冬小麥節水栽培群體“穗葉比”及其與產量和水分利用的關系
2013-12-16 08:20:14張永平張英華王志敏
生態學報 2013年12期
關鍵詞:高產
張永平,張英華,黃 琴,王志敏,*
(1.中國農業大學農學與生物技術學院,北京 100193;2.內蒙古農業大學農學院,呼和浩特 010018)
由于水資源日趨緊缺,我國北方冬小麥生產必須走節水高產之路。研究在限水灌溉條件下實現高產與高水分利用效率相結合的理論與技術具有重要意義[1]。建立合理的群體結構既是高產形成的基礎,也是實現高水效的關鍵。然而,對不同品種、在不同生態栽培條件下,獲取高產的群體結構雖然有共同的質量要求,但卻可能有不同的個體形態特征及其數量組成,而不同的株群構型與數量組合,其水分利用效率可能有很大差異,需要深入探索小麥節水高產群體適宜結構指標及其調控途徑[2]。自Mason和Maskell 1928年提出了“源庫”學說后,作物栽培生理理論探討中,常用源、庫關系闡明作物產量形成的規律,探索實現高產的途徑[3-6]。20世紀70年代,日本學者曾提出,可用“粒葉比(單位面積粒數與葉面積之比)”來反映作物產量庫/源比[6-7]。近年來,我國許多學者將粒葉比作為品種光合生產力和“源庫”協調性的指標[8-10]。凌啟鴻等以“粒葉比”為指標分析了稻麥高產栽培途徑,認為粒/葉比是反映群體質量的綜合指標,在最大葉面積指數基本相同條件下,粒葉比越大產量越高[9]。在限水灌溉下對小麥不同基因型的聚類分析也表明[10],單莖葉面積適中,粒葉比高的品種經濟產量最高,粒葉比與經濟產量具有正相關性。然而,粒葉比指標中的粒數是由單位面積穗數和穗粒數兩個因素組成的,提高群體粒/葉比的途徑可以是在一定穗數基礎上增加穗粒數,也可以是在保證一定穗粒數基礎上增加總穗數。在生產實踐中,采取何種途徑應考慮品種和生態條件。在華北地區氣候生態條件下實行節水栽培,前期需要通過適當晚播來控制耗水,而后期又處于干旱和高溫脅迫環境,提高穗粒數或粒重難度較大,穗數對群體庫容的擴大和最終產量具有決定性影響[1],因此,在適宜葉面積基礎上增加穗數是提高粒葉比、進而提高產量的重要策略。同時,增加穗數也增加了群體非葉光合面積,對于后期逆境下光合供源的增加和維持也具有重要作用[2]。相對于“粒葉比”來說,“穗葉比(單位面積穗數與葉面積之比)”可能是節水栽培群體更直接、更為適合的調控指標。為驗證這一想法,明確穗葉比與產量和水分利用效率的關系,本研究基于華北平原冬小麥節水高產栽培要求[1],考察了不同品種、不同水分條件、不同密度條件下群體穗葉比的變化,分析了穗葉比變化與群體光合生產、物質運轉、產量和水分效率等的關系,探討了穗葉比作為節水高產群體調控指標的可能性及其生理學意義,以期為華北地區冬小麥節水高產實踐提供指導。
1 材料與方法
1.1 試驗地與氣象條件
試驗于2001—2003年在中國農業大學吳橋試驗站進行。供試土壤為壤質底粘潮土,土層深厚,地下水位6—9m,土壤肥力中等偏上,兩年度播前測定的耕層養分含量基本相近,平均值為:有機質1.1 g/kg,全氮0.9 g/kg,堿解氮70.6mg/kg,速效磷21.1mg/kg,速效鉀 90.5mg/kg。2001—2002 年度小麥生長期間共降雨 53.5 mm,相當于常年平均降水量(124 mm)的43%,屬于嚴重干旱的年份;2002—2003年度小麥生長期間降水量為112.1 mm,為平水年份。
1.2 試驗材料與試驗設計
1.2.1 田間不同灌水處理試驗
2001/2002年度供試品種為魯麥21,播種前澆底墑水750 m3/hm2,于2001年10月12日播種,基本苗為6×106株/hm2。不澆凍水。試驗設春季不灌水(W0)、春季灌1水(拔節期,W1)、春季灌2水(拔節+開花水,W2)和春季灌4水(起身+孕穗+開花+灌漿水,W4)4個處理,每次灌水定額750 m3/hm2,3次重復,隨機區組排列,共12個試驗小區,小區面積50 m2。
2002/2003年度采用2個品種魯麥21和石家莊8號,播種前澆底墑水750 m3/hm2,于2002年10月13日播種,基本苗為6×106株/hm2。不澆凍水。試驗設春季不灌水(W0)、春季灌2水(拔節+開花,W2)和春季灌4水(起身+孕穗+開花+灌漿,W4)3個處理,每次灌水定額750 m3/hm2,3次重復,隨機區組排列,共計18個試驗小區,小區面積50 m2。
兩個試驗年度均按節水栽培技術施肥標準進行施肥,施肥量為有機肥(雞糞+土雜糞)30 m3/hm2,磷酸二銨300 kg/hm2,硫酸鉀150 kg/hm2,尿素225 kg/hm2,硫酸鋅 15 kg/hm2,均作底肥一次性施入。小區間有隔離帶。其它田間措施按節水高產栽培技術[1]實施。
1.2.2 限水灌溉下不同密度試驗
2001/2002年度供試品種為萊州137,設置不同密度基本苗處理,包括:3.0×106株/hm2(P3)、4.5×106株/hm2(P4.5)、5.25×106株/hm2(P5.25)、6.0×106株/hm2(P6)、6.75×106株/hm2(P6.75)、7.5×106株/hm2(P7.5)6個處理,3次重復,隨機區組排列,小區面積40m2。于2001年10月11日播種,播種前澆底墑水750m3/hm2,春季灌2水(拔節水+開花水,每次灌水定額750m3/hm2),施肥水平及其他管理措施同前述不同灌水試驗。
2002/2003年度供試品種為魯麥21和石家莊8號,分別設置基本苗4.5×106株/hm2(P4.5)、6×106株/hm2(P6)、7.5×106株/hm2(P7.5)3個處理,各處理3次重復,隨機區組排列,小區面積40m2。于2003年10月13日播種,播種前澆底墑水750m3/hm2,春季灌2水(拔節水+開花水,每次灌水定額750m3/hm2),施肥水平及其他管理措施同前述不同灌水試驗。
1.2.3 限水灌溉下品種比較試驗
2001/2002年度選擇穗型(大穗、小穗)、葉型(葉片大小)等不同的基因型品種材料23份,每品種小區面積8.4m2,重復3次。2001年10月12日播種,播前澆底墑水750 m3/hm2,春季拔節期灌1水(750 m3/hm2),施肥水平及其他管理措施同前述不同灌水試驗。
1.3 測定內容與方法
1.3.1 植株綠色器官面積測定
于小麥各生育時期,從每處理小區選取2行、每行50cm典型樣段帶回室內,測量樣段內所有植株各器官綠色面積。葉面積采用長寬系數法,穗面積的測定參照Teare[11]和裘邵峰[12]的方法,分別測穗穎片及芒的表面積,莖鞘面積按圓柱形表面積測定。
1.3.2 光合與呼吸速率測定
群體光合與呼吸測定參照董樹亭[13]同化箱法,同化箱體積為1m3,分別在抽穗、開花和灌漿期測定,每處理測3個重復。每次測定時間為10:00—11:00。
1.3.3 器官干重及貯藏物利用的測定
小麥開花后,每隔5d從不同處理小區取2行、每行50 cm代表性樣段,按葉片、莖鞘、穗穎和籽粒等分器官處理,80℃烘干至恒重,稱量。根據下列公式計算非葉器官(莖鞘和穗穎)貯藏物利用指標:
總的貯藏物質轉運量(g/m2)=器官最大干物重-成熟期器官干物重
總的貯藏物質轉移率(%)=(器官最大干物重-成熟期器官干物重)/器官最大干物重×100
總的貯藏物質貢獻率(%)=(器官最大干物重-成熟期器官干物重)/籽粒產量干重×100
開花前貯藏物質轉運量(g/m2)=開花期器官干物重-成熟期器官干物重
開花前貯藏物質轉移率(%)=(開花期器官干物重-成熟期器官干物重)/開花期器官干物重×100
開花前貯藏物質貢獻率(%)=(開花期器官干物重-成熟期器官干物重)/籽粒產量干重×100
開花后貯藏物質轉運量(g/m2)=總的貯藏物質轉運量-開花前貯藏物質轉運量
開花后貯藏物質貢獻率(%)=(總的貯藏物質轉運量-開花前貯藏物質轉運量)/籽粒產量干重×100
1.3.4 考種與測產
小麥成熟時,分區調查和測定群體穗數、粒數和粒重。在各試驗小區選取2m2×2代表性樣點,單獨收割、脫粒測產。實測籽粒產量按13%籽粒含水量進行校正。同時,測算生物產量、收獲指數等指標。
1.3.5 土壤水分含量與水分利用效率測定
在小麥播前和主要生育時期,分層測定2m土體內的土壤含水量,采用土壤水分平衡法計算不同處理的耗水狀況。總耗水量和水分利用效率按如下公式計算:

1.3.6 穗葉比與粒葉比的測算
在調查群體實際穗數和測定葉面積基礎上,按下式計算穗葉比和粒葉比:

2 結果與分析
2.1 不同處理下小麥群體粒葉比和穗葉比的變化
本文用粒葉比和穗葉比兩個指標來反映小麥群體或個體庫、源關系。由表1、表2可見,兩個年度不同小麥品種開花期的穗葉比均隨著灌水減少、密度增加而顯著增大,即單位葉面積承載的穗數增多。開花期粒葉比也表現出相似的變化趨勢。說明,減少灌水次數明顯限制了葉面積的增大,尤其是上三葉的面積,而在本試驗范圍內增加密度主要是擴大了庫,其結果均使庫容/葉面積相對比值增大。相關分析表明,兩個年度不同灌水和密度處理下,開花期穗葉比與粒葉比均呈極顯著正相關,相關系數分別為0.7243和0.7664(P<0.01)。
在節水條件下,對23個小麥品種開花期穗葉比及相關指標的測定表明(表2),穗葉比、粒葉比、經濟產量、葉及非葉器官面積在品種間均存在較大差異,從變異系數來看,品種間粒葉比的變異系數要大于穗葉比的變異系數。總體來看,經濟產量居前的品種其穗葉比和粒葉比亦表現較高。相關分析表明,在本試驗條件下,穗葉比、粒葉比與經濟產量均呈顯著正相關,而且,穗葉比與粒葉比達到極顯著的正相關(P<0.01)。以上結果說明,在晚播節水栽培下,穗葉比和粒葉比作為衡量庫源關系的指標具有相對一致性。
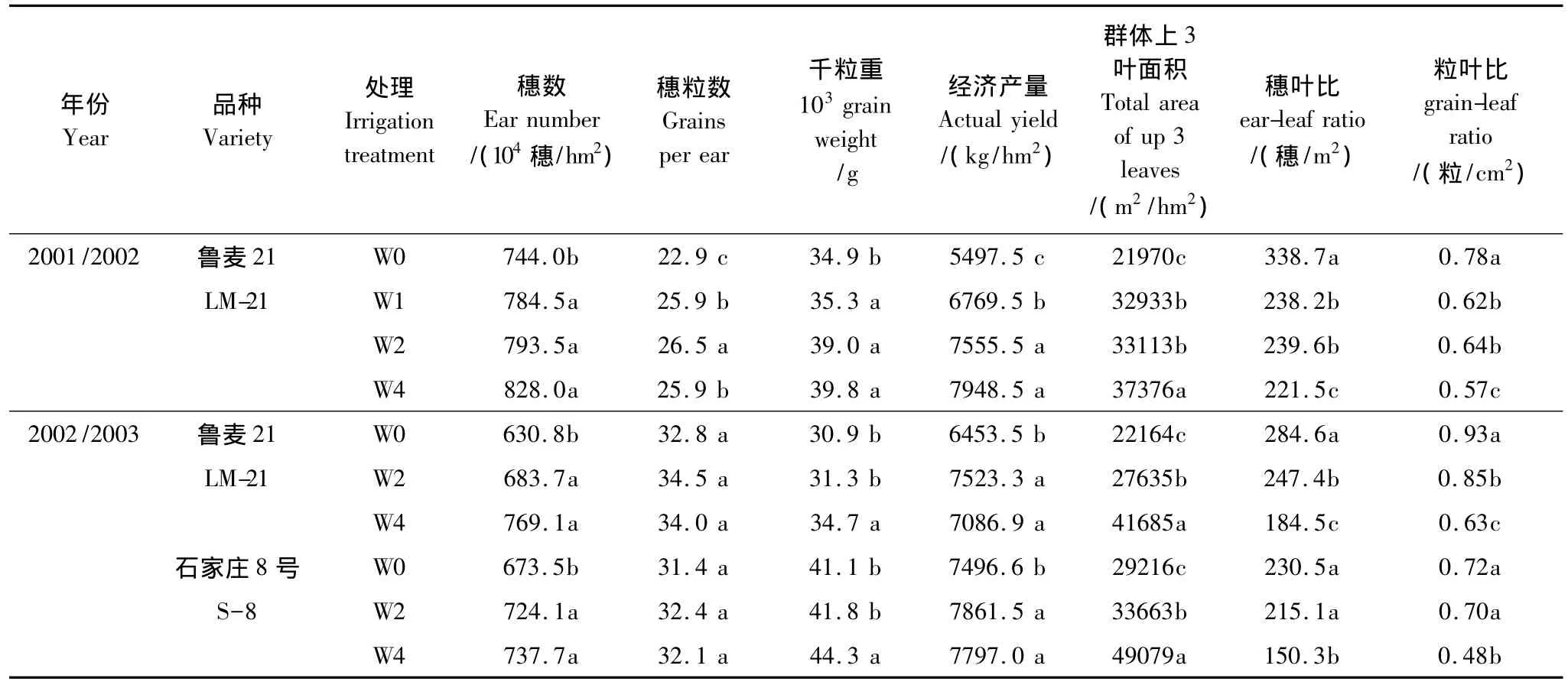
表1 不同灌水處理群體穗葉比的變化Table 1 Ear-leaf area ratio of population in different irrigation treatments
2.2 穗/葉比與群體光合面積的關系
穗葉比為群體穗數與上三葉葉面積的比值,植株個體及群體光合面積的組成與分布必然影響到其比值高低。回歸分析表明(圖1),開花期穗葉比與單莖葉面積、單莖非葉器官面積均呈顯著的線性回歸關系,即隨著單莖葉面積和單莖非葉面積降低,穗葉比明顯增大。從群體角度分析發現(圖2),不同灌水及密度處理下,穗葉比與葉面積指數(LAI)均呈極顯著負相關,而與非葉面積指數相關表現不盡一致,不同灌水處理下穗葉比與非葉面積指數(NAI)呈極顯著負相關,不同密度處理下則表現為正相關關系,但未達到顯著水平。也即,通過減少灌水次數或在有限灌溉下適當增加密度提高了穗葉比,主要是群體葉面積相對降低所致,而非葉面積并未顯著減少,從而使群體內非葉綠色面積相對比例增加,這有利于開花后土壤水分虧缺條件下保持較高的群體光合活性。

表2 不同密度處理群體穗葉比的變化Table 2 Ear-leaf ratio of population in different density treatments

圖1 群體穗葉比與單莖光合面積指標的關系Fig.1 Relationship between ear-leaf ratio and photosynthetic area indices per stem

表3 節水條件下不同冬小麥品種穗葉比及其相關指標的變化(開花期)Table 3 Ear-leaf area ratio of population and related indices of different wheat varieties under water-saving irrigation treatment(anthesis)
2.3 穗/葉比與群體光合速率和呼吸速率的關系
對2年不同密度處理開花期的群體光合與呼吸速率進行了測定。回歸分析結果表明(圖3),開花期穗葉比與群體光合、呼吸速率均呈二次拋物線關系,相關系數檢驗分別達極顯著和顯著水平,即在一定范圍內隨著穗葉比的增大,群體光合速率和呼吸速率都相應增加。2002/2003年度,隨著穗葉比增大群體光合的增加幅度略大于群體呼吸的增幅;2001/2002年度,群體光合的增加幅度明顯大于群體呼吸的增幅,導致凈光合物質生產增多。對回歸方程求極值可知,兩個年度群體光合速率達到最大值時的穗葉比分別為270穗/m2和255穗/m2。說明,在冬小麥節水栽培中,通過增加種植密度增加穗數,使開花期穗葉比達到適宜較高值,可改善小麥群體內的光照狀況,提高產量形成期群體的光合物質生產能力。
2.4 穗/葉比與貯藏物質利用的關系
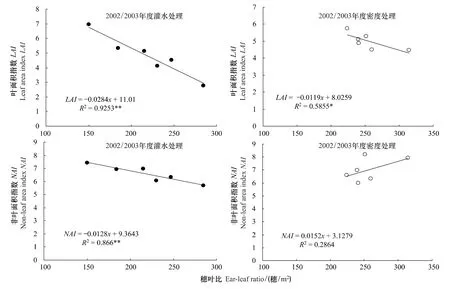
圖2 群體穗葉比與群體光合面積指標的關系Fig.2 Relationship between ear-leaf ratio and photosynthetic area indices in canopy
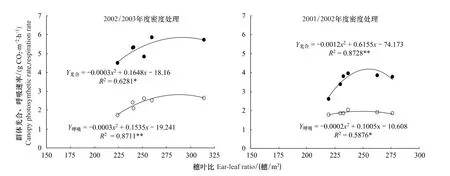
圖3 開花期穗葉比與群體光合、呼吸速率的關系Fig.3 Relationship between ear-leaf ratio and photosynthetic rate and respiration rate of population at anthesis
小麥籽粒產量一方面來自于開花后光合生產并直接運輸到籽粒中的物質,另一方面來自于花前和花后貯藏于植株體而后再運轉到籽粒中的物質,非葉器官莖鞘和穗穎是主要的暫時貯藏器官。對不同灌水處理、不同密度處理非葉器官(莖鞘+穗穎)貯藏物質轉運的分析表明(表4),隨著灌水次數減少或種植密度增加,非葉器官貯藏物質轉運量、轉移率及對籽粒的貢獻率均明顯增加。如在春澆2水的節水栽培條件下,非葉器官平均總的貯藏物質轉運量、轉移率和對籽粒貢獻率分別為357.8g/m2、40.3%和46.5%,其中花前對應數值為185.8 g/m2、26.3%和 24.2%,花后貯藏物轉運量和貢獻率分別為 172.0 g/m2和 22.2%;葉片平均總的貯藏物質轉運量、轉移率和對籽粒貢獻率分別為115.0 g/m2、48.4%和15.1%,其中花前對應數值為71.2 g/m2、36.9%和9.3%,花后轉運量和貢獻率分別為43.8 g/m2和5.7%。非葉器官貯藏物質轉移率低于葉片,但轉運量和對籽粒貢獻率顯著高于葉片。
統計分析表明,開花期穗葉比與非葉器官貯藏物質轉移率呈顯著線性相關(圖4),與貯藏物對產量的貢獻率也存在相關性,但相關不顯著,與貯藏物轉運量無明顯的線性關系。說明,增加穗葉比可促進非葉器官貯藏物質的轉移和經濟利用。
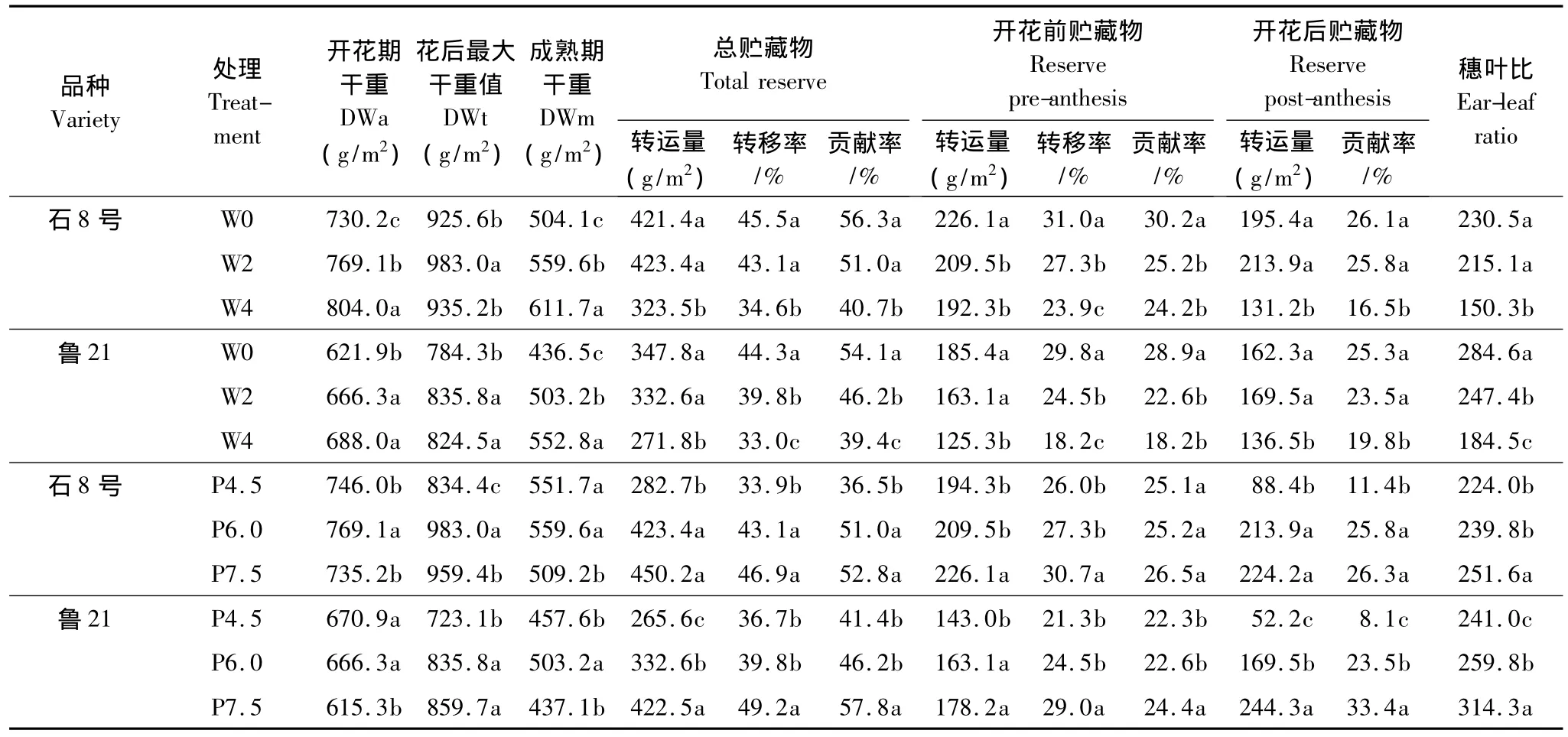
表4 非葉器官物質再分配及其對籽粒的貢獻(2002/2003)Table 4 Reallocation of dry weight(DW)and contribution to grains of non-leaf organs after anthesis

圖4 穗葉比與非葉器官物質轉運的關系Fig.4 Relationship between ear-leaf ratio and remobilization of reserve in non-leaf organs
2.5 穗/葉比與產量和收獲指數的關系
由圖5可見,不同處理下群體穗葉比與籽粒產量均符合二次拋物線曲線變化,相關系數檢驗達到顯著或極顯著水平。對回歸方程分別求導并綜合分析得出,經濟產量達到最大值的適宜穗葉比值為230—270穗/m2。
不同密度處理及不同品種處理下,開花期群體穗葉比與收獲指數均呈極顯著正相關,即在節水條件下,選擇高穗葉比品種或通過增加密度增大穗葉比,均可顯著提高收獲指數。
2.6 穗/葉比與耗水量和水分利用效率的關系
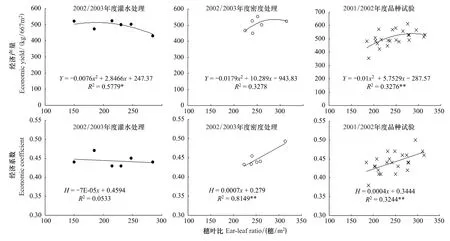
圖5 穗葉比與經濟產量和收獲指數的關系Fig.5 Relationship between ear-leaf ratio and yield,harvest index
由圖6可見,不同灌水處理下穗葉比與總耗水量呈極顯著的負相關關系,而不同密度處理下穗葉比與總耗水量的相關關系不明顯,但仍可看出較高的穗葉比易獲得較低的耗水量;不同處理下穗葉比與水分利用效率(WUE)均呈拋物線關系,即在一定范圍內,隨著穗葉比的增加,冬小麥生育期總耗水量明顯減少,而水分利用效率則顯著提高。對回歸方程分別求解得出,獲得最高WUE的穗葉比值為250穗/m2左右。
3 討論
源庫學說認為,作物產量既取決于源的光合物質生產能力,又取決于庫的大小和庫強度,較大的庫容和庫強度可促進光合物質源的生產與運轉[14-15]。高產栽培需要建立合理的群體結構,以協調源-庫關系。小麥高產的途徑可以走“小群體、壯個體”途徑,也可以走“大群體、小個體”途徑,關鍵是因地制宜,通過密度與株型的合理組配,擴庫強源,使源、庫在高水平上協調[6]。華北地區冬小麥實施節水栽培,需要適當推遲播期以減少前期水分消耗,而在晚播和有限的水分供給下,增加穗粒數的潛力有限,擴大群體庫容量的重點是增加穗數,應在確保足夠穗數的基礎上穩定穗粒數。晚播增穗的可靠方式是增加基本苗,因此必須走大群體高產之路。在大群體條件下,若葉面積過分擴大,則會使中后期群體惡化,葉片遮光,光合生產率下降,而無效呼吸、無效耗水增加,源庫不能協調。因此必須控制葉面積,在適宜的葉面積范圍之內增穗擴庫。許多研究表明,粒葉比是反映群體源庫是否協調的綜合指標[8-10]。本研究表明,在限水灌溉條件下,小麥群體穗葉比與粒葉比具有顯著的正相關性。由于穗數和葉面積均在孕穗前基本決定且易觀察,因此穗葉比可以作為小麥節水高產栽培群體源庫關系的適宜調控指標。
小麥的光合源器官不僅包括葉片,還包括穗、莖、鞘等非葉綠色器官,因此,穗/葉比既是庫/源比指標,也是非葉源/葉源比指標。在小麥抽穗后,直立分布于群體上層的穗、穗下節間和旗葉鞘這些非葉器官不僅具有光、CO2截獲的空間優勢,而且具有逆境下的光合生理優勢。近年來的研究表明[16-21]:小麥穗部器官具有旱生型結構;水勢相對穩定;具有較高的PEP羧化酶活性,并可能存在類C4型光合代謝機制;籽粒呼吸釋放的CO2能被穗苞片(護穎、內外稃)重新固定。在干旱和高溫脅迫下,葉片功能易早衰,而穗等非葉器官光合具有較強的穩定性,對產量具有較大貢獻。穗等非葉綠色器官與葉片相比,具有逆境下的高光合潛力和相對節水優勢。因此,在適宜葉面積基礎上適當增加穗數,既可增加群體非葉綠色面積,從而擴大群體光合面積,也可增強群體光合耐逆性能。從本研究結果看,隨著灌水減少或密度增加,開花和灌漿期的穗/葉比均相應增大,說明控制灌水主要抑制了葉源的增大,而在限水灌溉下適當增大密度則相對增大了穗庫容。回歸分析表明,隨著穗葉比增大,群體葉面積指數(LAI)雖明顯降低,但非葉面積指數(NAI)卻維持相對較高水平;而且灌漿期群體光合速率顯著增加。同時,穗/葉比與營養器官貯藏物質轉運率呈正相關關系,這說明庫/源比的提高促進了貯藏物質轉運,增加了花前貯藏物對籽粒產量的貢獻。因此,穗葉比可作為節水栽培群體光合系統質量的調控指標。在小麥節水栽培體系中,通過限制供水來控制葉片面積并提高葉片質量,通過增加密度來提高群體中非葉綠色器官的比例,增加穗/葉比,可以充分發揮非葉綠色器官的抗旱節水作用和光合潛力,并促進貯藏物質運轉,有利于維持或增加后期產量形成的物質供源。

圖6 穗葉比與耗水量和WUE的關系Fig.6 Relationship between ear-leaf ratio and water consumption,WUE
葉片是光合器官,也是主要的蒸騰器官。葉面積較大,而經濟器官(穗)比例較小的群體,耗水多,水分利用不經濟[10]。在有限供水條件下,適當控制葉面積,有利于提高水分利用效率[22]。本研究發現,群體穗葉比與經濟產量的關系符合二次拋物線曲線變化,穗葉比與收獲指數則呈極顯著正相關。從水分利用方面看,穗葉比增加,冬小麥生育期總耗水量明顯減少,而水分利用效率則顯著提高。因此,穩定適宜葉面積、適當增加穗/葉比,創造適宜穗葉比的群體結構,可以實現高產與高水效的結合。綜合分析認為,冬小麥節水高產栽培群體上3葉適宜葉面積指數約為(3±0.2),適宜的穗/葉比值為230—270穗/m2,即每平方米高效葉面積可承載的穗數最適值在230—270穗之間。
[1] Lan L W,Zhou D X.Studies on Water-Saving and High-Yielding of Winter Wheat.Beijing:China Agricultural University Press,1995.
[2] Zhang Y P,Wang Z M,Wang P,Zhao M.Canopy photosynthetic characteristics of population of winter wheat in water-saving and high-yielding cultivation.Scientia Agricultura Sinica,2003,36(10):1143-1149.
[3] Cao X Z,Zhu Q S.Study on characteristics of the relationship between source and sink in rice varieties and their classification.Acta Agronomica Sinica,1987,13(4):265-272.
[4] Guo W S,Feng C N,Yan L L,Peng Y X,Zhu X K,Zong A G.Analysis on source-sink relationship after anthesis in wheat.Acta Agronomica Sinica,1995,21(3):334-340.
[5] Koshkin E I,Tararina V V.Yield and source/sink relations of spring wheat cultivars.Field Crop Research,1989,22(4):297-306.
[6] Wang Z M,Fang B T.A review on theoretical models and development of yield analysis in crop production system.Journal of China Agricultural University,2009,14(1):1-7.
[7] Welbank P J,Witts K J,Thorne G N.Effect of radiation and temperature on efficiency of cereal leaves during grain growth.Annals of Botany,1968,32(1):79-95.
[8] Yan X P,Zhao X L,Yen M Q.A study on the grain-leaf ratio of wheat.Journal of Henan Agricultural University,1989,23(2):103-108.
[9] Ling Q H,Yang J C.Studies on“grain-leaf ratio”of population and cultural approaches of high yield in rice plants.Scientia Agricultura Sinica,1986,(3):1-8.
[10] Zhang Y P,Wang Z M,Wu Y C.Analysis on photosynthetic characteristics of wheat variety under the water-saving and high-yielding cultivation condition.Acta Agriculturae Boreali-Sinica,2004,19(3):47-54.
[11] Teare I D,Law A G,Simmons G F.Stomatal frequency and distribution on the inflorescence of Triticum aestivum L.Canadian Journal of Plant Science,1992,52(1):89-94.
[12] Qiu S F,Zai L Y.The estimation for surface area of spike and awn of the common wheat.Acta Agronomica Sinica,1985,11(2):138-144.
[13] Dong S T.Studies on the relationship between canopy apparent photosynthesis and grain yield in high-yielding winter wheat.Acta Agronomica Sinica,1991,17(6):461-469.
[14] Blade S F,Baker R J.Kernel weight response to source-sink changes in spring wheat.Crop Science,1991,31(5):1117-1120.
[15] Plaut Z,Mayoral M L,Reinhold L.Effect of altered sink:source ratio on photosynthetic metabolism of source leaves.Plant Physiology,1987,85(3):786-791.
[16] Morgan J M.Osmotic adjustment in the spikelets and leaves of wheat.Journal of Experimental Botany,1980,31(2):655-665.
[17] Wang Z M,Wei A L.Photosynthetic characteristics of non-leaf organs of winter wheat cultivars differing in ear type and their relationship with grain mass per ear.Photosynthetica,2001,39(2):239-244.
[18] Araus J L,Brown H R,Febrero A,Bort J,Serret M D.Ear photosynthesis,carbon isotope discrimination and the contribution of respiratory CO2to differences in grain mass in durum wheat.Plant,Cell and Environment,1993,16(4):383-392.
[19] Wei A L,Wang Z M,Zhai Z X,Gong Y S.Effect of soil drought on C4photosynthesis enzyme activities of flag leaf and ear in wheat.Scientia Agricultura Sinica,2003,36(5):508-512.
[20] Xu X L,Zhang Y H,Wang Z M.Effect of heat stress during grain filling on phosphoenolpyruvate carboxylase and ribulose-1,5-bisphosphate carboxylase/oxygenase activities of various green organs in winter wheat.Photosynthetica,2004,42(2):317-320.
[21] Xu X L,Zhang Y H,Wang Z M.Effect of heat stress on photosynthetic characteristics of different green organs in winter wheat during grain-filling.Acta Botanica Sinica,2001,43(6):571-577.
[22] Richards R A.Manipulation of leaf area and its effect on grain yield in droughted wheat.Australian Journal of Agricultural Research,1983,34(1):23-31.
參考文獻:
[1] 蘭林旺,周殿璽.小麥節水高產研究.北京:中國農業大學出版社,1995.
[2] 張永平,王志敏,王璞,趙明.冬小麥節水高產栽培群體光合特征.中國農業科學,2003,36(10):1143-1149.
[3] 曹顯祖,朱慶森.水稻品種的庫源特征及其類型劃分的研究.作物學報,1987,13(4):265-272.
[4] 郭文善,封超年,嚴六零,彭永欣,朱新開,宗愛國.小麥開花后源庫關系分析.作物學報,1995,21(3):334-340.
[6] 王志敏,方保停.論作物生產系統產量分析的理論模式及其發展.中國農業大學學報,2009,14(1):1-7.
[8] 閻新甫,趙獻林,任明全.小麥粒葉比的研究.河南農業大學學報,1989,23(2):103-108.
[9] 凌啟鴻,楊建昌.水稻群體“粒葉比”與高產栽培途徑的研究.中國農業科學,1986,(3):1-8.
[10] 張永平,王志敏,吳永成.節水高產栽培小麥品種光合性狀分析.華北農學報,2004,19(3):47-54.
[12] 裘昭峰,翟立業.小麥穗和芒表面積的估測.作物學報,1985,11(2):138-144.
[13] 董樹亭.高產冬小麥群體光合能力與產量關系的研究.作物學報,1991,17(6):461-469.
[19] 魏愛麗,王志敏,翟志席,龔元石.開花后水分脅迫對小麥不同器官C4光合酶活性的影響.中國農業科學,2003,36(5):508-512.
猜你喜歡
現代畜牧科技(2021年6期)2021-07-16 05:50:06
河北農機(2020年10期)2020-12-14 03:13:30
今日農業(2019年15期)2019-01-03 12:11:33
新農業(2017年22期)2018-01-03 05:46:00
中國農業文摘-農業工程(2016年5期)2016-04-12 05:38:15
現代農業(2016年5期)2016-02-28 18:42:33
現代農業(2016年5期)2016-02-28 18:42:33
現代農業(2016年4期)2016-02-28 18:42:19
現代農業(2015年5期)2015-02-28 18:40:54
作物研究(2014年6期)2014-03-01 03:39:10
