施氮對木荷3個種源幼苗根系發育和氮磷效率的影響
2013-12-16 08:20:38金國慶周志春豐忠平
生態學報 2013年12期
張 蕊,王 藝,金國慶,周志春,*,豐忠平
(1.中國林業科學研究院亞熱帶林業研究所,富陽 311400;2.浙江省淳安縣姥山林場,淳安 311700)
近年來隨著人類活動及工業污染的加劇,大氣氮沉降成為人們關注的重大環境問題之一。目前我國無機氮的排放量已逐漸達到一個較高水平。遆超普指出,我國南方氮沉降量在2007年就已達到45 kg N hm-2·a-1以上,比1980年增長了2倍多[1]。研究指出,較低水平的氮素可以促進植株生長[2-6],如膏桐(Jatropha curcas)[7]、黑楊 (Populus nigra)[8]、長白落葉松 (Larix algensis)[9]、西南樺 (Betula alnoides)[10]和馬褂木(Liriodendron chinense)[11]等主要造林樹種在低氮處理下其地徑、苗高、干物質積累量均明顯增加,地下部與地上部比率增大,當施氮量達到一定水平后,植株整體水平則趨于穩定后下降,說明高水平的氮素處理會對植株造成明顯的抑制作用。氮沉降向土壤中輸入了大量的氮素[12-13],這類似于人工施肥,造成土壤營養環境的改變,影響了植株根系的生長發育[14-16]。對長白落葉松的研究表明,低水平氮素增加了細根的根尖數、較粗根(2—5mm)的長度、根表面積和體積,高氮處理下根系生長受抑[9]。從根系發育角度研究氮沉降對林木生長的影響,了解植株對營養元素的吸收利用情況,充分發掘林木有效利用土壤有限養分的潛力意義重大。
木荷(Schima superba)是山茶科木荷屬常綠闊葉大喬木,為亞熱帶地帶性常綠闊葉林的主要建群種,也是我國南方主栽的生態防護樹種和珍貴用材樹種,造林成效好,優質速生,豐產性顯著。本研究組自2001年來,開展了木荷的多目標育種,發現其存在豐富的種源和家系變異,具有較強的覓取利用斑塊養分能力的特性,同時揭示了水分和磷素對木荷種源生長的影響,已初選出一批優良種源和家系[17-20]。有研究認為適量的氮素處理會增加木荷幼苗生長量,促進凈光合速率和水分利用效率,但卻降低了根系在全株的分配比例等[4-5,21]。本文擬通過人工控制施氮,研究不同地理種源木荷在氮素增多情況下的生長表現,了解幼苗根系生長發育和氮磷利用效率的變化,探討氮素對木荷不同種源苗木根系生長發育的影響及機制,進一步闡明不同種源苗木對不同水平氮沉降的響應差異,為在大氣N沉降環境背景下,選育營養高效利用的木荷速生優質新品種提供理論依據。
1 材料和方法
1.1 試驗材料
我國南方土壤多為紅壤和棕紅壤,其養分含量較低,并呈現出緯向變異模式[22]。同時,我國南方氮沉降豐富,據文獻資料,2005年浙江杭嘉湖地區氮沉降量約為30.9 kg hm-2a-1[23],江西鷹潭地區氮沉降量約為30.7 kg hm-2a-1[24],而大崗山地區氮沉降量高達 57 kg hm-2a-1[25],由于工業生產和農業耕作的影響,氮沉降量大小具有地域差異。據統計,2007年浙江、福建和江西三省的氮沉降平均水平分別為32.7、27.8和22.2 kg hm-2a-1[1]。因此,以浙江杭州、福建建甌和江西信豐3個不同緯度的木荷代表性種源為試驗材料,以林地貧瘠土壤為基質,開展不同施氮水平的盆栽試驗。實驗選用的盆栽基質取自浙江省淳安縣姥山林場的酸性貧瘠紅壤,其有機質含量為6.68 g/kg,全氮和全磷含量分別為0.41 g/kg和0.35 g/kg,水解氮、速效鉀和有效磷含量分別為 46.35、91.11 和 5.49 mg/kg,pH5.06。
1.2 試驗設計
試驗以全株噴施NH4NO3溶液來進行施氮處理。分為4個處理組,分別為N0(對照0 kg N hm-2·a-1)、N50(低氮,50 kg N hm-2·a-1)、N100(中氮,100 kg N hm-2·a-1)和N200(高氮,200 kg N hm-2·a-1)。每處理30株,共計360株。選用上端內徑16cm,下端內徑13.5cm,高18cm的營養杯作為盆栽容器。每營養杯裝土約3.75kg,整個盆栽試驗在浙江省淳安縣姥山林場實驗大棚內進行。木荷種源試驗種子于2011年3月7日播種,5月中旬,將生長整齊一致的芽苗移栽,每盆移栽2株芽苗,成活后保留1株,幼苗在充分供水條件下培養。從2011年5月下旬到11月上旬,以設計施氮量的18次均分后,每月月初和月中分2次向幼苗全株噴施NH4NO3,其中7—9月補加2次直至收獲。
1.3 試驗采收及指標測定
試驗于2011年11月上旬收獲。每個處理選取9株生長正常的盆栽苗木,量測其苗高和地徑,并用根系圖象分析系統(WINRHIZO ProSTD1600+型,加拿大REGENT公司)測定根系總長、根系平均直徑、根總表面積和根系總體積等根系參數。同時植株分根、莖、葉,經105℃殺青1h后80℃烘干至恒量,測定各部分的干物質量。用濃H2SO4-H2O2消煮-鉬銻抗比色法和凱氏定氮法分別測定各部分磷、氮含量[26-27]。
1.4 數據處理
以單株測定值為單元,利用SAS軟件ANOVA程序進行性狀方差分析,并用DUNCAN法進行多重比較,以檢驗種源、氮素及其互作效應的顯著性。磷素吸收效率=各器官或整株磷素吸收量/各器官或整株;磷素利用效率=各器官或整株干物質積累量/各器官或整株的磷素吸收量;氮素吸收及利用效率同上。
2 結果與分析
2.1 施氮對木荷不同種源幼苗生長及干物質積累量影響
從施氮處理下木荷種源苗高和地徑的生長結果看出(表1),施氮對木荷苗木的生長具有極顯著的促進作用,其中苗高在中氮條件下達到最大值(較對照高34%),而地徑在低氮條件下即達到最大值(較對照高19%),但苗高和地徑在高氮條件下均呈現下降趨勢。木荷幼苗干物質積累量在施氮條件下呈現增長趨勢,且在中、高氮條件下達到最大,說明在貧瘠土壤環境下,施氮有效地促進了木荷苗木的生長。3個木荷種源幼苗生長對氮素表現出的敏感程度不同。如浙江杭州和江西信豐種源在低氮水平下表現出顯著的生長勢,其中苗高較對照分別增長了34%和44%,地徑分別增長了26%和33%,這種生長優勢顯著高于同水平氮量下福建建甌種源,同時改變了3個種源原有的生長規律表現,即在低氮處理下,處于木荷中心產區,且具有高生長速率的福建建甌種源的生長下降,而北部浙江杭州和南部江西信豐種源生長提高。福建建甌種源則在中、高氮水平下表現出顯著的生長優勢,苗高和地徑分別較對照增長了46%、19%和27%、15%,說明中心產區的福建建甌種源地上部生長對低水平氮添加反應敏感性一般。隨著氮水平的提高,3個種源又恢復為中心產區高而南北部種源低的生長規律。
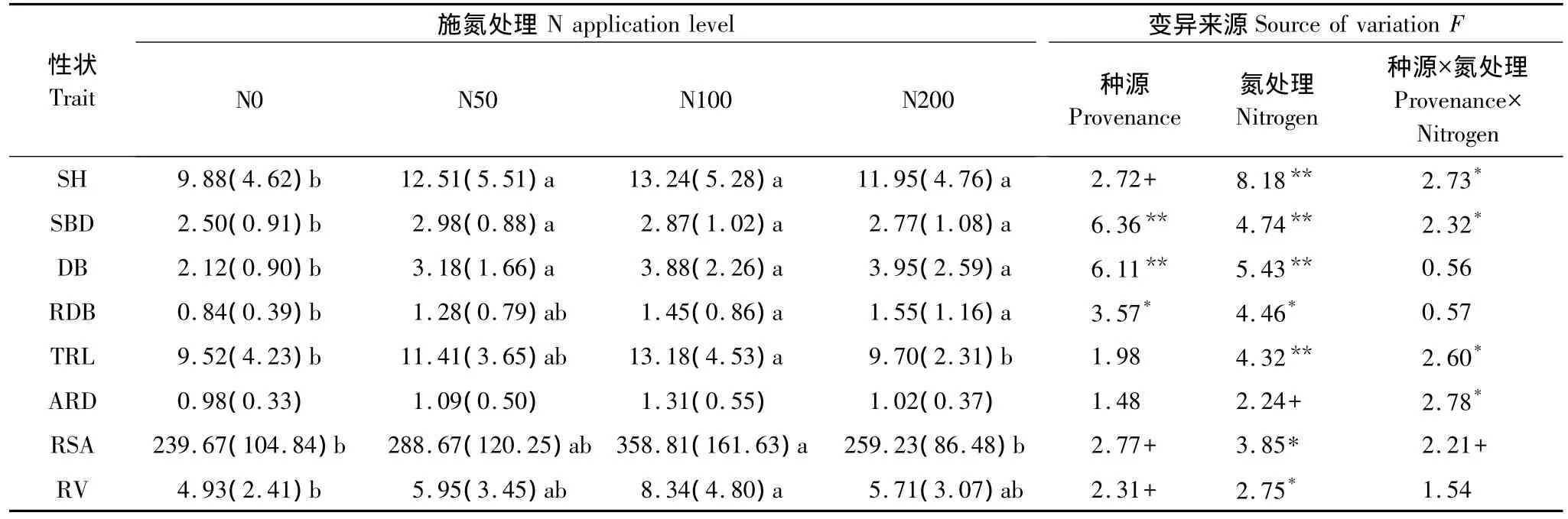
表1 施氮對木荷生長和根系參數的影響(平均值(標準偏差),n=9)Table 1 Effects of N addition on seedling growth and root characters of S.superba(mean(SD),n=9)
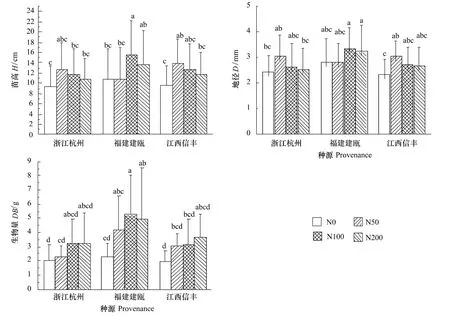
圖1 施氮對3個木荷種源苗高、地徑和生物量的影響(平均值(標準偏差),n=9)Fig.1 Effects of nitrogen addition on seedling height(H),stem base diameter(D)and dry biomass(DB)of three provenances of S.superba(mean(SD),n=9)
2.2 施氮對不同種源木荷苗木根系生長發育影響
木荷根系發達,較多的側根數量滿足了其在貧瘠土壤中的營養利用。氮添加不僅影響了木荷地上部的生長,同時也改變其根系的生長發育。在施氮處理下,隨著氮水平的提高,木荷幼苗根系干物質量呈現上升趨勢,并在中、高氮水平下達到最高,根系總長、根系平均直徑、根總表面積和根總體積則分別在中氮水平下最大(表1),其中>0.5mm直徑的根系生長最為顯著,說明在貧瘠土壤環境下,增加的氮素刺激了根系的生長,尤其促進了側根的增粗生長,根系體積增大,根系粗壯舒張。隨著氮水平的升高,高氮沉降則抑制了木荷幼苗根系生長。
在貧瘠土壤環境下,3個種源木荷幼苗根系生長對施氮處理的敏感程度不同(圖2,圖3,表2)。北部浙江杭州種源對中、高氮反應強烈,根系干物質量、總長、平均直徑、總表面積和總體積較對照增長了3%—75%,平均增長了 33%,0.5—1.0 mm 和>1.0 mm 直徑的根長平均分別增加了16%和43%,而福建建甌和江西信豐種源則在中、低氮水平下根系生長反應就已較為突出,中部福建建甌種源增長幅度較大,根系干物質量增長平均為2.13倍,4個根系參數增長量平均在54%,0.5—1.0 mm 和>1.0 mm 直徑的根長平均分別增加了55%和63%。南部江西信豐種源根系干物質量增長了49%,4個根系參數平均增長量僅有4%,>1.0 mm直徑的根系較對照增加7%以上。說明氮素對中南部種源根系生長作用明顯,尤其中部福建建甌種源根系生長發育潛力更大,而氮素對北部種源木荷根系生長作用較弱。因此,不同種源木荷幼苗根系生長對氮素反應不同,造成了其在自然環境中適應性的改變,這有可能最終改變不同木荷種源苗木對外界環境的適應能力,導致種源之間生長差異,故而在育種時要充分考慮氮素對其根系生長的影響。
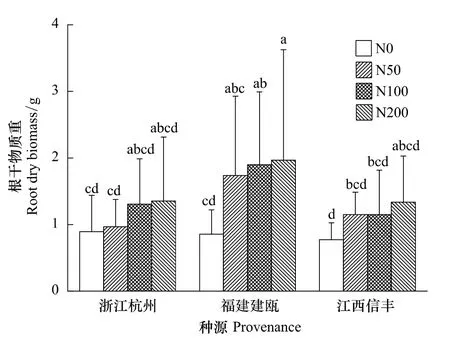
圖2 施氮對3個木荷種源苗木根系干物質量的影響Fig.2 Effects of nitrogen addition on root dry biomass of three provenances of S.superba
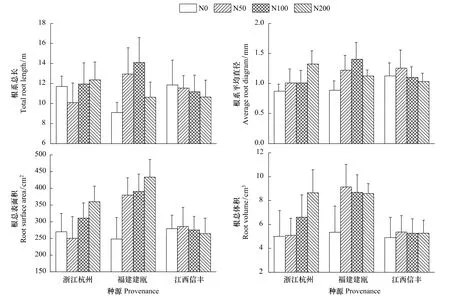
圖3 施氮對3個木荷種源苗木根系生長發育的影響Fig.3 Effects of nitrogen addition on seedling root growth of three provenances of S.superba
2.3 施氮對木荷不同種源苗木氮磷效率的影響
施氮不僅改變了木荷幼苗根系的形態特征,而且影響了根系的氮磷營養的吸收和利用效率,且在種源間差異顯著(表3)。中、低氮處理增加了木荷幼苗的氮磷吸收效率和利用效率,但高氮水平下,磷素吸收效率開始下降,氮素利用效率則低于對照,說明施氮促進了木荷幼苗對貧瘠低磷土壤中磷素的吸收利用,同時由于土壤氮素的提高,其對氮素營養的吸收利用也開始增強。種源間變異規律和根系生長發育相似,浙江杭州種源在中、高氮水平下磷素吸收利用效率較高,平均增長23%以上,低氮處理降低了杭州種源磷素吸收效率,但磷素利用效率有所提高;中、低氮處理對福建建甌和江西信豐種源磷素吸收和利用效率促進作用顯著,但氮利用效率在中氮水平時則開始降低。
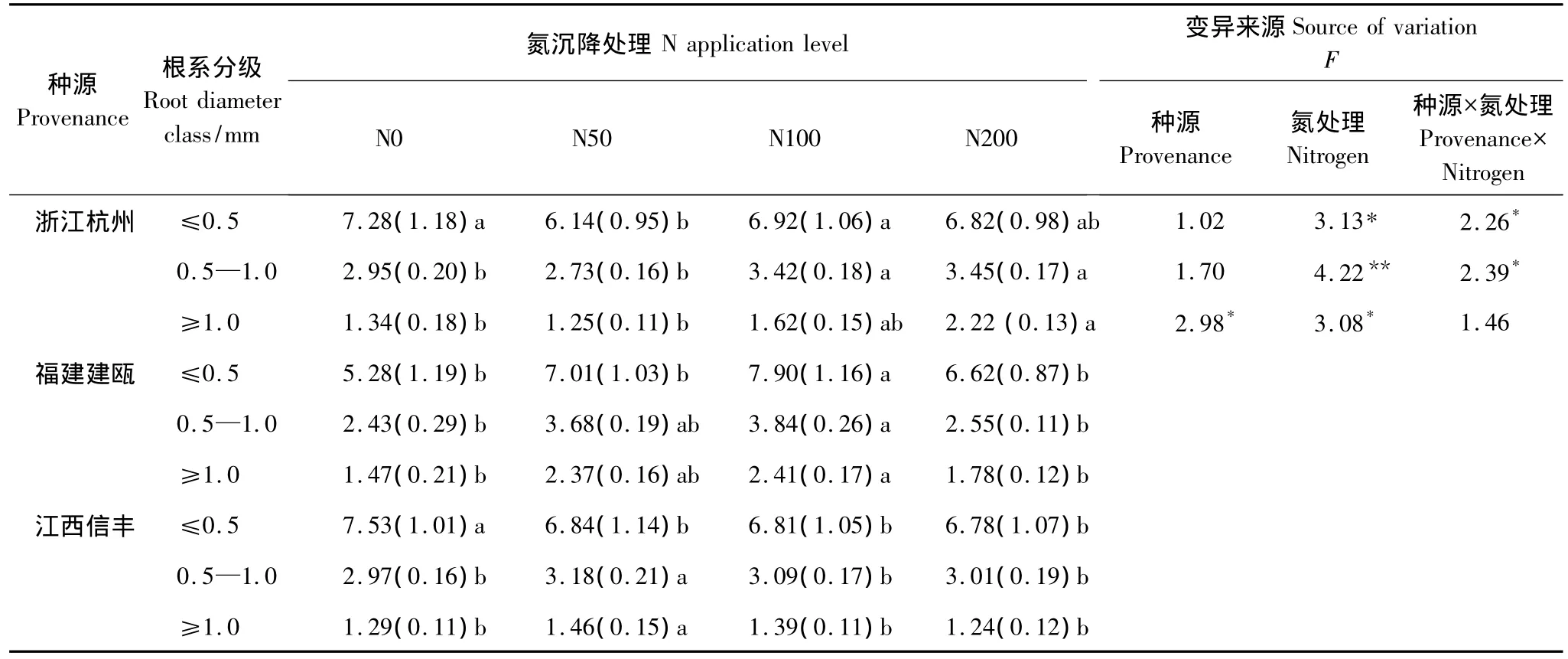
表2 施氮處理對3個木荷種源苗木不同徑級根長的影響(平均值(標準偏差),n=9)Table 2 Root length(m/plant)per diameter class of three provenances of S.superba in Nitrogen addition experiment(mean(SD),n=9)

表3 施氮處理中3個木荷種源苗木的氮磷效率Table 3 Nitrogen and phosphorus efficiency of three provenance of S.superba in Nitrogen addition experiment
表4結果說明,木荷幼苗根系氮磷吸收利用效率在不同氮沉降水平下受根系結構參數影響顯著。隨氮水平的提高,PAE和NAE與根系總長、根系體積、根系平均直徑和總表面積相關性增強,在中氮水平下相關性達極顯著水平。在中氮水平下,木荷幼苗根系NAE受>0.5 mm根長的作用顯著,而PAE則與3種不同直徑根系根長均顯著相關,說明木荷根系對氮磷吸收的敏感部位不盡相同,這也可能和貧瘠土壤中磷素缺乏有關。氮素吸收效率極顯著影響了磷素吸收效率,同時氮磷素吸收效率又和木荷生物量呈顯著正相關關系,從而最終導致木荷植株在生長上的差異。
3 討論
本研究的結果表明,在林地貧瘠土壤條件下,施氮對木荷幼苗生長產生了不同程度的促進作用,類似于其他研究結果,說明低水平的氮沉降有利于木荷幼苗的生長[2-3,6,20],但隨著氮水平提高,高氮作用下木荷幼苗生長速度減緩,抑制作用開始顯現。由于實驗土壤較為瘠薄,在50 kg N hm-2·a-1的氮水平時,木荷幼苗就已表現出明顯的生長勢,而地下部根系生長發育最大值仍出現在100 kg N hm-2·a-1的中氮水平,這說明在貧瘠土壤環境下,增加的氮素不僅改變了土壤的營養環境,而且噴施于葉片上的氮素可被木荷幼苗直接吸收利用,地上部首先受到影響且第一時間做出反應,由于氮素與光合作用密切相關,氮素的增加促進了葉片生長,葉綠素增多,光合作用增強,從而增加了生物量積累[28],之后營養物質供給到根部,促進根系生長,造成根系對氮素反應相對滯后。
木荷適應性較強,其發達的須側根保障了其在多種立地環境條件下生長,在養分競爭中具有一定的優勢[29]。本實驗結果證實,氮沉降增加了木荷根系干物質量,根系總長度、平均直徑、總表面積和體積亦有所增加,木荷幼苗根系更為粗壯舒張,且較粗根數量開始增多,密度加大,相關分析證實這種根系結構和木荷幼苗的氮磷吸收效率呈顯著正相關,即這種根系結構更有利于對土壤營養元素的吸收。同時,氮磷吸收效率的提高促進了木荷幼苗干物質量的積累,并最終影響木荷生長。有研究認為,氮素可作為根系發育的局部信號,直接刺激和誘導側根的發生和生長,通過增加根尖區細胞數量,刺激側根伸長,同時氮素的增加會造成植株細根生物量和根尖數的減少[30-34]。同時,葉片中高濃度的NO-3亦可對側根生長發育進行調控[35]。但土壤氮飽和后過量的氮素會對植株造成傷害,Zhang等認為當外界NO-3濃度超過10mmol/L時,植物側根發育受抑,地上部生長發育阻滯[31]。從土壤營養狀況來看,該實驗土壤基質取自森林中的貧瘠土壤,氮磷素含量較低,施氮后補充了土壤氮素養分,使得土壤N/P比率升高。研究顯示,植物體內N/P比具有一定的范圍,且該值和土壤N、P水平相關,具有指示植物是否受到營養限制的作用,一般認為該值<14時,植物受到氮素限制,>16時受到磷素限制[36-37]。施氮處理后,木荷葉片在中氮水平時N/P約為16.0,而高氮處理下則約為17.7,顯然,隨著施氮水平的提高,木荷生長更多的受到磷素的限制。植物為了保持和土壤相同水平的N/P比,因而調節代謝機制,降低了磷素的吸收,增加氮素吸收,葉片營養缺乏導致植物光合、糖代謝、碳水化合物庫源分配產生變化,碳素更多的向地下部分配,導致了根部生物量的提高[38-39]。因此,不同水平氮沉降對木荷幼苗生長發育的影響,關鍵在于植株生長土壤的營養狀況。
氮沉降下,木荷不同種源幼苗苗高、地徑、干物質量、根系參數和氮磷吸收利用效率皆存在顯著的種源差異,參試的3個種源分別來自浙北、閩北和贛南,分屬木荷不同產區,具有較大的緯度差異。試驗結果表明,木荷北部產區的杭州種源生長在整體上對施氮反應敏感,施氮初期,該種源地上部生長即已表現出優勢,而地下根系生長緩慢,隨著施氮水平的提高,地上部生長下降,而根系開始加速生長;中心產區的福建建甌種源,在低水平氮沉降時根系生長旺盛,在氮水平達到一定程度后,地上部才表現出旺盛的生長勢,生長表現相對滯后;南部產區的江西信豐種源地上部表現和浙江杭州種源相似,但地下部根系始終處于緩慢或抑制生長狀態。木荷種源試驗結果認為,福建建甌屬木荷中心產區,該區域氣候適宜,水熱資源豐富,立地條件優良,長期適應的結果致使福建建甌木荷種源生長迅速,生物積累量大[17,40]。該區域范圍的馬尾松(Pinus massoniana)[41]和杉木(Cunninghamia lanceolata)[42]種源等亦有類似結果。這種長期適宜環境因子作用下形成的速生基因型,對低水平氮沉降的生長反應較小,只有在氮水平達到一定范圍后,其體內N、P養分平衡發生改變,植株會根據機體需求吸收利用額外的氮磷營養,最終導致加速或抑制生長發育;相反,浙江杭州和江西信豐種源分別位于非中心產區和次中心產區,生長環境因子相對較差,適度的營養刺激則會表現出較為明顯的生長勢,但整體生長表現仍低于中心產區種源。因此,現階段對木荷的育種工作要充分考慮到氮沉降的影響,才能更好地適應環境改變。

氮素和磷素是植物生長所需大量元素,對植物的生長發育影響顯著,施肥即是人們通過提高氮素和磷素在土壤中的配比含量來達到增產的目的,但事實證明過度施肥是造成減產的原因之一。本實驗中模擬大氣氮沉降對植物生長的促進效應可獲得更多可利用的優良種源,達到速生增產的目的,但是長期的氮沉降環境下,林木是否依然會保持旺盛的生長,積累更多的干物質量仍需進行長期觀測研究,以進一步更客觀的揭示大氣氮沉降下,氮素和磷素互作對木荷種源苗木生長發育的作用機理。
[1] Ti C P,Yan X Y.Estimation of atmospheric nitrogen wet deposition in China mainland from based on N emission data.Journal of Agro-Environment Science,2010,29:1606-1611.
[2] Nakaji T,Fukami M,Dokiya Y,Izuta T.Effects of high nitrogen load on growth,photosynthesis and nutritrient status of Cryptomeria japonica and Pinus densiflra seedlings.Trees,2001,15:4533-461.
[3] Nakaji T,Takenaga S,Kuroha M,Izuta T.Photosynthetic response of Pinus densiflora seedling to high nitrogen load.Environmental Sciences,2002,9:269-282.
[4] Li D J,Mo J M,Fang Y T,Li Z A.Effects of simulated nitrogen deposition on biomass production and allocation in Shima superba and Cryptocarya concinna seedlings in subtropical China.Chinese Journal of Plant Ecology,2005,29:543-549.
[5] Mo J M,Li D J,Gundersen P.Seedling growth response of two tropical tree species to nitrogen deposition in southern China.European Journal of Forest Research,2008,127:275-283.
[6] Wu Q,Ding J,Yan H,Zhang S R,Fang T,Ma K P.Effects of simulated precipitation and nitrogen addition on seedling growth and biomass in five tree species in Gutian Mountain,Zhejiang Province,China.Chinese Journal of Plant Ecology,2011,35:256-267.
[7] Peng M J,Lang N J,Wu T,He L P,Zheng K,Xiang Z Y,Kong J J,Yuan R L.Growth response of Jatropha curcas seedlings to different levels of nitrogen supply.Journal of Northwest Forestry University,2010,25:97-100.
[8] Liu X H,Ding C J,Zhang W X,Li W W,Huang Q J,Su X H.Study on difference and primarily mechanism of nitrogen use efficiency(NUE)in Populus nigra seedlings genotypes.Forest Research,2010,23:368-374.
[9] Jiang L N,Wei H X,Liu Y,Cheng C Y,Ma L Y.Relationships between root morphological plasticity and heterogeneous N for Larix algensis seedlings.Journal of northeast forestry university,2010,38:24-27.
[10] Chen L,Zeng J,Xu D P,Zhao Z G,Guo J J,Lin K Q,Sha E.Effects of exponential nitrogen loading on growth and foliar nutrient status of Betula alnoides seedlings.Scientia Silvae Sinicae,2010,46:35-40.
[11] Fan R H,Yang S P,Zhou Z C,Jin G Q.Effect of nitrogen on the seedling growth of Liriodendron Chinese families.Forest research,2009,22(1):85-90.
[12] Holland E A,Braswell B H,Sulzman J.Nitrogen Deposition Onto The United States And Western Europe:Synthesis Of Observations And Models.Ecological Applications,2004,15(1):38-57.
[13] Pearson J,Stewart G R.The deposition of atmospheric ammonia and its effects on plants.New Phytologist,1993,125:283-305
[14] Galloway J N,Cowling E B.Reactive nitrogen and the world:200 years of change.Ambio,2002,31:64-71.
[15] Linkohr B I,Williamson L C,Fitter A H,Leyser H M.Nitrate and phosphate availability and distribution have different effects on root system architecture of Arabidopsis.Plant Journal,2002,29:751-760.
[16] Desnos T.Root branching responses to phosphate and nitrate.Plant Biology,2008,11:82-87.
[17] Zhou Z C,Fan H H,Jin G Q,Wang Y S,Chen Y L,Rao L B.Geographic Genetic Variation and Preliminary Selection of Superior Provinance in Schima superba.Forest Research,2006,19:718-724.
[18] Lin L,Zhou Z C.Effects of soil moisture condition and phosphorus supply on the seedlings growth and phosphorus efficiency of Schima superba provenances.Chinese Journal of Applied Ecology,2009a,20:2617-2623.
[19] Lin L,Zhou Z C,Fan H H,Jin G Q,Chen Y L,Hong G M.Provenance difference in stable carbon isotope discrimination of Schima superba.Chinese Journal of Applied Ecology,2009b,20:741-746.
[20] Lin L,Zhou Z C,Fan H H,Jin G Q,Feng J G,Chen L Y,Wang Y S.Inheritance and Variation of Seedling Growth Traits of Open-pollinated Families from Plus-tree in Schima superba.Forest Research,2009,22:155-160.
[21] Li D J,Mo J M,Fang Y T,Cai X A,Xue J H,Xu G L.Effects of simulated nitrogen deposition on growth and photosynthesis of Schima superba,Castanopsis chinensis and Cryptocarya concinna seedlings.Acta Ecologica Sinica,2004,24:876-882.
[22] Lu R K.General status of nutrients(N,P,K)in soils of China.Acta Pedologica Sinica,1989,26(3):280-286.
[23] Chen Y,Tang X,Yang M S,Wu CY,Wang J Y.Atmospheric N wet deposition in Hangzhou region under rice-wheat-vegetable cropping system.Acta Ecologica Sinica,2009,29(11):6102-6109.
[24] Wang T J,Liu Q,Zhao H,Zhou J,Fan J L.Atmospheric nitrogen deposition in agrecosystem in red soil region of Jiangxi province.Acta Pedologica Sinica,2008,45(2):280-287.
[25] Ma X H.The ion elution effect of the rainwater in Pinus massaniana and Cunninghamia lanceolata plantation.Acta Ecologica Sinica,1989,9(1):15-20.
[26] Bremner J M,Mulvaney C S.Nitrogen-total.In:Page AL,Miller RH,Keeney DR,eds.Methods of soil analysis.Part 2:chemical and microbial properties,Agronomy Monograph 9.Agronomy Society of America,Madison,1982,595-624.
[27] Anderson J M,Ingram J S I.Tropical soil biology and fertility:a handbook of methods.2th eds.CAB International,Wallingford press,1993.
[28] Lu X K,Mo J M,Li D J,Zhang W,Fang Y T.Effects of simulated N deposition on the photosynthetic and physiologic characteristics of dominant understory plants in Dinghushan mountain of subtropical China.Journal of Beijing Forestry University,2007,29:1-9.
[29] Ma X H,Zhou Z C,Jin G Q,Zhang Y.Effects of competition on foraging behavior of Pinus massoniana and Schima superba in a heterogeneous nutrient environment.Chinese Journal of Plant Ecology,2009,33:81-88.
[30] Persson H,Ahlstr?m K,Clemensson A L.Nitrogen addition and removal at G?rdsj?n-effects on fine-root growth and fine-root chemistry.Forest Ecology and Management,1998,101:199-206.
[31] Zhang H M,Jennings A,Barlow P W,Forde B G.Dual pathways for regulation of root branching by nitrate.Proceedings of the National Academy of Sciences of the USA,1999,96:6529-6534.
[32] Sas L,Rengel Z,Tang C X.The effect of nitrogen nutrition on cluster root formation and proton extrusion by Lupinus albus.Annals of Botany,2002,89:435-442.
[33] Liu P W,Ivanov I I,Filleur S,Gan Y,Remans T,Forde B G.Nitrogen regulation of root branching.Annals of Botany,2006,97:875-881.
[34] Remans T,Nacry P,Pervent M,Girin T,Tillard P,Lepetit M,Gojon A.A central role for the nitrate transporter NRT2.1 in the integrated morphological and physiological responses of the root system to nitrogen limitation in Arabidopsis.Plant physiology,2006,140:909-921.
[35] Maga?a R H,Adamowicz S,Pagès L.Diel changes in nitrogen and carbon resource status and use for growth in young plants of tomato(Solanum lycopersicum).Annals of Botany,2009,103:1025-1037.
[36] Koerselman W,Meuleman A F M.The vegetation N∶P ratio:a new tool to detect the nature of nutrient limitation.Journal of Applied Ecology,1996,33:1441-1450.
[37] Han W X,Fang J Y,Guo D L,Zhang Y.Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China.New Phytologist,2005,168:377-385.
[38] Hermans C,Hammond J P,White P J,Verbruggen N.How do plants respond to nutrient shortage by biomass allocation?.Trends in Plant Science,2006,11:610-617.
[39] Marschner P.Marschner's Mineral Nutrition of Higher Plants.3th edn.Academic Press,2012.
[40] Wang X H,Chen L Y,Ma L Z,Fan H H,Ye S W,Zhou Z C.Geographical provenance variation of growth and wod basic density of 7-year-old Schina superba and its provenance selection.Forest Research,2011,24(3):307-313.
[41] Liu Q H,Jin G Q,Zhang R,Chu D Y,Qin G F,Zhou Z C.Provenance variation in growth,stem-form and wood density of masson pine at 24-year-old and the provenance division.Scientia Silvae Sinicae,2009,45:55-61.
[42] Sun C Z,Xie G N,Li P.Variations of wood properties and characheristics and a tentative selection for structural timbers of Cunninghamia lanceolata provenances.Scientia Silvae Sinicae,1993,29:429-436.
參考文獻:
[1] 遆超普,顏曉元.基于氮排放數據的中國大陸大氣氮素濕沉降量估算.農業環境科學學報,2010,29:1606-1611.
[4] 李德軍,莫江明,方運霆,李志安.模擬氮沉降對南亞熱帶兩種喬木幼苗生物量及其分配的影響.植物生態學報,2005,29(4):543-549.
[6] 吳茜,丁佳,閆慧,張守仁,方騰,馬克平.模擬降水變化和土壤施氮對浙江古田山5個樹種幼苗生長和生物量的影響.植物生態學報,2011,35:256–267.
[7] 彭明俊,郎南軍,吳濤,和麗萍,鄭科,向正勇,孔繼君,袁瑞玲.不同供氮水平對膏桐幼苗生長的影響.西北林學院學報,2010,25(3):97-100.
[8] 劉希華,丁昌俊,張偉溪,李文文,黃秦軍,蘇曉華.不同基因型歐洲黑楊幼苗氮素利用效率差異及其機理初探.林業科學研究,2010,23(3):368-374.
[9] 江俐妮,魏紅旭,劉勇,徐程揚,馬履一.長白落葉松播種苗根系形態可塑性與氮素空間異質性關系.東北林業大學學報,2010,38(1):24-27.
[10] 陳琳,曾杰,徐大平,趙志剛,郭俊杰,林開勤,沙二.氮素營養對西南樺幼苗生長及葉片養分狀況的影響.林業科學,2010,46(5):35-40.
[11] 樊瑞懷,楊水平,周志春,金國慶.氮素營養對馬褂木家系苗木生長效應分析.林業科學研究,2009,22(1):85-90.
[17] 周志春,范輝華,金國慶,王月生,陳奕良,饒龍兵.木荷地理遺傳變異和優良種源初選.林業科學研究,2006,19,718-724.
[18] 林磊,周志春,范輝華,金國慶,陳奕良,洪桂木.木荷穩定碳同位素分辨率的種源差異.應用生態學報,2009,20(4):741-746.
[19] 林磊,周志春.水分和磷素對木荷不同種源苗木生長和磷效率的影響.應用生態學報,2009,20(11):2617-2623.
[20] 林磊,周志春,范輝華,金國慶,馮建國,陳柳英,王月生.木荷優樹子代苗期生長遺傳和變異研究.林業科學研究,2009,22,155-160.
[21] 李德軍,莫江明,方運霆,蔡錫安,薛璟花,徐國良.模擬氮沉降對三種南亞熱帶樹苗生長和光合作用的影響.生態學報,2004,24(5):876-882.
[22] 魯如坤.我國土壤氮、磷、鉀的基本狀況.土壤學報,1989,26(3):280-286.
[23] 陳義,唐旭,楊生茂,吳春艷,王家玉.杭州稻麥菜輪作地區大氣氮濕沉降.生態學報,2009,29(11):6102-6109.
[24] 王體鍵,劉倩,趙恒,周靜,樊建凌.江西紅壤地區農田生態系統大氣氮沉降通量的研究.土壤學報,2008,45(2):280-287.
[25] 馬雪華.在杉木林和馬尾松林中與水的養分淋溶作用.生態學報,1989,9(1):15-20.
[28] 魯顯楷,莫江明,李德軍,張煒,方運霆.鼎湖山主要林下層植物光合生理特性對模擬氮沉降的響應.北京林業大學學報,2007,29,1-9.
[29] 馬雪紅,周志春,金國慶,張一.競爭對馬尾松和木荷覓取異質分布養分行為的影響.植物生態學報,2009,33(1):81-88.
[40] 王秀花,陳柳英,馬麗珍,范輝華,葉穗文,周志春.7年生木荷生長和木材基本密度地理遺傳變異及種源選擇.林業科學研究,2011,24(3):307-313.
[41] 劉青華,金國慶,張蕊,儲德裕,秦國峰,周志春.24年生馬尾松生長、形質和木材基本密度的種源變異與種源區劃.林業科學,2009,45:55-61.
[42] 孫成志,謝國恩,李萍.杉木地理種源材性變異及建筑材優良種源評估.林業科學,1993,29:429-436.
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
小讀者(2021年2期)2021-03-29 05:03:48
火花(2019年12期)2019-12-26 01:00:28
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
人大建設(2019年12期)2019-05-21 02:55:32
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
