青藏高原高寒草原區工程跡地面積對其恢復植物群落特征的影響
2013-12-16 08:19:48郭正剛
生態學報 2013年11期
毛 亮,周 杰,郭正剛
(蘭州大學草地農業科技學院草地農業生態系統國家重點實驗室,蘭州 730020)
高寒草原是青藏高原草地的重要組分[1],也是我國重要的高山植物基因庫和碳匯載體[2],更是流域生態安全屏障和當地畜牧業生產的物質基礎[3]。青藏公路和鐵路,高壓電線和光纜等人類線性工程穿越青藏高原高寒高原時,總會在施工地段或附近地區形成面積大小不同的工程跡地[4],因此工程跡地植被恢復是目前青藏高原受損高寒草原生態系統修復的重要內容。西方國家工程跡地恢復最早始于20世紀40年代的筑路跡地恢復[5],而我國筑路跡地恢復的研究始于高速公路護坡[6],21世紀初期開始逐漸增多。筑路跡地恢復包括人工恢復和自然恢復兩種。由于我國筑路跡地恢復始于高速公路建設,因此筑路跡地恢復目前以人工恢復研究較多[4,6-7]。雖然筑路跡地自然恢復是植被恢復的一種主要方式,且具有省時、省力和省財,避免草皮移植對客地原生草原影響等優點,但目前研究相對較少。馬世震等[5]通過對比不同恢復時期植物群落及多樣性特征,指出青藏高原高寒草原區工程跡地植被恢復至少需要20年;祝廣華等[6]通過比較不同草地類型經歷等同恢復時間的特征,認為高寒草原和高寒草甸是受工程活動影響最廣的草地類型;Guo等[3]通過分析未干擾群落與離工程跡地不同距離處恢復群落的特征,明晰筑路工程對高寒草原的干擾尺度在離路基100 m的范圍內。然而目前關于工程跡地自然恢復研究均集中于工程跡地上已經恢復的植物群落,忽略經歷相同時期因面積較大至今仍然沒有植被恢復的工程跡地,這說明工程跡地面積影響著工程跡地植物群落的自然恢復進程,但工程跡地面積變化與恢復植物群落的關系多限于直觀的感受,缺乏科學試驗提供佐證。
人類工程跡地主要包括路基周邊,取棄土場,沙石料場,施工便道,營地場地及施工地等[7],其不同的受損程度決定了植被恢復進程存在著差異。青藏公路路基主體顯著高于周邊取土跡地,路基周邊土壤因匯集了路面降水而持有較高的含水量,而且容易滯留風媒種子和花粉[8],從斑塊恢復學上,路基周邊跡地擁有最好的植物群落自然恢復物質基礎,從干擾程度上,路基周邊植被破壞嚴重(筑路一般就近取土),分布范圍廣,具有一定的代表性和典型性[3,7]。本研究以青藏高原平坦地區青藏公路的路基周邊跡地為例,通過調查經歷相同恢復時期而面積不同工程跡地上的植物群落特征,明確工程跡地面積對青藏高原高寒草原區植被恢復進程的影響。
1 材料與方法
1.1 研究區概況
研究地區位于青藏公路沿線沱沱河北岸,距沱沱河20 km,(即國道109線3144號路基樁和3126號路基樁之間),地理坐標為92°37′E,34°22′N,海拔4550—4560 m。氣候屬高原大陸性氣候,全年冰凍期 331 d,無霜期9—50 d,平均氣溫-4.2℃,最低氣溫-14.8℃,最高氣溫6.7℃,年均降水量275.92 mm,夏季降水占全年降雨70%[9]。土壤以高寒草原土為主。未受工程干擾的草原為青藏苔草(Carex moorcroftii)紫花針茅(Stipa purpurea)草原,優勢種為紫花針茅和青藏苔草,主要伴生種有草地早熟禾(Poa pratensis)、短穗兔耳草(Lagotis brachystachya)和矮火絨草(Leontopodium nanum)等[10]。
1.2 研究方法
1.2.1 樣地設置與調查
在1994年擴建和修繕青藏公路時遺留的路基周邊跡地上布設樣地。為了選擇研究樣地,首先實測56個單獨連片的工程跡地,記錄各個工程跡地恢復植物群落的高度、蓋度,觀察紫花針茅是否出現。筑路跡地形狀多為狹長型,中間為長方形,兩邊為半圓形,在測定其面積時,采用分割法測定求和。然后以恢復植物群落的高度、蓋度、紫花針茅出現否作為變量,對56個工程跡地聚類,聚類結果表明,56個工程跡地可劃分為4個類群,4個類群工程跡地面積的平均值分別為55,156,254,583 m2。根據4個類群工程跡地面積平均值設置樣地,工程跡地面積梯度分別為(55±10)m2,(156±20)m2,(254±30)m2,(583±40)m2,形成4 個面積梯度處理,然后每個梯度選擇3個樣地作為研究對象(3次重復),共計12個,12個樣地的周長和面積基本接近(表1)。同時在工程跡地鄰近、未受工程干擾的青藏苔草紫花針茅草原布設3個10 m×10 m的對照樣地。共計15個樣地,各個樣地的海拔、坡向、地表基況基本保持一致。在每個樣地對角線上布設4個1 m×1 m的樣方,分別統計樣方內的物種組成、群落蓋度、高度、頻度,然后收獲期地上生物量,帶回實驗室烘干稱重,共計調查樣方60個。野外調查于2011年7—8月進行。
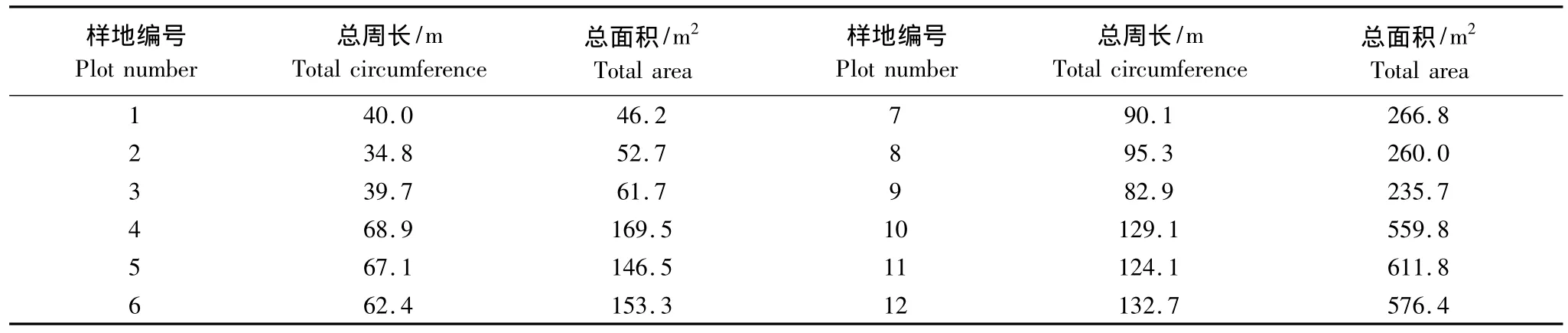
表1 12個筑路工程跡地樣地的周邊和面積Table 1 Total circumferences and areas of 12 plots from land used for engineering construction
1.2.2 α多樣性的測度
分別選用Shannon-Wiener指數、Pielou指數、Simpson指數、樣方內出現的物種數,測度多樣性指數(H)、均勻度指數(J)、優勢度指數(D)和豐富度指數(S)[11],其計算公式分別為:
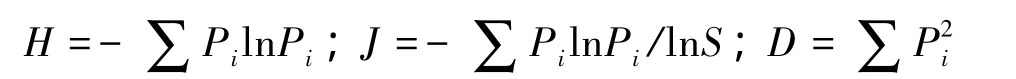
式中,Pi為種i的重要值,即Pi=(相對蓋度+相對密度+相對頻度)/3,S以樣方內出現的物種數量度量。
β多樣性選用Whittaker指數(βws)測度[3],其計算公式分別為:

式中,S為系統的總物種數;ma為樣方的平均物種數。
1.2.3 植被恢復能力評價
評價工程跡地與自然地植被恢復能力的綜合指標用SL[12-13],計算公式為:
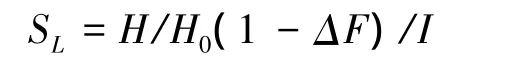
式中,H代表工程跡地Shannon-Wiener指數;H0代表未干擾地Shannon-Wiener指數;ΔF指相同地點工程跡地與未干擾草地植被覆蓋度差值;I是頂級物種侵入指數,計算方法為:
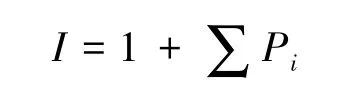
式中,Pi表示原優勢建群物種出現的頻度(即優勢建群物種的重要值)。
1.3 數據處理
采用SPSS 17.0軟件進行ANOVA方差分析,Excel制圖。
2 結果
2.1 工程跡地面積對恢復植物群落組分的影響
工程跡地上恢復植物群落的優勢種和主要伴生種隨著工程跡地面積變化而發生明顯變化(表2)。工程跡地面積為583 m2時,群落優勢種為短穗兔耳草,雖然青藏苔草和草地早熟禾等草原植物成功定居,但沒有成為群落優勢種群。工程跡地面積為254 m2時,優勢種被青藏苔草替代,而短兔耳草淪落為伴生種,此時定居的伴生種還有矮火絨草和草地早熟禾等。工程跡地面積縮至156 m2時,群落優勢種演變為青藏苔草和草地早熟禾,伴生種出現沙生鳳毛菊(Saussurea arenaria)、羽葉點地梅(Pomatosace filicula)和紫花針茅。工程跡地面積縮小到55 m2,優勢種除了青藏苔草以外,還有矮火絨草,此時高寒草原指示植物紫花針茅作為首要伴生種出現。未受筑路工程干擾的高寒草原主要以紫花針茅和青藏苔草為優勢種,伴生種種類豐富。這說明隨著工程跡地面積的增加,植物群落恢復速率越慢,環境旱生化,恢復植物群落組分結構越單一,草原指示物種出現越困難,而工程跡地面積越小,恢復植物群落越接近頂級群落,在工程跡地156 m2內出現了高寒草原指示物種紫花針茅,然后隨著工程跡地面積縮小,群落結構趨向復雜。

表2 青藏高原高寒草原區不同工程跡地面積上恢復植物群落的組分Table 2 Component of restorable plant community at different land areas used for engineering construction in the alpine steppe of the Qinghai-Tibetan Plateau
2.2 工程跡地面積對恢復植物群落空間結構的影響
2.2.1 群落高度
青藏高原高寒草原區,恢復植物群落的高度隨著工程跡地面積減小而逐漸增加(圖1)。當工程跡地面積小于或等于156 m2時,雖然恢復群落間或者恢復群落與未干擾群落間的高度有所差異,但并不顯著,但其顯著大于工程跡地面積為583 m2時恢復植物群落的高度(P<0.05)。工程跡地面積為55、156、254、583 m2時恢復植物群落的高度分別為未干擾群落高度的65%、69%、73%和83%,反映了工程跡地面積越小,恢復植物群落高度約接近于未干擾群落的高度。
2.2.2 群落總蓋度
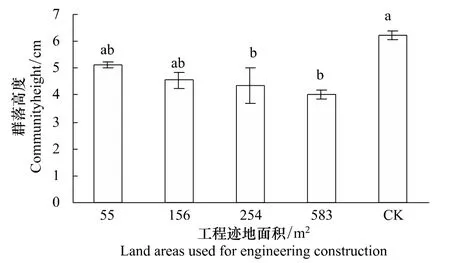
圖1 青藏高原高寒草原區不同工程跡地面積上恢復植物群落高度Fig.1 Height of restorable plant community at different land areas used for engineering construction in the alpine steppe of the Qinghai-Tibetan Plateau
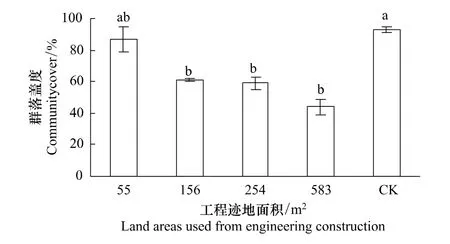
圖2 青藏高原高寒草原區不同面積工程跡地上恢復群落總蓋度Fig.2 Covers of restorable plant community at different land areas used for engineering construction in the alpine steppe of the Qinghai-Tibetan Plateau
隨著工程跡地面積增加,恢復群落總蓋度逐漸減少(圖2),當工程跡地面積為55 m2,恢復群落蓋度和未干擾群落蓋度沒有顯著差異,而他們顯著大于工程跡地面積大于或等于156 m2時恢復植物群落的總蓋度(P<0.05),工程跡地面積為55、156、254、583 m2時恢復植物群落的總蓋度分別為未干擾群落蓋度的47%、62%、65%和87%,說明工程跡地面積越小,植物群落蓋度越易恢復,反之亦然。
2.3 工程跡地面積對植物多樣性的影響
2.3.1 α多樣性
隨著工程跡地面積減小,植物物種多樣性指數和豐富度指數均逐漸增大(表3),其中多樣性指數在工程跡地面積55 m2時與未干擾草地沒有顯著差異,但當工程跡地面積大于156 m2,其恢復群落的多樣性指數顯著小于未干擾草地的多樣性指數(P<0.05),說明工程跡地面積為156 m2,工程跡地恢復群落的物種多樣性指數已接近于未干擾地群落的多樣性指數。植物物種豐富指數和多樣性指數隨著工程跡地面積變化過程基本趨同。隨著工程跡地面積減小,植物均勻度指數變化較為復雜,其中均勻度指數的最大值和最小值分別出現在工程跡地面積最大的樣地和未干擾地,在恢復植物群落序列中,均勻度指數在程跡地面積為254 m2時最小,此時均勻度指數和未干擾群落均勻度指數差異不顯著。優勢度指數隨著工程跡地面積變化雖然存在分異,但其在恢復群落之間,以及恢復群落和未干擾群落間均差異不顯著,說明工程跡地面積變化對優勢度指數沒有顯著的影響。

表3 青藏高原高寒草原區不同面積工程跡地上恢復植物群落的豐富度指數、均勻度指數、多樣性指數和優勢度指數Table 3 Richness index,evenness index,diversity index and dominant index of restorable plant community at different land areas used for engineering construction in the alpine steppe of the Qinghai-Tibetan Plateau
2.3.2 β多樣性
植物群落β多樣性指數隨工程跡地面積減小而逐漸減小(圖3),工程跡地面積為583 m2時群落的β多樣性指數顯著大于工程跡地面積156 m2和254 m2的β多樣性指數(P<0.05),而后者顯著大于工程跡地面積為55 m2時的恢復群落的β多樣性指數(P<0.05),所有恢復群落的β多樣性指數顯著大于未干擾群落的β多樣性指數(P<0.05),說明工程跡地面積越小,環境異質性越小,群落內物種替代速率降低。
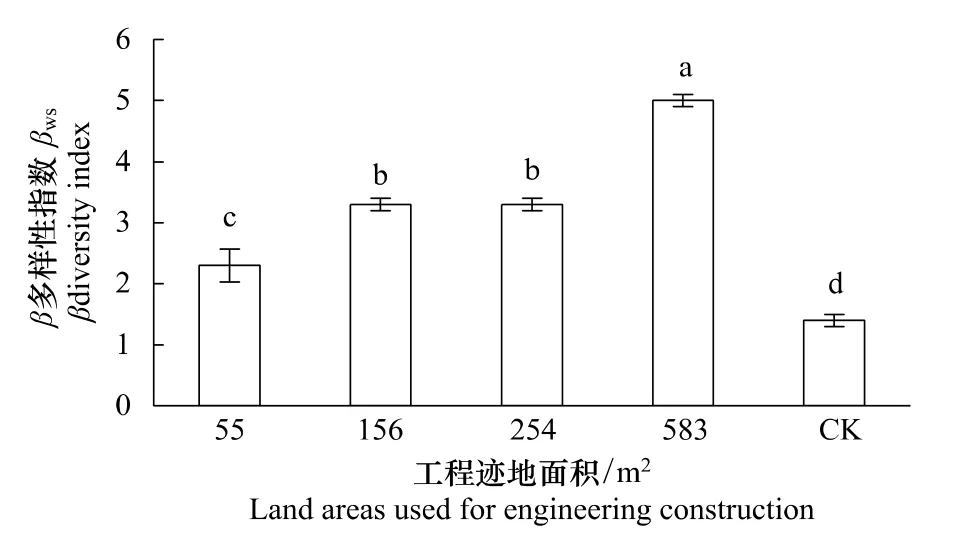
圖3 青藏高原高寒草原區不同面積工程跡地恢復群落的β多樣性指數Fig.3 β diversity index of restorable plant community at different land areas used for engineering construction in the alpine steppe of the Qinghai-Tibetan Plateau
2.4 工程跡地面積對植物地上生物量的影響
地上生物量是植物地上生產能力的綜合測度指標,更是退化草原恢復生產流程的物質基礎。研究結果表明,經過19a的恢復,工程跡地上植物地上生物量從無到有,但增加量隨著工程跡地面積變化而明顯不同(圖4),當工程跡地面積小于或等于156 m2時,其地上生物量和未干擾群落的生物量差異不顯著,說明已經基本恢復到未干擾的水平,而當工程面積大于或等于254 m2后,經歷相同時期恢復群落的生物量仍然顯著小于未干擾群落的生物量(P<0.05),工程跡地面積為583 m2和254 m2時恢復植物群落的生物量僅為未干擾群落的48%和49%,說明植物群落生物量的恢復存在一個工程跡地面積的臨界值。
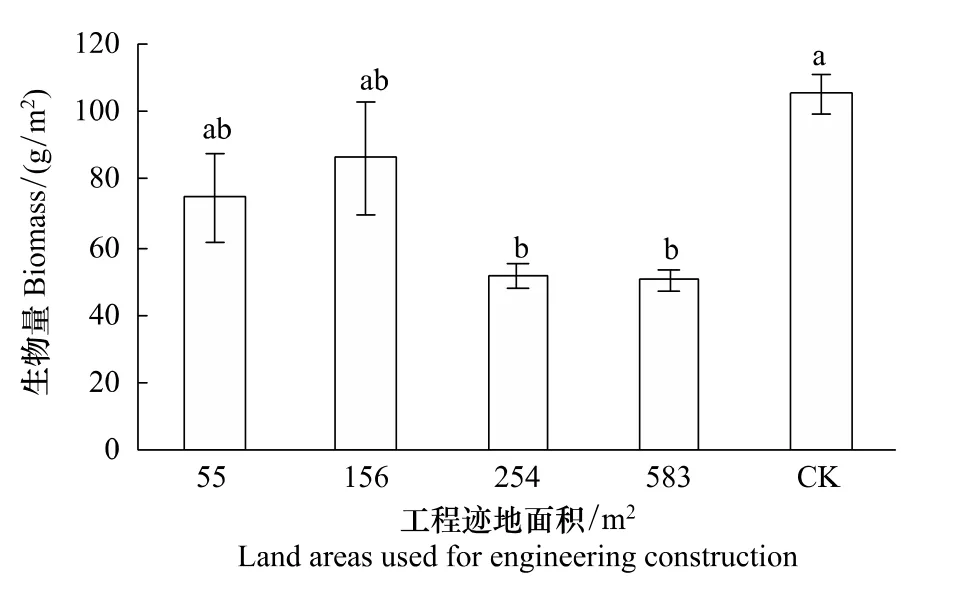
圖4 青藏高原高寒草原不同面積工程跡地上的生物量Fig.4 Biomass of restorable plant community at different land areas used for engineering construction in the alpine steppe of the Qinghai-Tibetan Plateau
2.6 工程跡地面積對群落恢復度的影響
群落恢復度是反應干擾跡地上植被恢復能力的綜合指標,綜合了植物多樣性,初級生產力和群落內種群分布特征的信息。試驗結果表明,經歷相同恢復時期,不同工程跡地面積植被恢復能力存在明顯差異(圖5),表現為隨工程跡地面積增加,SL值逐漸顯著減小(P<0.05),說明植物恢復能力越弱。
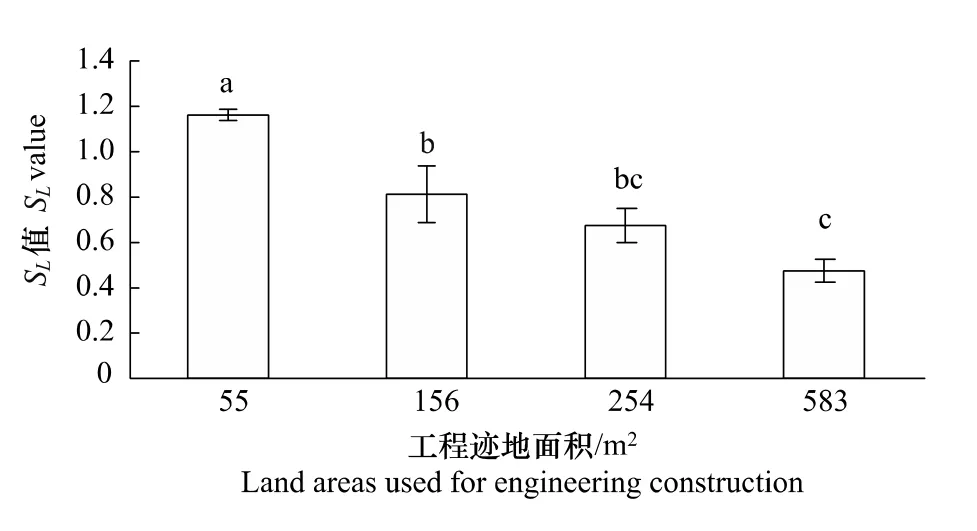
圖5 青藏高原高寒草原區不同面積工程跡地植被恢復能力Fig.5 Plant Restoration capability at different land areas used for engineering construction in the alpine steppe of the Qinghai-Tibetan Plateau
3 討論
3.1 工程跡地面積變化對高寒草原區恢復植物群落組分、結構和生產力的影響
受損草地生態系統在自然恢復過程中,通過逐漸優化和改善群落組分和結構,正向演替,這種過程貫穿植被恢復演替進程[14-15]。工程跡地產生過程具有突發特性,其植被恢復過程較不同于逐漸退化草地的恢復過程[3],這是因為工程跡地土壤內植物繁殖體較逐漸退化草地要少的多,而植物繁殖體萌發后是否健康生長,與工程跡地面積大小密切相關。本研究結果表明,經歷相同恢復時期,不同工程跡地面積上恢復植物群落的組分,蓋度,高度和生物量均存在明顯差異,表現為工程跡地面積越小,恢復植物群落的組分、空間結構和生產力指標越接近于未干擾群落的指標,說明其恢復較好,而面積越大,恢復程度越慢,這是因為工程跡地面積較小時,生境相對溫和,此時土壤內殘存成熟植物種子或其它繁殖構件,或者其他入侵物種的繁殖體,在適宜環境下萌發或繁殖,新生植物根系容易獲取土壤養分和水分,容易存活,正向演替,經過一定時期后逐漸形成穩定植物群落[16],而工程跡地面積較大時,很多植物繁殖體萌發后,由于裸斑面積太大,土壤蒸發大,特別是土壤淺層含水量較低,新生植物因不易獲取土壤養分和水分,在嚴酷環境中很快死亡,只有抗逆性較強的植物能夠殘活[3,17],但因其環境惡劣,生長過程較其在面積較小工程跡地上的過程要慢,這客觀上減緩了面積較大工程跡地上植物群落的自然恢復過程。當工程跡地面積小于156 m2時,群落優勢種由過渡性植物向原有群落演替,大量屬于未干擾草原群落的植物成分涌現,草原指示植物紫花針茅已零星出現,當工程跡地面積繼續減少到55 m2時,草原指示物種紫花針茅已成為主要伴生種,群落組分多樣化,伴生種數量、優勢種高度和重要值,群落高度和蓋度均明顯增加,說明工程跡地面積小于156 m2時,高寒草原受到干擾后具有較強的自我恢復能力[18]。而當工程跡地面積超過254 m2時,只有能夠存活于極端耐嚴酷條件的青藏苔草出現,而其余物種均為雜草,群落組分簡單,伴生種種類較少,群落蓋度和高度相對較低,優勢種高度較低,說明工程跡地面積超過254 m2時,工程跡地植物群落恢復能力較弱。工程跡地面積較大時,植物繁殖體定居及其種群擴張需要的時期可能更長,后者土壤水分和養分條件無法支持新定居種群的需要,實現恢復相對困難。采用綜合反映群落蓋度、頂級群落物種出現數與頻度的植被恢復能力綜合評價指標評價的結果再次印證了這個結果,從SL的分異性可以看出,工程跡地面積越小,其值越大,而工程跡地面積越小,其值越大,充分反映了青藏高原高寒草原區,面積較小的工程跡地較面積較大的工程跡地在經歷相同恢復期后,更容易形成穩定的群落組分體系[12],逐漸趨向于未受干擾的群落。
3.2 工程跡地面積變化對高寒草原區恢復群落植物多樣性的影響
植物多樣性是高寒草原維持穩產和生態系統健康的關鍵所在[3,11],工程跡地恢復群落的植物多樣性變化反應了其演替趨向。本研究結果表明,工程跡地面積對恢復群落的植物多樣性具有明顯影響,多樣性指數和豐富度指數隨著工程跡地面積減小而逐漸增大,其臨界發生在工程跡地面積為254 m2時,當工程跡地面積小于或等于此值時,雖然物種豐富度指數仍然小于未干擾群落,但已經達到未干擾群落的67%以上,面積越小,越接近于未干擾群落的物種豐富度,而多樣性指數基本恢復至未干擾群落的水平,群落內種群分布相對均勻[19],主要是工程跡地面積在一定的范圍內,小環境一致性增大,伴生種在較一致的環境中能夠很好的生長,各個種出現機率差異小。當工程跡地面積大于254 m2時,植物繁殖體因入侵途徑延長而變得困難[20],部分新生物種因無法適應土壤水分蒸發快的環境而死亡,只有能夠忍耐極端環境的短穗兔耳草、青藏苔草和矮火絨草等植物生長[11],導致群落樣方內物種數很少,僅為未干擾群落的30%—40%。
物種多樣性反映了群落內部物種數量的變化,而β多樣性則反映的是群落間多樣性在質方面的分異,其反映了不同群落間共有物種數的多少,揭示群落間物種的替代速率[21],兩個群落間共有物種數少,說明其分布地環境差異大[22]。β多樣性指數對工程跡地面積變化的響應結果表明,工程跡地面積越小,β多樣性指數逐漸減少,說明群落內物種替代速率越低,其和未干擾群落共有的物種數增加,工程跡地的環境條件越趨向于未干擾群落的環境[23],反之亦然。
恢復群落的α多樣性和β多樣性對工程跡地面積變化的響應過程說明,工程跡地恢復群落的植物多樣性恢復過程受控于面積,且面積的拐點出現在254 m2,當工程跡地面積大于此值時,植物多樣性恢復較慢,群落間物種替代速率高[24],其生境和未干擾群落的生境差異大[3],而當工程跡地面積小于此值時,經過19年的恢復,工程跡地α多樣性已基本接近未受干擾的草原群落,恢復群落和未受干擾群落間共有物種數逐漸增多,生境異質性減弱[15],說明青藏高原高寒草原區工程跡地植物多樣性的恢復不僅受時間的調控,而且受工程跡地面積大小的調控,當工程跡地面積小于254 m2時,經歷一定時期工程跡地上植物多樣性可以恢復,而當工程跡地面積大于254 m2時,植物多樣性恢復的時間可能更長或不易恢復。
致謝:蘭州大學草地農業科技學院的藕洋、馮成庸、王丹丹、張曉黎、趙旭同學參加野外調查工作,特此致謝。
[1] Ren Z H,Zhang Y G,Li D Q,Xiao Q M,Cai C Y.The soil microbial activities and microbial biomass in Sanjiangyuan alpine grassland.Acta Ecologica Sinica,2011,31(11):3232-3238.
[2] Han D R,Cao G M,Guo X W,Zhang F W,Li Y K,Lin L,Li J,Tang Y H,Gu S.The potential of carbon sink in alpine meadow ecosystem on the Qinghai-Tibetan Plateau.Acta Ecologica Sinica,2011,31(24):7408-7417.
[3] Guo Z G,Long R J,Niu F J,Wu Q B,Hu Y K.Effect of highway construction on plant diversity of grassland communities in the permafrost regions of the Qinghai-Tibet plateau.The Rangeland Journal,2007,29(2):161-167.
[4] Zhou G Y,Li T C,Xu W H,Sun J,Ma H,Chen G C.Characteristics analysis of mineral elements content in dominant species and soil of artificial restoration vegetation of Qinghai-Tibetan Plateau permafrost region.Resources and Environment in the Yangtze Basin,2011,20(2):191-196.
[5] Spellerberg I F.Ecological Effects of Roads.Plymouth,UK:Science Publishers Inc,2002.
[6] Liu C X,Han L B.Review of researches in vegetation restoration of freeway slopes.Acta Ecologica Sinica,2007,27(5):2090-2098.
[7] Cheng P,Ji W S.Research on the test of vegetation recover in summit area of Tanggula Mountain along Qinghai-Tibet railway.Journal of Railway Engineering Society,2008,25(1):48-52.
[8] Ma S Z,Chen G C,Peng M,Zhou G Y,Zhao Y L.The alpine steppe vegetation restoration process of fountainhead region of Yangtze River.China Environmental Science,2004,24(2):188-191.
[9] Zhu G H,Tao L,Ren J.Evaluation of using land for constructing Qinghai-Tibet railway on native vegetation.Acta Agrestia Sinica,2006,14(2):160-164.
[10] Yang S Z,Jin H J,Ji Y J,Wei Z,He R X.Revegetation in permafrost regions along a linear project.Journal of Glaciology and Geocryology,2008,30(5):875-882.
[11] Norris J E.Root reinforcement by hawthorn and oak roots on a highway cut-slope in Southern England.Plant and Soil,2005,278(12):43-53.
[12] Zhang G S,Shi X H,Li D l,Wang Q C,Dai S.Climate change in Tuotuohe area at the headwaters of Yangtze River.Journal of Glaciology and Geocryology,2006,28(5):678-685.
[13] Liu Y,Hu G,Liang S C,Peng M,Lu X F,Yue P P.Interspecific associations analysis of Stipa purpurea community in Tuotuo River area.Guihaia,2007,27(5):720-724.
[14] Guo Z G,Li X F,Liu X Y,Zhou X R.Response of alpine meadow communities to burrow density changes of plateau pika(Ochotona curzoniae)in the Qinghai-Tibet Plateau.Acta Ecologica Sinica,2012,32(1):44-49.
[15] Wang G X,Yao J Z,Guo Z G,Wu Q B,Wang Y B.Changes in permafrost ecosystem under the influences of human engineering activities and its enlightenment to railway construction.Chinese Science Bulletin,2004,49(16):1741-1750.
[16] Li D M,Guo Z G,An L Z.Assessment on vegetation restoration capacity of several grassland ecosystems under destroyed disturbance in permafrost regions of Qinghai-Tibet Plateau.Chinese Journal of Applied Ecology,2008,19(10):2182-2188.
[17] Huang Z L,Kun G H,He D Q.Plant community diversity in Dinghushan Nature Reserve.Acta Ecologica Sinica,2000,20(2):193-198.
[18] Hou Y,Guo Z G,Long R J.Changes of plant community structure and species diversity in degradation process of Shouqu wetland of Yellow River.Chinese Journal of Applied Ecology,2009,20(1):27-32.
[19] Han L H,Shang Z H,Ren G H,Wang Y L,Ma Y S,Li X L,Long R J.The response of plants and soil on black soil patch of the Qinghai-Tibetan Plateau to variation of bare-patch areas.Acta Prataculturae Sinica,2011,20(1):1-6.
[20] Wheeler A P,Angermeier P L,Rosenberger A E.Impacts of new highways and subsequent landscape urbanization on stream habitat and biota.Reviews in Fisheries Science,2005,13(3):141-164.
[21] Trombulak S C,Frissel C A.Review of ecological effects of roads on terrestrial and aquatic communities.Conservation Biology,2000,14(1):18-30.
[22] Guo Z G,Niu F J,Zhan H,Wu Q B.Changes of grassland ecosystem due to degradation of permafrost frozen soil in the Qinghai-Tibet Plateau.Acta Ecologica Sinica,2007,27(8):3294-3301.
[23] Katopodis C,Aadland L P.Effective dam removal and river channel restoration approaches.International Journal of River Basin Management,2006,4(3):153-168.
[24] Guo Z G,Liu H X,Wang G X,Cheng G D.Effect of the Qinghai-Tibetan Highway on the β diversity of grassland plant communities in the northern region of the Qinghai-Tibetan Plateau.Acta Ecologica Sinica,2004,24(2):384-388.
參考文獻:
[1] 任佐華,張于光,李迪強,肖啟明,蔡重陽.三江源地區高寒草原土壤微生物活性和微生物量.生態學報,2011,31(11):3232-3238.
[2] 韓道瑞,曹廣民,郭小偉,張法偉,李以康,林麗,李婧,唐艷鴻,古松.青藏高原高寒草甸生態系統碳增匯潛力.生態學報,2011,31(24):7408-7417.
[4] 周國英,李天才,徐文華,孫菁,馬海,陳桂琛.多年凍土區工程跡地人工恢復區植物和土壤的礦質元素含量特征.長江流域資源與環境,2011,20(2):191-196.
[6] 劉春霞,韓烈保.高速公路邊坡植被恢復研究進展.生態學報,2007,27(5):2090-2098.
[7] 陳鵬,季維生.青藏鐵路唐古拉山越嶺地段植被恢復試驗研究.鐵道工程學報,2008,25(1):48-52.
[8] 馬世震,陳桂琛,彭敏,周國英,趙以蓮.青藏公路取土場高寒草原植被的恢復進程.中國環境科學,2004,24(2):188-191.
[9] 祝廣華,陶玲,任珺.青藏鐵路工程跡地對植被的影響評價.草地學報,2006,14(2):160-164.
[10] 楊思忠,金會軍,吉延峻,魏智,何瑞霞.寒區線性工程沿線凍土區的植被恢復.冰川凍土,2008,30(5):875-882.
[12] 張國勝,時興合,李棟梁,汪青春,戴升.長江源沱沱河區45a來的氣候變化特征.冰川凍土,2006,28(5):678-685.
[13] 劉洋,胡剛,梁士楚,彭敏,盧學峰,岳鵬鵬.沱沱河地區紫花針茅群落種間聯結性分析.廣西植物,2007,27(5):720-724.
[16] 李東明,郭正剛,安黎哲.青藏高原多年凍土區不同草地生態系統恢復能力評價.應用生態學報,2008,19(10):2182-2188.
[17] 黃忠良,孔國輝,何道泉.鼎湖山植物群落多樣性的研究.生態學報,2000,20(2):193-198.
[18] 后源,郭正剛,龍瑞軍.黃河首曲濕地退化過程中植物群落組分及物種多樣性的變化.應用生態學報,2009,20(1):27-32.
[19] 韓立輝,尚占環,任國華,王彥龍,馬玉壽,李希來,龍瑞軍.青藏高原“黑土灘”退化草地植物和土壤對禿斑面積變化的響應.草業學報,2011,20(1):1-6.
[22] 郭正剛,牛富俊,湛虎,吳青柏.青藏高原北部多年凍土退化過程中生態系統的變化特征.生態學報,2007,27(8):3294-3301.
[24] 郭正剛,劉慧霞,王根緒,程國棟.人類工程對青藏高原北部草地群落β多樣性的影響.生態學報,2004,24(2):384-388.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
太空探索(2016年6期)2016-07-10 12:09:06
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
筑路機械與施工機械化(2015年11期)2015-07-01 16:28:43
