狗牙根與牛鞭草在三峽庫區(qū)消落帶水淹結(jié)束后的抗氧化酶活力
2013-12-16 08:19:42李兆佳熊高明鄧龍強(qiáng)謝宗強(qiáng)樊大勇
生態(tài)學(xué)報(bào) 2013年11期
李兆佳,熊高明,鄧龍強(qiáng),謝宗強(qiáng),樊大勇,*
(1.中國科學(xué)院植物研究所植被與環(huán)境變化國家重點(diǎn)實(shí)驗(yàn)室,北京 100093;2.中國科學(xué)院大學(xué),北京 100049;3.湖北神農(nóng)架國家級自然保護(hù)區(qū)管理局,湖北神農(nóng)架 442400)
三峽水庫運(yùn)行后,在庫區(qū)沿岸海拔145—175 m之間形成落差高達(dá)30 m的消落帶,原來的陸生生態(tài)系統(tǒng)演變?yōu)榧竟?jié)性濕地生態(tài)系統(tǒng)。生長環(huán)境的巨大變化和反季節(jié)水淹的頻繁干擾導(dǎo)致原有的大部分植物種類因不適應(yīng)而逐漸消亡[1]。三峽水庫消落帶將面臨生物多樣性減少,污染加劇,生態(tài)系統(tǒng)更為脆弱等問題[2]。因此,三峽水庫消落帶植被的生態(tài)治理對維持三峽庫區(qū)生態(tài)環(huán)境的穩(wěn)定具有十分重要的意義[3]。
對三峽水庫消落帶植被的生態(tài)治理首先需要研究的問題就是消落帶適宜植物的水淹耐受機(jī)理。對水淹脅迫耐受能力的強(qiáng)弱直接決定了植物是否適宜在消落帶環(huán)境生長。淹沒條件下,植物通過增加通氣組織、伸長生長、調(diào)節(jié)葉片形態(tài)結(jié)構(gòu)等方式增強(qiáng)水下氣體交換和水下光合[4]。克隆植物特有的克隆整合機(jī)制并有助于整個(gè)克隆片段抵御水淹脅迫[5]。此外,水淹前的碳水化合物儲存能力[6]、水淹過程中對碳水化合物的分配[7-8]以及水淹過后積極恢復(fù)的能力[9]被認(rèn)為是構(gòu)成植物水淹耐受能力的重要組分。
水淹結(jié)束后積極恢復(fù)的能力有利于消落帶的多年生植物占據(jù)空間生態(tài)位,在出露周期內(nèi)迅速恢復(fù)生長很可能使植株在水淹周期前儲存更多碳水化合物。多年野外調(diào)查結(jié)果發(fā)現(xiàn),三峽庫區(qū)消落帶形成后狗牙根和牛鞭草是適合當(dāng)?shù)厣车亩嗄晟荼局参铮?,10]。長期模擬水淹研究結(jié)果表明狗牙根和牛鞭草能夠在水淹結(jié)束后迅速恢復(fù)生長[11]。6個(gè)月水淹處理后狗牙根在15 d以內(nèi)開始恢復(fù)生長,且蓋度迅速增加[12]。Luo等[13]經(jīng)過30 d模擬水淹處理顯示,牛鞭草在水淹結(jié)束后10—20 d就開始恢復(fù)生長,且光合生理恢復(fù)領(lǐng)先于生長恢復(fù)。然而,目前是何種生理生態(tài)過程決定了恢復(fù)過程的快慢仍需要進(jìn)一步的研究。
長期水淹脅迫導(dǎo)致無氧呼吸替代有氧呼吸、細(xì)胞內(nèi)還原勢增加等一系列細(xì)胞生理狀態(tài)變化[14]。消落帶適宜物種要重新適應(yīng)水淹結(jié)束后正常的生長條件,其中一個(gè)問題是水淹過后植物與空氣的重新接觸可能會(huì)導(dǎo)致其受到的氧化脅迫程度明顯增高[15],即使植物可以忍耐長期的水淹脅迫,但在恢復(fù)期內(nèi)卻可能因?yàn)椴荒褪芨咚窖趸{迫而死亡[16],這可能是河岸帶和消落帶植物分布的重要限制因子[17]。大量研究表明植物體內(nèi)抵御氧化脅迫的體系非常完備,其中超氧化歧化酶(SOD,EC 1.15.1.1)、抗壞血酸過氧化物酶(APX,EC 1.11.1.11)、過氧化氫酶(CAT,EC1.11.1.6)是清除活性氧(ROS)、降低氧化脅迫的關(guān)鍵酶[18-19]。因此,水淹后的恢復(fù)階段,狗牙根和牛鞭草的恢復(fù)動(dòng)態(tài)是否會(huì)與其體內(nèi)清除ROS的SOD、APX、CAT的活性動(dòng)態(tài)相關(guān)?
目前,有關(guān)三峽消落帶適宜植物的水淹耐受機(jī)理的研究大部分采用模擬水淹方式[11,13,20]。由于模擬水淹實(shí)驗(yàn)不能完全對應(yīng)自然狀況(自然狀況下水壓、水深、光照等條件與模擬淹水實(shí)驗(yàn)條件差別很大),采用野外采樣、對比測定的手段,研究了三峽庫區(qū)消落帶生長的狗牙根和牛鞭草的根系在水淹后恢復(fù)過程中清除ROS的關(guān)鍵酶的活性動(dòng)態(tài),以期回答上述科學(xué)問題,并為消落帶人工植被恢復(fù)重建提供理論依據(jù)。
1 材料與方法
1.1 實(shí)驗(yàn)地點(diǎn)
實(shí)驗(yàn)地位于重慶市忠縣石寶鎮(zhèn)共和村三峽水庫消落帶(30°24′22″N,108°24′26″E)。屬暖濕亞熱帶山地氣候,溫?zé)岷疀觯募痉置鳎炅砍渑妫照粘渥恪?/p>
1.2 實(shí)驗(yàn)材料
狗牙根,Cynodon dactylon(L.)Pars,禾本科多年生草本植物,具根狀莖或匍匐莖,匍匐莖節(jié)上生根及分枝;對Cu、Cd、Sb等重金屬具有較強(qiáng)富集能力①陶星星,兩個(gè)典型礦區(qū)植物積累重金屬調(diào)查與研究(碩士論文).2008,湖南農(nóng)業(yè)大學(xué).,可作為牧草或草坪草;在國內(nèi)分布于華南、黃河流域及以南地區(qū)、長江流域及以南地區(qū)和新疆等地。
牛鞭草,Hemarthria altissima,禾本科多年生草本,有長而橫走的根莖。牛鞭草在我國分布較廣,東北、華北、華中、華南、西南各地皆有存在,多生于田地、水溝、河灘等濕潤處,常用作牧草。
兩者均具有克隆習(xí)性,能較快擴(kuò)張占領(lǐng)生境,在長江中上游地區(qū)牛鞭草常與狗牙根等相伴而生[21]。
1.3 實(shí)驗(yàn)設(shè)計(jì)
2008年3—6月劃定5 m×30 m樣地4塊,其中2塊位于海拔158 m,作為水淹及恢復(fù)處理組;另外2塊樣地位于海拔175 m,作為對照組。每個(gè)海拔的樣地按照20 cm×30 cm株行距分別移栽當(dāng)?shù)胤N源的狗牙根和牛鞭草,移栽后只作簡單的除雜草維護(hù)。
2009年春三峽水庫水位下降,158 m樣地出露前一天(第0天)進(jìn)行第1次取樣,此后每8d②本研究取樣間隔根據(jù)參考文獻(xiàn)以及實(shí)地觀察結(jié)果確定:檢測水淹(或缺氧)條件下抗氧化酶活力的取樣間隔根據(jù)處理時(shí)間長短從數(shù)十分鐘到數(shù)天不等;水淹處理長達(dá)數(shù)個(gè)月的文獻(xiàn)報(bào)道表明植株在水淹結(jié)束后3—10d內(nèi)開始恢復(fù)生長;據(jù)實(shí)地觀察,三峽庫區(qū)消落帶冬季水淹結(jié)束后,植株在1周內(nèi)基本開始復(fù)蘇,在1個(gè)月內(nèi)生長至正常水平取樣一次:分別在4塊樣地上各挖取植株,每塊樣地取4棵作為重復(fù),迅速洗凈根系,吸干水分后稱取0.5—1.0 g(鮮重)根部組織放入液氮中保存,用于檢測SOD、APX、CAT酶活力。為盡力保證植物樣品一致性,減少因日間光照、溫度、水分的日變化引起的生理狀態(tài)差異,每次取樣均在清晨5:00—8:00之間進(jìn)行。
根據(jù)長江水文網(wǎng)(http://www.cjh.com.cn)的三峽水庫忠縣水位記錄,實(shí)驗(yàn)期間海拔175 m樣地水淹共0 d,海拔158 m樣地水淹共195 d。
1.4 根系取樣與粗酶提取
取樣時(shí),把植株完整挖出,輕柔洗凈根上的土,并用蒸餾水沖洗3遍。稱取根系0.5—1 g,立即置于液氮中急凍,并于-70℃下保存,用于酶活力檢測。酶提取與活力檢測方法在Jiang和Zhang[22]基礎(chǔ)上作修改[23]:在預(yù)冷的研缽中放入剪碎的根系樣本,加入液氮,將樣本研磨至粉末,加入適量粗酶提取液,勻漿液4℃下以15000 g離心20 min,上清液為粗酶液。
1.5 SOD(EC 1.15.1.1)活力檢測
在盛有3 mL SOD反應(yīng)混合液(預(yù)熱至25℃)的試管中,加入100 μL粗酶液。終濃度分別為50 mmol/L磷酸鉀緩沖液(pH值7.8),13 mmol/L甲硫氨酸,75 μmol/L氮藍(lán)四唑(NBT),2 μmol/L 核黃素,0.1 mmol/L EDTA-Na2。混合后放在單排透明試管架上,在光照培養(yǎng)箱(25℃,12000lx)內(nèi)照光10 min,迅速測定OD560。以加粗酶液但不照光的反應(yīng)液為空白、蒸餾水代替粗酶液的照光管做對照。以抑制反應(yīng)50%的酶量為一個(gè)SOD酶活力單位。
1.6 APX(EC 1.11.1.11)活力檢測
2.5 mL APX反應(yīng)混合液(預(yù)熱至20℃)中依次加入100 μL粗酶液、0.4 mL H2O2稀釋液(預(yù)熱至20℃)。終濃度分別為50 mmol/L磷酸鉀緩沖液(pH值7.0),0.1 mmol/L EDTA-Na2,0.03 mmol/L AsA(抗壞血酸,ascorbate),0.06 mmol/L H2O2。加入H2O2后立即測定0—5 min內(nèi)的OD290值變化,每30 s記錄1次讀數(shù),計(jì)算單位時(shí)間內(nèi)AsA減少量(AsA比吸光系數(shù)2.8 mmol·L-1·cm-1)并計(jì)算酶活性(1個(gè)酶活定義為1U=1μmol AsA/s減少量)。蒸餾水作空白,對照用蒸餾水代替酶液。
1.7 CAT(EC 1.11.1.6)活力檢測
2.8 mL反應(yīng)液(預(yù)熱至20℃)中加入200 μL粗酶液,終濃度分別為50 mmol/L磷酸鉀緩沖液(pH值7.0),10 mmol/L H2O2。混合后立即測定0—2 min內(nèi)的OD240值變化,每30 s記錄1次讀數(shù),計(jì)算單位時(shí)間內(nèi)H2O2減少量(H2O2比吸光系數(shù)39.4 mmol·L-1·cm-1)及酶活性(1個(gè)酶活定義為1U=1 μmol H2O2/s減少量)。蒸餾水作空白,對照用蒸餾水代替酶液。
1.8 統(tǒng)計(jì)分析方法
數(shù)據(jù)的統(tǒng)計(jì)分析使用SPSS 16.0,采用t檢驗(yàn)分析海拔175 m與158 m之間對應(yīng)指標(biāo)的差異顯著性;用二元相關(guān)分析方法確定指標(biāo)之間的相關(guān)關(guān)系。用Origin 8.0作圖。
2 結(jié)果
長期水淹脅迫(圖1,海拔158 m,第0天)條件下,狗牙根和牛鞭草根部SOD、APX酶活力均高于對照(圖1,海拔175 m,第0天)且大部分呈顯著差異,與此相反,處理組CAT均顯著低于對照。
水淹結(jié)束后(圖1,第8天,第16天),SOD、APX酶活性均維持或上升到較高水平。狗牙根CAT稍低于對照而牛鞭草CAT上升幅度較大,但差異不顯著。
酶活力的變化反映了物種間的差異。牛鞭草SOD、APX、CAT酶活力相對值(處理/對照)均在8—16 d達(dá)最高,分別為對照的3.9倍、9.3倍、3.6倍,而狗牙根3個(gè)酶活力的變化逐漸趨向于對照值(圖1)。
盡管起始特征、恢復(fù)期變化動(dòng)態(tài)各有特點(diǎn),但恢復(fù)階段后期(第24天)兩個(gè)物種所有酶活力均與對照水平相近,沒有顯著差異。
相關(guān)分析表明,實(shí)驗(yàn)觀察過程中3個(gè)酶活力兩兩正相關(guān)。這種相關(guān)性在很大程度上與受脅迫然后逐漸恢復(fù)的植株有關(guān),因?yàn)槿舭凑蘸0蝿澐謹(jǐn)?shù)據(jù),相關(guān)顯著性完全集中于158 m處理組;不同物種之間則以牛鞭草根系各酶活力之間的相關(guān)性為主,狗牙根根系只有SOD-APX之間顯著相關(guān)(圖2)。
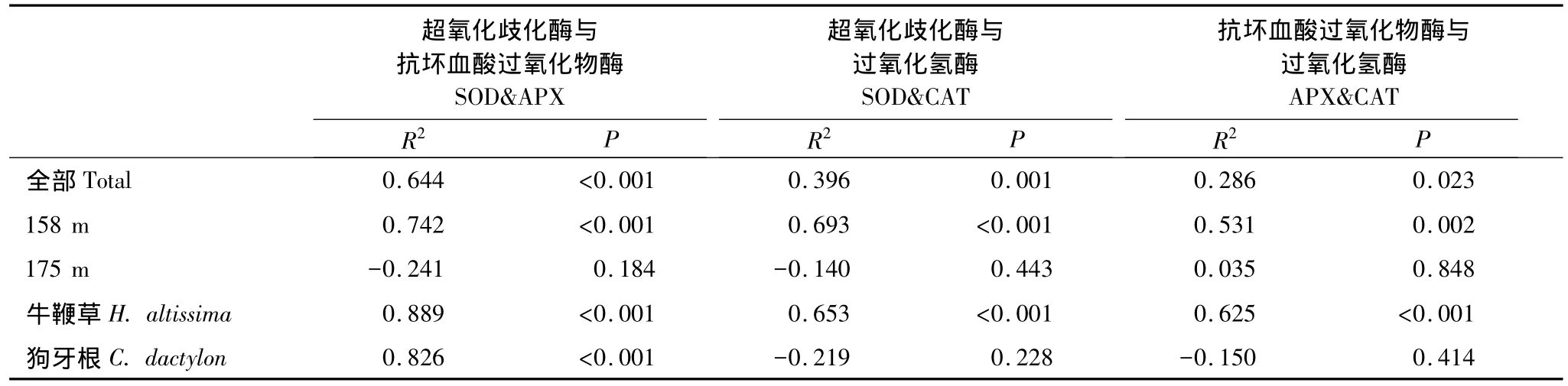
表1 基于檢測個(gè)體數(shù)據(jù)的SOD、APX、CAT酶活力Pearson相關(guān)系數(shù)及p值Table 1 Pearson correlation coefficient and p value of correlation analysis among antioxidant enzyme activities based on plant individuals
3 討論
3.1 長期水淹脅迫導(dǎo)致SOD、APX酶活力增強(qiáng)而CAT酶活力降低
長期水淹脅迫導(dǎo)致細(xì)胞內(nèi)ROS過度積累:細(xì)胞缺氧引起的高還原勢條件下NAD(P)H/NAD(P)+循環(huán)受阻、線粒體電子傳遞鏈飽和,電子在傳遞過程中泄漏并還原氧分子[14,24]。植物體可以通過提高ROS清除酶的活力以清除過度積累的ROS,降低氧化脅迫程度[25]。長期水淹脅迫后(圖1,第0天)狗牙根、牛鞭草的根部SOD、APX活力普遍顯著高于對照。這與一些研究報(bào)道相近:復(fù)氧階段羽扇豆(Lupinus luteus L.,cv.Juno)根系SOD、CAT活力上升,但POD只受到輕微影響[26];黃菖蒲(Iris pseudacorus L.)與德國鳶尾(Iris germanica L.)在缺氧處理和恢復(fù)階段的SOD酶活性均高于對照,且耐受性越高其增加幅度越大[27];缺氧增加了小麥根系SOD在非變性凝膠電泳中的條帶數(shù)量,導(dǎo)致SOD總活力上升并持續(xù)至復(fù)氧階段[28]。而狗牙根與牛鞭草的根部CAT起始活力均低于對照(圖1),這可能是受到對CAT活力的抑制作用影響。這種作用在體外實(shí)驗(yàn)中得到證實(shí),因而CAT在體內(nèi)條件下很可能也受影響[29]。
3.2 水淹結(jié)束后恢復(fù)期間清除ROS的關(guān)鍵酶活性發(fā)生了動(dòng)態(tài)變化
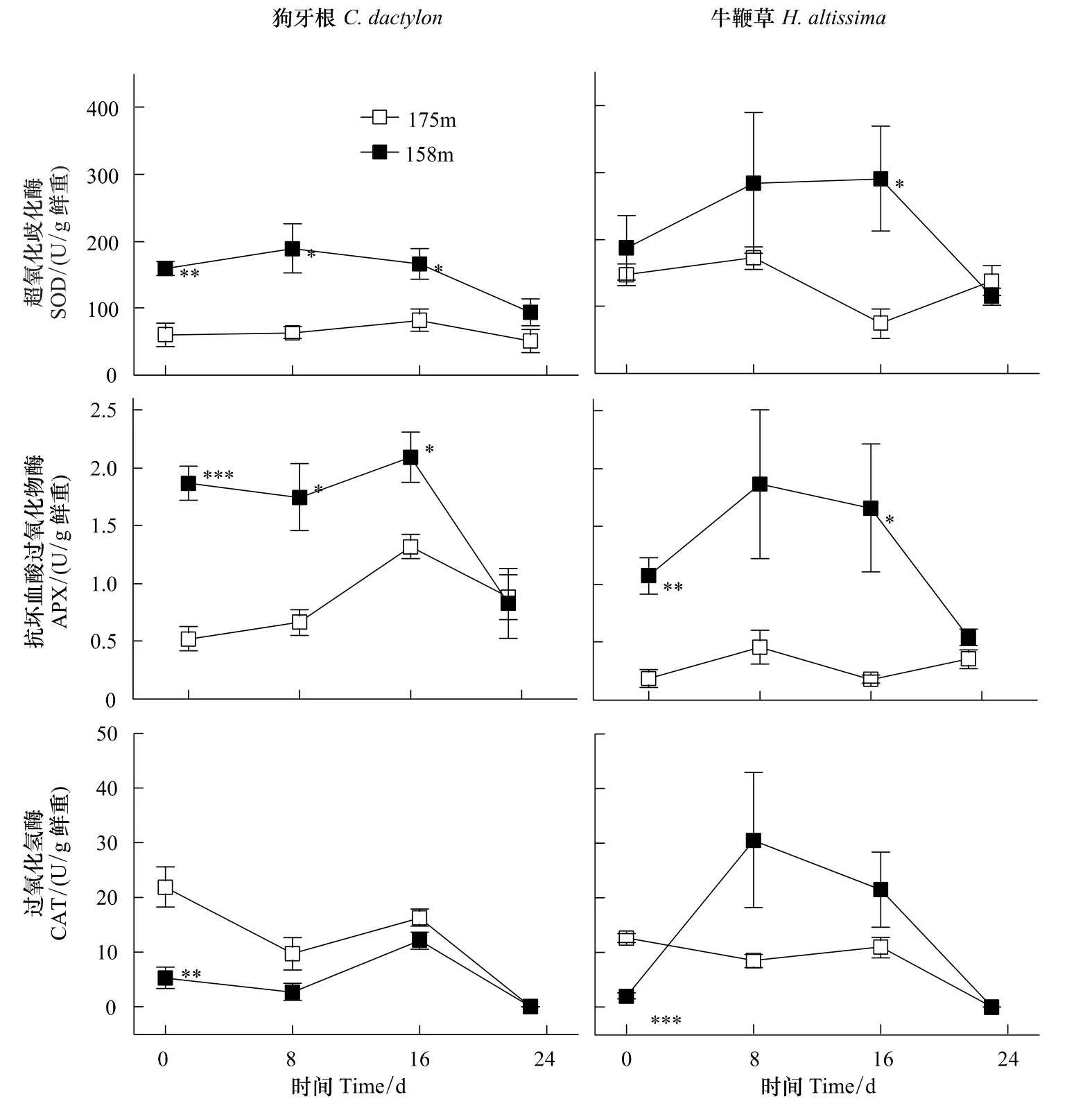
圖1 狗牙根與牛鞭草根系SOD、APX和CAT酶活力動(dòng)態(tài)Fig.1 Activity dynamics of SOD,APX and CAT from roots of C.dactylon and H.altissima
Unger等[30]對土壤化學(xué)特性的研究顯示,水淹結(jié)束后土壤孔隙的O2濃度與水淹期間相比存在顯著躍升。植物體與空氣重新接觸,水淹脅迫導(dǎo)致的處于過還原狀態(tài)的根際和細(xì)胞環(huán)境將O2還原為[31],這將會(huì)誘導(dǎo)或者加重(水淹脅迫引起的)氧化脅迫[32],并誘導(dǎo)清除ROS的關(guān)鍵酶的活性上升[33]。研究結(jié)果表明:相對于對照,這兩個(gè)物種在恢復(fù)階段的SOD和APX均保持了較高水平(圖1,第8天,第16天),說明恢復(fù)階段確實(shí)出現(xiàn)了較高水平的氧化脅迫。
水淹結(jié)束一段時(shí)間后,植物根部受脅迫誘導(dǎo)的無氧呼吸途徑關(guān)鍵酶活力下降[34]。可以認(rèn)為呼吸代謝逐漸恢復(fù)到正常的有氧代謝模式,O2重新作為電子最終受體,因而植物細(xì)胞內(nèi)ROS生產(chǎn)水平較恢復(fù)起始階段下降,ROS清除酶活力也可能因此隨之下降(圖1,第24天)。結(jié)果表明:狗牙根和牛鞭草根部的酶活力在24 d左右完全回復(fù)至對照水平,可以認(rèn)為細(xì)胞內(nèi)的ROS代謝已經(jīng)趨于穩(wěn)定平衡,植株已基本處于未受氧化脅迫的狀態(tài)。這個(gè)結(jié)果與長期模擬水淹實(shí)驗(yàn)的結(jié)果[12-13,20]吻合,說明恢復(fù)期內(nèi)氧化脅迫水平與恢復(fù)生長狀態(tài)緊密相關(guān)。
3.3 恢復(fù)過程中兩個(gè)物種酶活力動(dòng)態(tài)并不一致
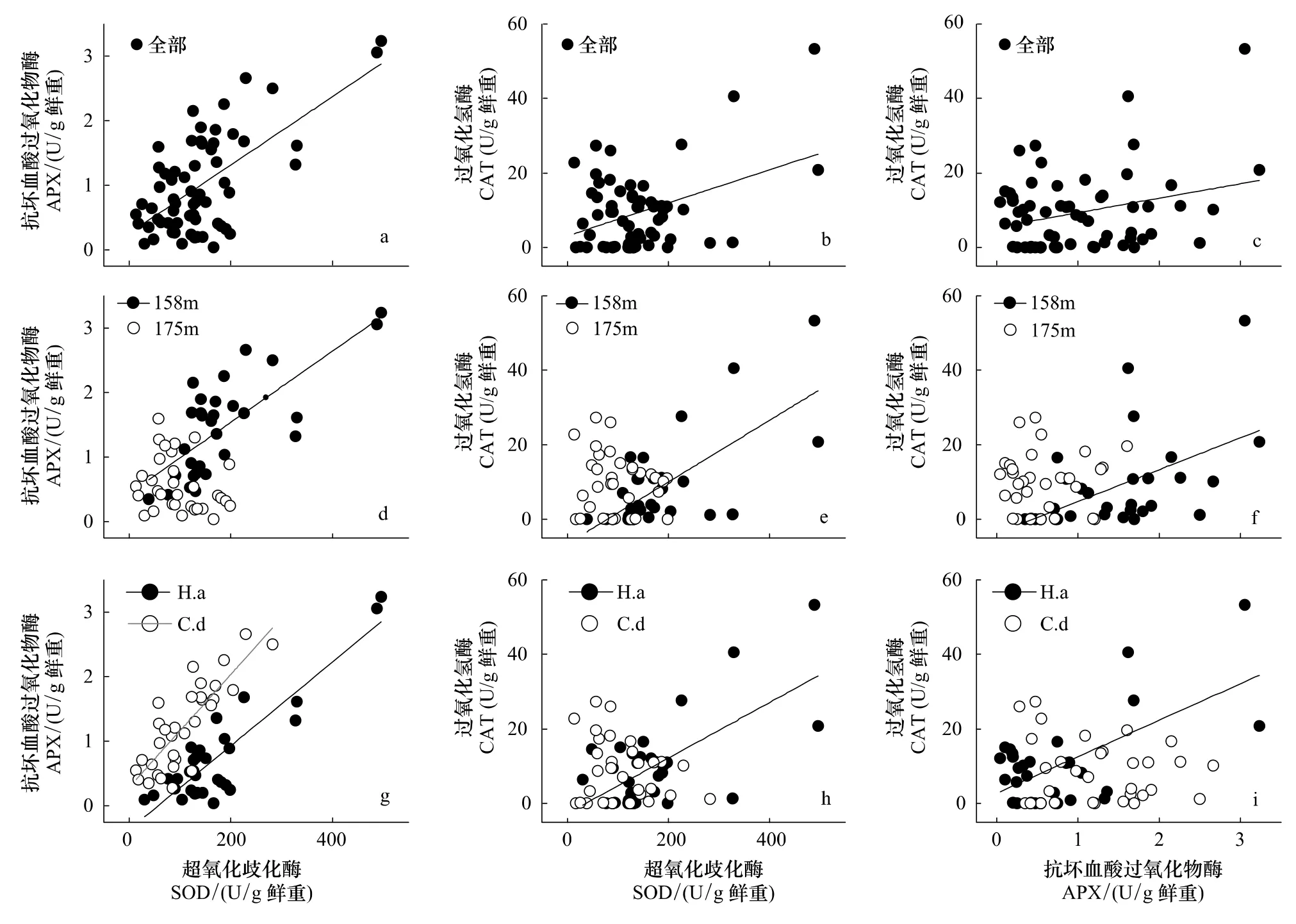
圖2 基于植株個(gè)體數(shù)據(jù)的SOD、APX和CAT酶活力相關(guān)分析Fig.2 Activity-correlation between SOD,APX and CAT based on plant individuals
本實(shí)驗(yàn)中,狗牙根CAT活力迅速恢復(fù)到對照水平,而牛鞭草CAT活力迅速增至較高水平,只因波動(dòng)較大而與對照未形成顯著差異(圖1)。這個(gè)差別可能源于兩個(gè)物種細(xì)胞中ROS調(diào)節(jié)途徑的不同響應(yīng)。在功能上,CAT、AsA-GSH循環(huán)(抗壞血酸-谷胱甘肽循環(huán))以及GPX(谷胱甘肽過氧化物酶)均能催化分解H2O2,在維持細(xì)胞氧化還原狀態(tài)穩(wěn)定方面,三者在一定程度上互為冗余、互相補(bǔ)充[35-37],因此,可能兩個(gè)物種在催化分解H2O2途徑上存在差異。此外,牛鞭草在恢復(fù)過程中3-個(gè)酶與對照組的相對活性均有不同程度的增加,而狗牙根3個(gè)酶的相對活性基本維持不變。這可能說明牛鞭草在恢復(fù)階段處于更高水平的氧化脅迫下;或其它機(jī)制也參與了對狗牙根的ROS清除過程。
3.4 消落帶位點(diǎn)恢復(fù)過程中3個(gè)酶的活性顯著正相關(guān)
海拔158 m位點(diǎn)的植物在恢復(fù)過程中抗氧化酶活力之間出現(xiàn)顯著相關(guān)(圖2),可能是植物細(xì)胞維持ROS代謝內(nèi)穩(wěn)態(tài)的體現(xiàn)。SOD分布于所有細(xì)胞器內(nèi),能夠催化超氧陰離子自由基()發(fā)生歧化反應(yīng),產(chǎn)生H2O2及O2[18],而APX和CAT則進(jìn)一步清除H2O2[19]。研究報(bào)道顯示,對細(xì)胞有潛在毒性的ROS同時(shí)也具有信號傳導(dǎo)作用[38-39],低濃度下,H2O2作為信號分子參與出發(fā)各種脅迫響應(yīng)[40],高濃度下則發(fā)出細(xì)胞程序性死亡[41]。因此,恢復(fù)期植物可以通過調(diào)節(jié)酶活力以及加強(qiáng)各關(guān)鍵酶的協(xié)作作用(相關(guān)關(guān)系)[32],最大能力清除過量積累的ROS,以避免細(xì)胞遭受不可逆轉(zhuǎn)的損傷。而對照組3個(gè)抗氧化酶活力之間沒有顯著相關(guān)性,原因可能是非氧化脅迫狀態(tài)下其他代謝途徑與ROS代謝的相互作用加強(qiáng),ROS代謝過程可能不是制約植物生存生長的唯一關(guān)鍵生理生態(tài)過程。
總之,我們采用野外采樣、對比測定的方法研究了三峽庫區(qū)消落帶生長的狗牙根和牛鞭草的根系在水淹后恢復(fù)過程中的ROS清除的關(guān)鍵酶的活性動(dòng)態(tài)。發(fā)現(xiàn):長期水淹脅迫導(dǎo)致SOD、APX酶活增強(qiáng)而CAT酶活降低;淹水后恢復(fù)期,清除ROS的關(guān)鍵酶活性發(fā)生了動(dòng)態(tài)變化,這可能與水淹過后的積極恢復(fù)生長能力有密切關(guān)系;恢復(fù)過程中兩個(gè)物種酶活動(dòng)態(tài)并不一致;恢復(fù)過程中3個(gè)酶的活性呈顯著相關(guān)。盡管有關(guān)ROS代謝與恢復(fù)生長之間的關(guān)系還需要進(jìn)一步的直接證據(jù),本文的研究結(jié)果有助于揭示消落帶植物適應(yīng)的生理生態(tài)機(jī)制,也為消落帶生態(tài)恢復(fù)治理實(shí)踐提供了重要的理論基礎(chǔ)。
致謝:感謝重慶大學(xué)郭勁松教授、夏玉仙教授和曾德玉老師提供檢測設(shè)備支持。
[1] Lu Z J,Li L F,Huang H D,Tao M,Zhang Q F,Jiang M X.Preliminary effects of impounding on vegetation in drawdown zone of the Three Gorges Reservoir region.Journal of Wuhan Botanical Research,2010,28(3):303-314.
[2] Tan S D,Wang Y,Zhang Q F.Environmental challenges and countermeasures of the water-level-fluctuation zone(WLFZ)of the Three Gorges Reservoir.Resources and Environment in the Yangtze Basin,2008,17(S1):101-105.
[3] Su W C.Main ecological and environmental problems of water-level-fluctuation zone(WLFZ)in Three Gorges reservoir and their controlling measures.Journal of Yangtze River Scientific Research Institute,2004,21(2):32-34.
[4] Voesenek L A C J,Colmer T D,Pierik R,Millenaar F F,Peeters A J M.How plants cope with complete submergence.New Phytologist,2006,170(2):213-226.
[5] Zhang X Y,F(xiàn)an D Y,Xie Z Q,Xiong G M,Li Z J.Clonal integration enhances performance of Cynodon dactylon subjected to submergence.Chinese Journal of Plant Ecology,2010,34(9):1075-1083.
[6] Sarkar R K.Saccharide content and growth parameters in relation with flooding tolerance in rice.Biologia Plantarum,1997,40(4):597-603.
[7] Angelov M N,Sung S J S,Doong R L,Harms W R,Kormanik P P,Black C C.Long-and short-term flooding effects on survival and sink-source relationships of swamp-adapted tree species.Tree Physiology,1996,16(5):477-484.
[8] Islam M A,Macdonald S E.Ecophysiological adaptations of black spruce(Picea mariana)and tamarack(Larix laricina)seedlings to flooding.Trees-Structure and Function,2004,18(1):35-42.
[9] Gibbs J,Greenway H.Mechanisms of anoxia tolerance in plants.I.Growth,survival and anaerobic catabolism.Functional Plant Biology,2003,30(1):1-47.
[10] Wang J C,Zhu B,Wang T.Characteristics of restoration of natural herbaceous vegetation of typical water-level fluctuation zone after flooding in the Three gorges Reservoir area.Resources and Environment in the Yangtze Basin,2011,20(5):603-610.
[11] Wang H F,Zeng B,Li Y,Qiao P,Ye X Q,Luo F L.Effects of long-term submergence on survival and recovery growth of four riparian plant species in Three Gorges Reservoir region,China.Journal of Plant Ecology,2008,32(5):977-984.
[12] Liu Y F,Liu Z X.Adaptability test of Cynodon dactylon(L.)Pers.in the ebb and flow zone of the Three Gorges Reservoir under extreme conditions.Journal of Southwest Agricultural University:Natural Science,2005,27(5):661-663.
[13] Luo F L,Nagel K A,Zeng B,Schurr U,Matsubara S.Photosynthetic acclimation is important for post-submergence recovery of photosynthesis and growth in two riparian species.Annals of Botany,2009,104(7):1435-1444.
[14] Blokhina O,Virolainen E,F(xiàn)agerstedt K V.Antioxidants,oxidative damage and oxygen deprivation stress:a review.Annals of Botany,2003,91(2):179-194.
[15] Crawford R M M,Braendle R.Oxygen deprivation stress in a changing environment.Journal of Experimental Botany,1996,47(2):145-159.
[16] Nabben R H M,Blom C W P M,Voesenek L A C J.Resistance to complete submergence in Rumex species with different life histories:the influence of plant size and light.New Phytologist,1999,144(2):313-321.
[17] Yi Y H,F(xiàn)an D Y,Xie Z Q,Chen F Q.The effects of waterlogging on photosynthesis-related eco-physiological processes in the seedlings of Quercus variabilis and Taxodium ascendens.Acta Ecologica Sinica,2008,28(12):6025-6033.
[18] Bowler C,Vanmontagu M,Inze D.Superoxide-dismutase and stress tolerance.Annual Review of Plant Physiology and Plant Molecular Biology,1992,43:83-116.
[19] May M,Vernoux T,Leaver C,Van Montagu M,Inze D.Glutathione homeostasis in plants:implications for environmental sensing and plant development.Journal of Experimental Botany,1998,49(321):649-667.
[20] Tan S D,Zhang S J,Zhang K R,Dang H S,Li M,Zhang Q F.Effect of long-time and deep submergence on recovery growth and photosynthesis of three grass species in Three Gorges Reservoir area.Journal of Wuhan Botanical Research,2009,27(4):391-396.
[21] Yang C H,Zhang X Q,Li X L,Du Y,Wu Y Q.Hemarthria germplasm resources and breeding.Acta Prataculturae Sinica,2004,13(2):7-12.
[22] Jiang M,Zhang J.Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves.Journal of Experimental Botany,2002,53(379):2401-2410.
[23] Li Z J,Yu J,F(xiàn)an D Y,Xie Z Q,Xiong G M,Zhang X Y.Clonal integration enhances the ability to scavenge reactive oxygen species in root of Cynodon dactylon subjected to submergence.Acta Ecologica Sinica,2011,31(17):4992-4999.
[24] Mittler R.Oxidative stress,antioxidants and stress tolerance.Trends in Plant Science,2002,7(9):405-410.
[25] Monk L S,F(xiàn)agerstedt K V,Crawford R M M.Oxygen-toxicity and superoxide-dismutase as an antioxidant in physiological stress.Physiologia Plantarum,1989,76(3):456-459.
[26] Garnczarska M,Bednarski W.Effect of a short-term hypoxic treatment followed by re-aeration on free radicals level and antioxidative enzymes in lupine roots.Plant Physiology and Biochemistry,2004,42(3):233-240.
[27] Monk L S,F(xiàn)agerstedt K V,Crawford R M M.Superoxide dismutase as an anaerobic polypeptide:a key factor in recovery from oxygen deprivation in iris pseudacorus?Plant Physiology,1987,85(4):1016-1020.
[28] Biemelt S,Keetman U,Mock H P,Grimm B.Expression and activity of isoenzymes of superoxide dismutase in wheat roots in response to hypoxia and anoxia.Plant,Cell and Environment,2000,23(2):135-144.
[29] Kono Y,F(xiàn)ridovich I.Superoxide radical inhibits catalase.Journal of Biological Chemistry,1982,257(10):5751-5754.
[30] Unger I M,Motavalli P P,Muzika R M.Changes in soil chemical properties with flooding:a field laboratory approach.Agriculture,Ecosystems and Environment,2009,131(1/2):105-110.
[31] Sairam R K,Kumutha D,Ezhilmathi K,Deshmukh P,Srivastava G.Physiology and biochemistry of waterlogging tolerance in plants.Biologia Plantarum,2008,52(3):401-412.
[32] Hossain Z,López-Climent M F,Arbona V,Pérez-Clemente R M,Gómez-Cadenas A.Modulation of the antioxidant system in citrus under waterlogging and subsequent drainage.Journal of Plant Physiology,2009,166(13):1391-1404.
[33] Veselova T V,Veselovsky V A,Turovetsky V B,Galchuk S V,Vanyushin V F,Aleksandrushkina N I,Rubin A B.Post-hypoxic oxidative stress after radicle protrusion as a possible cause for the production of abnormal seedlings in pea.Seed Science and Technology,2004,32(2):283-296.
[34] Dubey H,Grover A.Respiratory pathway enzymes are differentially altered in flood tolerant and sensitive rice types during O2deprivation stress and post-stress recovery phase.Plant Science,2003,164(5):815-821.
[35] Willekens H,Chamnongpol S,Davey M,Schraudner M,Langebartels C,Van Montagu M,Inze D,Van Camp W.Catalase is a sink for H2O2and is indispensable for stress defence in C3plants.The EMBO Journal,1997,16(16):4806-4816.
[36] Vanderauwera S,Zimmermann P,Rombauts S,Vandenabeele S,Langebartels C,Gruissem W,Inzé D,Van Breusegem F.Genome-wide analysis of hydrogen peroxide-regulated gene expression in Arabidopsis reveals a high light-induced transcriptional cluster involved in anthocyanin biosynthesis.Plant Physiology,2005,139(2):806-821.
[37] Mhamdi A,Hager J,Chaouch S,Queval G,Han Y,Taconnat L,Saindrenan P,Gouia H,Issakidis-Bourguet E,Renou J P,Noctor G.Arabidopsis glutathione reductase1 plays a crucial role in leaf responses to intracellular hydrogen peroxide and in ensuring appropriate gene expression through both salicylic acid and jasmonic acid signaling pathways.Plant Physiology,2010,153(3):1144-1160.
[38] Jabs T,Dietrich R A,Dangl J L.Initiation of runaway cell death in an Arabidopsis mutant by extracellular superoxide.Science,1996,273(5283):1853-1856.
[39] Foreman J,Demidchik V,Bothwell J H F,Mylona P,Miedema H,Torres M A,Linstead P,Costa S,Brownlee C,Jones J D G,Davies J M,Dolan L.Reactive oxygen species produced by NADPH oxidase regulate plant cell growth.Nature,2003,422(6930):442-446.
[40] Mittler R,Vanderauwera S,Gollery M,Van Breusegem F.Reactive oxygen gene network of plants.Trends in Plant Science,2004,9(10):490-498.
[41] Dat J,Vandenabeele S,Vranová E,Van Montagu M,Inzé D,Van Breusegem F.Dual action of the active oxygen species during plant stress responses.Cellular and Molecular Life Sciences,2000,57(5):779-795.
參考文獻(xiàn):
[1] 盧志軍,李連發(fā),黃漢東,陶敏,張全發(fā),江明喜.三峽水庫蓄水對消漲帶植被的初步影響.武漢植物學(xué)研究,2010,28(3):303-314.
[2] 譚淑端,王勇,張全發(fā).三峽水庫消落帶生態(tài)環(huán)境問題及綜合防治.長江流域資源與環(huán)境,2008,17(S1):101-105.
[3] 蘇維詞.三峽庫區(qū)消落帶的生態(tài)環(huán)境問題及其調(diào)控.長江學(xué)院院報(bào),2004,21(2):32-34.
[5] 張想英,樊大勇,謝宗強(qiáng),熊高明,李兆佳.克隆整合有助于狗牙根抵御水淹.植物生態(tài)學(xué)報(bào),2010,34(9):1075-1083.
[10] 王建超,朱波,汪濤.三峽庫區(qū)典型消落帶淹水后草本植被的自然恢復(fù)特征.長江流域資源與環(huán)境,2011,20(5):603-610.
[11] 王海鋒,曾波,李婭,喬普,葉小齊,羅芳麗.長期完全水淹對4種三峽庫區(qū)岸生植物存活及恢復(fù)生長的影響.植物生態(tài)學(xué)報(bào),2008,32(5):977-984.
[12] 劉云峰,劉正學(xué).三峽水庫消落區(qū)極限條件下狗牙根適生性試驗(yàn).西南農(nóng)業(yè)大學(xué)學(xué)報(bào):自然科學(xué)版,2005,27(5):661-663.
[17] 衣英華,樊大勇,謝宗強(qiáng),陳芳清.模擬淹水對池杉和栓皮櫟光合生理生態(tài)過程的影響.生態(tài)學(xué)報(bào),2008,28(12):6025-6033.
[20] 譚淑端,張守君,張克榮,黨海山,黎明,張全發(fā).長期深淹對三峽庫區(qū)三種草本植物的恢復(fù)生長及光合特性的影響.武漢植物學(xué)研究,2009,27(4):391-396.
[21] 楊春華,張新全,李向林,杜逸,吳彥奇.牛鞭草屬種質(zhì)資源及育種研究.草業(yè)學(xué)報(bào),2004,13(2):7-12.
[23] 李兆佳,喻杰,樊大勇,謝宗強(qiáng),熊高明,張想英.克隆整合提高淹水脅迫下狗牙根根部的活性氧清除能力.生態(tài)學(xué)報(bào),2011,31(17):4992-4999.
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
