷,徐兆禮,姚煒民,張釗,陳勝,蔡圣偉,鮑旭平.甌江口春季營養鹽、浮游植物和浮游動物的分布.生態學報,2009,29(3):1571-1577.
[14] 李揚,李歡,呂頌輝,江天久,蕭云樸,陳舜.南麂列島海洋自然保護區浮游植物的種類多樣性及其生態分布.水生生物學報,2010,34(3):618-628.
[15] 浙江省海岸帶和海涂資源綜合調查報告編寫委員會.浙江省海岸帶和海涂資源綜合調查報告.北京:海洋出版社,1988.
[16] 國家標準化管理委員會.海洋監測規范(GB/T17378.4—2007).北京:中國標準出版社,2007.
[17] 國家標準化管理委員會.海洋調查規范(GB/T 12763.6—2007).北京:中國標準出版社,2007.
[21] 孫軍,劉東艷.中國海區常見浮游植物種名更改初步意見.海洋與湖沼,2002,33(3):271-286.
[22] 王朝暉,齊雨藻,陳菊芳,徐寧.大亞灣角毛藻細胞數量波動及其與環境因子關系的多元分析.生態學報,2006,26(4):1096-1102.
[23] 王艷,聶銳,李揚,呂頌輝.廣東沿海角毛藻(Chaetoceros)的種類多樣性及其地理分布.海洋科學進展,2010,28(3):342-352.
[25] 陳菊芳,徐寧,王朝暉,黃偉健,謝隆初,江天久,齊雨藻.大亞灣擬菱形藻(Pseudo-nitzschia spp.)種群的季節變化與環境因子的關系.環境科學學報,2002,22(6):743-748.
[31] 國家海洋局第二海洋研究所.浙江省海島海域生物資源綜合調查報告.杭州,1993.
猜你喜歡
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
華人時刊(2020年13期)2020-09-25 08:21:30
趣味(語文)(2020年3期)2020-07-27 01:42:46
意林·全彩Color(2019年8期)2019-09-23 02:12:36
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
中國火炬(2014年11期)2014-07-25 10:31:58
中國體育(2004年3期)2004-11-11 08:53:02
棋藝(2001年19期)2001-11-25 19:55:34
棋藝(2001年23期)2001-01-06 19:08:36
