基于MAXENT模型的賀蘭山巖羊生境適宜性評價
2013-12-20 05:59:56劉振生滕麗微王曉勤孔芳毅
生態學報 2013年22期
劉振生,高 惠, 滕麗微,*,蘇 云,王曉勤,孔芳毅
(1.東北林業大學野生動物資源學院,哈爾濱 150040;2.國家林業局野生動物保護學重點開放實驗室,哈爾濱 150040;3.內蒙古賀蘭山國家級自然保護區管理局,巴彥浩特 750300)
生境為野生動物提供了生存、繁衍及種群發展所必須的資源[1],生境適宜度是指生境能支持一個特定物種的潛在能力[2],是影響野生動物生存和發展的重要因素,對物種進行生境評價是對野生動物進行有效保護和管理的第一步,并且能夠為相關部門制定有效的物種保護對策提供科學依據。
巖羊(Pseudois nayaur)屬偶蹄目、牛科、羊亞科、巖羊屬,是青藏高原的特有種,也是青藏高原分布最廣的有蹄類之一,被世界自然保護聯盟(IUCN)收錄為低危種[3],在我國被列為國家Ⅱ級重點保護野生動物[4]。國內主要分布于西藏、云南、四川、內蒙古、新疆、甘肅、青海、寧夏和陜西等省區[5]。目前,專家學者已對其行為生態學[6-7]、食性[8]、種群動態[9-10]和家域[11]等方面做了大量研究。劉振生等采用直接觀察法對該區域巖羊冬春季的取食生境選擇進行了研究[12]。劉振生等對賀蘭山地區巖羊的種群數量及分布做了調查和報道,2003年巖羊種群數量約為10 000只,東坡是巖羊的主要分布區[13]。劉振生等利用距離取樣法對賀蘭山地區不同季節巖羊的種群密度進行了估測[14]。在巖羊的生境評價方面,尚缺乏相關的報道。
近年來,隨著3S(GIS,RS,GPS)技術的發展,從大尺度上對野生動物進行生境研究成為熱點[15-16]。國內外的眾多學者利用機理模型、回歸模型和生態位模型結合3S技術來對目標物種進行生境適宜性評價[17-19]。最大熵模型是生態位模型的一種,近年來已廣泛應用于保護動物的生境分布預測。李明陽等基于WEB數據庫和MAXENT模型,對白頭葉猴的潛在生境進行評價研究[20]。齊增湘等利用MAXENT模型,進行秦嶺山系黑熊潛在生境預測[21]。徐衛華等采用MAXENT模型,對秦嶺地區的川金絲猴的生境進行評價[22]。本文應用GIS技術和MAXENT模型對賀蘭山巖羊生境適宜性進行評價,分析影響巖羊生存的主要生境因子并模擬其空間分布區域,以期為賀蘭山國家級自然保護區的合理規劃及巖羊的保護提供科學依據。
1 材料與方法
1.1 研究地區概況
賀蘭山位于銀川平原與阿拉善高原之間,地理坐標為 N38°21'—39°22',E105°49'—106°42',海拔一般是2 000—3 000 m,由內蒙古賀蘭山國家級自然保護區和寧夏賀蘭山國家級自然保護區兩部分組成,是巖羊的一個重要的集中分布區。賀蘭山具有典型的大陸性氣候特征,地處荒漠與半荒漠地帶,主要特點是干旱、高寒。年平均氣溫為-0.9℃,平均年降水量在200—400 mm之間[23]。冬季長達5個月,降雪較少。賀蘭山是典型的溫帶山地森林系統,其植被垂直分布明顯:山地草原帶(1400—1600 m),山地疏林草原帶(1600—2000 m),山地針葉林帶(1900—3000 m),亞高山灌叢和草甸帶(3000—3556 m)[24]。
1.2 模型介紹
最大熵原理(MAXENT)起源于信息科學,最早由Jaynes提出,在許多學科中有廣泛應用,近年來引起了生態學家們的關注[25]。MAXENT是一種基于生態位原理的模型,根據物種“出現點”的環境變量特征得出約束條件,探尋此約束條件下最大熵的可能分布,以此來預測目標物種在研究地區的生境分布[26]。2004年,Phillips等人開發了MAXENT軟件,近來廣泛用于物種生境適宜區的預測和評價,表現出了良好的預測能力[27]。該模型提供了自檢驗功能,可以自動生成ROC曲線進行模型的模擬預測自檢,且在對動物生境進行評價與預測時,只需動物“出現點”的數據,并且具有較高的精度[28-30]。在巖羊“未出現點”的數據難以準確采集的實際情況下,MAXENT模型相比其他模型具有更大的優勢。
1.3 數據來源及預處理
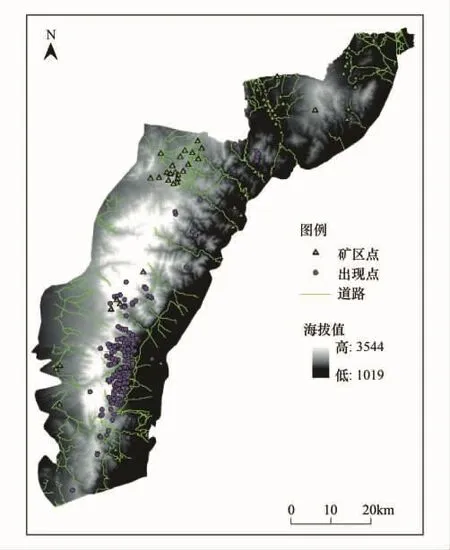
圖1 賀蘭山概況及樣點分布圖Fig.1 The overview of Helan Mountain and distribution of sample points
應用MAXENT模型對巖羊進行生境適宜性評價需要兩方面數據:一是巖羊的現實地理分布點數據,二是賀蘭山地區的環境變量數據。其中巖羊分布點數據來源于野外調查以及賀蘭山保護區各管理站的野外監測,自2010年至2011年,根據賀蘭山各個溝段的實際情況,分別在寧夏賀蘭山國家級自然保護區的馬蓮口、蘇峪口、大水溝、紅果子4個管理站和內蒙古賀蘭山國家級自然保護區的腰壩、哈拉烏、水磨溝、古拉本4個管理站的25條溝系,按春、夏、秋、冬4個季節以步行的方法對巖羊進行調查。這些溝系分布于不同的海拔高度,覆蓋了賀蘭山巖羊棲息的全部生境類型,保證了分布點數據的代表性和準確性。用GPS對所觀察到的巖羊實體或巖羊利用痕跡的位置進行定點記錄,在賀蘭山與巖羊同域分布的有蹄類還有馬麝(Moschus chrysogaster)和馬鹿(Cervus elaphus),可根據體形大小和形態特征對它們進行區分,共得到了317個巖羊出現的GPS坐標點。環境變量數據包括:①地形因子數據,包括海拔、坡度和坡向,由中國科學院科學數據庫30 m分辨率的DEM(數字高程圖)計算提取得到;②植被因子數據,利用ERDAS 9.1遙感影像處理軟件,采用監督分類方法對2007年賀蘭山的TM影像進行分類處理,將其分為針葉林、闊葉林、灌叢、草甸、草地、荒漠和水體等7種類型;③其他數據,包括護林點、礦區、道路及水源因子,從賀蘭山1∶50000的矢量化地圖中提取獲得,利用ARCGIS的空間分析求算到各因子的距離柵格圖層。
以ARCGIS為平臺,去除邊界之外的GPS坐標點,將所有環境變量的圖層統一邊界,坐標系統統一為WGS-1984-UTM-Zone-48N,柵格大小統一為30×30 m,并轉化成MAXENT軟件所要求的ASCII格式的文件。
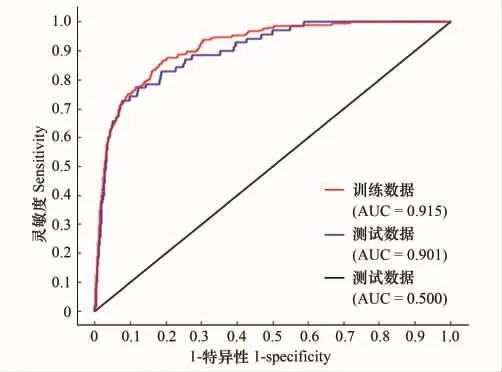
圖2 賀蘭山巖羊生境評價結果的ROC曲線驗證Fig.2 ROC curve verification of blue sheep in the Helan Mountains,China
1.4 模擬方法
將巖羊現實分布點的數據和環境變量數據導入MAXENT 3.3中,隨機選取75%的巖羊分布點用于建立模型,其余25%的巖羊分布點用于模型驗證,選擇利用Jackknife來檢測變量的重要性,并對各生境因子進行敏感性分析,其它參數均為模型的默認值,結果以Logistic格式輸出。模型的預測結果利用受試者工作特征曲線下的面積值進行檢驗,其評價標準為:曲線下的面積的值在 0.5—0.6,不及格;0.6—0.7,較差;0.7—0.8,一般;0.8—0.9,良好;0.9—1.0,優秀[31]。將模型輸出結果放入 ArcGIS 中進行進一步分析,按照專家經驗法[32],對模型預測的賀蘭山巖羊分布圖進行重新分類。
2 結果與分析
2.1 MAXENT預測結果檢測
ROC曲線評價結果為:訓練集與驗證集的AUC值分別為0.915和0.901,表明MAXENT模型的預測結果達到優秀水平(圖1)。
2.2 巖羊分布與環境因子的關系
Jackknife檢驗結果表明:坡度、距礦區距離、海拔、距道路距離、距水源距離是影響巖羊生境分布的主要環境因子。環境變量對MAXENT模型的貢獻率說明:距礦區距離(35.5%)、坡度(27.8%)、海拔(12.6%)、距道路距離(10.2%)和距水源距離(7.3%)5個環境變量的累積貢獻率達86%,對巖羊的生境選擇具有重要影響,礦區和道路是對巖羊影響最大的2個人為干擾因子;針葉林、灌木、水體和荒漠等因子的貢獻率為零,對巖羊的生境選擇沒有影響;其余因子的影響率較小,貢獻值由大到小排序依次為坡向、護林點、草地、草甸和闊葉林,坡向對巖羊的生境選擇具有一定影響,巖羊的主要食物為草本科植物和灰榆,灰榆主要分布于闊葉林中,因此草地、草甸和闊葉林對巖羊的選擇具有影響。各生境因子的敏感性分析表明,隨著坡度、距礦區的距離、距道路的距離的增大,生境適宜度逐漸增加,表明巖羊偏愛崎嶇陡峭地帶,躲避存在人為干擾的區域,在道路分析中,距其約100 m的范圍內干擾值最大,超過100 m后,道路對巖羊的干擾逐漸減少;隨著距水源距離的增加,生境適宜度逐漸減小;適宜度在1500—2300 m之間隨著海拔增加而增大,超過2 300 m之后隨著海拔的增加而逐漸減小。
2.3 賀蘭山巖羊生境適宜性分布
利用MAXENT模型對賀蘭山區域巖羊的生境適宜性進行評價,將模型輸出的ASCII文件導入到ArcGIS 10中,轉換成浮點型柵格數據,按照專家經驗法將生境適宜圖重新分為3個適宜等級:0.00—0.12為不適宜等級,0.12—0.40為較適宜等級,0.40—1.00為最適宜等級,最終得到賀蘭山地區巖羊的生境適宜性分布圖(圖2)。
從圖中可以看出,賀蘭山巖羊的適宜生境主要分布于賀蘭山東坡(寧夏賀蘭山國家級自然保護區)西南部及西坡(內蒙古賀蘭山國家級自然保護區)的山地疏林草原帶。其中最適宜生境的面積為262.3 km2;較適宜生境面積為744.6 km2,總的適宜性生境面積占研究區域面積的28%;不適宜生境面積為2649.1 km2。
3 討論
生境評價模型目前可以分為3種類型:機理模型、回歸模型和生態位模型。機理模型并不需要物種分布點數據,而是根據生境因子對物種分布的影響建立相應的評價準則,進而模擬得到物種的適宜性生境,但因其沒有考慮生境的可達性,且在對因子劃分等級及確定權重上存在主觀性,因此具有一定局限性。相比回歸模型,生態位模型只需動物“出現點”的數據而不需要“未出現點”的數據,根據在賀蘭山野外調查的實際經驗,發現在某個地點未發現巖羊的蹤跡并不代表巖羊未曾在此出現過,因此所記錄的“未出現點”的數據并非十分準確,相比較而言,利用生態位模型可信度更高。本次研究利用MAXENT模型對賀蘭山巖羊生境適宜性進行評價,所取得的結果較為理想。
巖羊“出現點”數據的收集方法是對發現巖羊實體或者巖羊利用痕跡的位置用GPS進行定點記錄,但是由于賀蘭山山勢起伏較大,多陡坡和斷崖等地形,影響了一些出現點的可達性,部分出現點數據不能精確記錄,如巖羊行走的險峻山崖,無法進行準確定點。實際的野外調查中,記錄者在能夠到達的最接近目標地位置進行定點記錄,以對評估結果造成的影響降到最小。此外,道路數據是從賀蘭山1∶50000的矢量化地圖中提取獲得,道路對巖羊影響力的大小與其車流量和寬度有關,而每條道路的建路時間以及車流量等具體信息并不明確,在數據提取過程中,忽略了山間小路這些干擾性小的數據信息,以減少誤差。對于采集數據點達到精確以及提高道路信息準確度的方法仍需進一步的研究改進,以期取得更準確的評價結果。
已有研究表明巖羊偏愛靠近裸巖及懸崖的生境[33],選擇山勢陡峭的山地疏林草原帶,避免選擇山地針葉林帶[34]。模型分析結果表明,自然生境因子中貢獻率最大的為坡度和海拔。隨著坡度的增大,地勢陡峭度增加,生境適宜性逐步提高。海拔高度決定了賀蘭山植被的分布,在1 500—2 300 m海拔內,主要為山地疏林草原帶,生境適宜性逐步增加,其后隨著海拔增高,進入山地針葉林帶,生境適宜性逐漸降低,這些分析結果與之前的研究相符。模型預測結果顯示賀蘭山東坡(寧夏賀蘭山國家級自然保護區)的巖羊適宜分布區的面積要遠大于西坡(內蒙古賀蘭山國家級保護區),這主要是因為東坡山勢陡峻狹短,溝谷深切,地面較破碎,崎嶇度較大,且有大面積的山地疏林草原帶,而西坡的坡面緩長,山地針葉林帶較多[13],因此相比較西坡,東坡具有更廣闊的巖羊適宜生境。
賀蘭山是巖羊的一個重要集中分布區域,也是巖羊分布的最北緣[35]。自20世紀80年代保護區建立以來,保護和管理力度逐年加強,采取了禁止和限制家畜放牧等一系列保護措施[36],使賀蘭山植被得以恢復,環境承載力得到提高。從本研究結果來看,目前最重要的2個人為干擾因素是礦區和道路。針對礦業開采對巖羊造成的影響,保護區應采取相應措施,嚴格控制巖羊生境區內礦區的范圍和數量,使其對巖羊的干擾降到最低。同時應該加強賀蘭山地區道路的規劃和管理,新建道路的選線應盡量避開巖羊頻繁出沒的地段。此外,在對賀蘭山開發建設的同時,應加強對周圍居民的宣傳教育,使人們充分認識到巖羊作為賀蘭山的優勢種具有重要的意義和價值,提高居民的野生動物保護意識,以期對巖羊種群的穩步發展有一定的積極作用。

圖3 賀蘭山巖羊生境適宜性分布Fig.3 The distribution of habitat suitability of blue sheep in Helan Mountains,China
[1]Wang J L,Chen Y.Applications of 3Stechnology in wildlife habitat researches.Geography and Geo-Information Science,2004,20(6):44-47.
[2]Wang X L.Landscape Dynamics and Suitability Assessment of Przewalski's Gazelle Habitat[D].Beijing:Chinese Academy of Forestry,2004.
[3]Baillie J,Groombridge B.IUCN red list of threatened animals.Gland,Switzerland:World Conservation Union,1996.
[4]Liu Z S.Ecology of Blue Sheep(Pseudois nayaur)in Helan Mountains,China[D].Shanghai:East China Normal University,2006.
[5]Sheng H L.Handbook of Fur-bearing Animal.Shanghai:Shanghai Lexicographical Publishing House,1994:218-219.
[6]Oli M K,Rogers M E.Seasonal Patten in group size and population composition of blue sheep in Manang,Nepal.Journal of Wildlife Management,1996,60(4):797-801.
[7]Liu G K,Zhou C Q,Yang Z S,Long S,Pan L,Wang W K,You Y Q,Zeng G W,Geng SS.Diurnal activity rhythm and time budgets of the dwarf blue sheep(Pseudois schaeferi)in Zhubalong Nature Reserve.Acta Ecologica Sinica,2011,31(4):972-981.
[8]Chang Y,Liu Z S,Zhang M M,Li Z G,Hu T H.Summer diets of sympatric blue sheep(Pseudois nayaur)and red deer(Cervus elaphus alxaicus)in the Helan Mountains,China.Acta Ecologica Sinica,2010,30(6):1486-1493.
[9]Wilson P.Ecology and habitat utilization of blue sheep Pseudois nayaur in Nepal.Biological Conservation,1981,21(1):55-74.
[10]Wang X M,Liu Z X,Xu H F,Li M,Li Y G.The blue sheep population ecology and its conservation in Helan Mountain,China.Biodiversity Science,1998,6(1):1-5.
[11]Harris R B,Miller D J.Overlap in summer habitats and diets of Tibetan Plateau ungulates.Mammalia,1995,59(2):197-212.
[12]Liu Z S,Wang X M,Li ZG,Cui D Y,Li X Q.Comparison of seasonal feeding habitats by blue sheep(Pseudois nayaur)during winter and spring in Helan Mountain,China.Zoological Research,2005,26(6):580-589.
[13]Liu Z S,Wang X M,Li Z G,Zhai H,Hu T H.Distribution and abundance of blue sheep in Helan Mountains,China.Chinese Journal of Zoology,2007,42(3):1-8.
[14]Liu Z S,Wang X M,Teng L W,Cui D Y,Li X Q.Estimating seasonal density of blue sheep(Pseudois nayaur)in the Helan Mountain region using distance sampling methods,Ecological Research,2008,23:393-400.
[15]Xu W H,Ouyang Z Y,Li Y,Liu J G.Habitat assessment on giant panda's in Qinling Mountain Range based on RS and GIS.Remote Sensing Technology and Application,2006,21(3):239-242.
[16]Ward D F.Modelling the potential geographic distribution of invasive ant species in New Zealand.Biological Invasions,2007,9(6):723-735.
[17]Steven JP,Miroslav D,Robert E S.A Maximum Entropy Approach to Species Distribution Modelling.Banff,Alberta:Proceedings of the Twenty-First International Conference on Machine Learning,2004:655-662.
[18]Li H M,Han H X,Xue D Y.Prediction of potential geographic distribution areas for the pine bark scale,Matsucoccus matsumurae(Kuwana)(Homoptera:Margarodidae)in China using GARP modelling system.Acta Entomologica Sinica.2005,48(1):95-100.
[19]Brotons L,Thuiller W,Miguel B A,Alexandre H H.Presence-absence versus presence-only modelling methods for predicting bird habitat suitability.Ecography,2004,27(4):437-448.
[20]Li M Y,Xi Q,Xu H G.Evaluation of potential habitat for white-headed langur based on WEB dataset and Maxent model.Forest Resources Management,2010,(4):31-36.
[21]Qi Z X,Xu W H,Xing X Y,Ouyang Z Y,Zheng H,Gan D X.Assessment of potential habitat for Ursusthibetanus in the Qingling Mountains,Biodiversity Science,2011,19(3):343-352.
[22]Xu WH,Luo C.Application of MAXENT model in Rhinopithecusroxllanae habitat assessment in Qinling Mountain.Forest Engineering,2010,26(2):1-4.
[23]Liu Z S,Li X Q,Wang X M,Li Z G,Hu T H,Zhai H.Seasonal variations in group types and sizes of blue sheep(Pseudoisnayaur)in the Helan M ountains,China.Acta Ecologica Sinica,2009,29(6):2782-2788.
[24]Cui D Y,Liu Z S,Wang X M,Zhai H,Hu T H,Li Z G.Winter food-habits of red deer(Cervus elaphus alxaicus)in Helan Mountains,China.Zoological Research,2007,28(4):383-388.
[25]Xing D L,Hao Z Q.The principle of maximum entropy and its applications in ecology.Biodiversity Science,2011,19(3):295-302.
[26]Phillips SJ,Anderson R P,Schapire R E.Maximum entropy modelling of species geographic distributions.Ecological Modelling,2006,190(3/4):231-259.
[27]Luo C,Xu W H,Zhou ZX,Ouyang ZY,Zhang L.Habitat prediction for forest musk deer(Moschusberezovskii)in Qinling mountain range based on niche model.Acta Ecologica Sinica,2011,31(5):1211-1229.
[28]Stachura-Skierczynska K,Tumiel T,Skierczyński M.Habitat prediction model for three-toed woodpecker and its implication for the conservation of biologically valuable forests.Forest Ecology and Management,2009,258(5):697-703.
[29]Haegeman B,Etienne R S.Entropy maximization and the spatial distribution of species.The American Naturalist,2010,175(4):74-90.
[30]Li M Y,Ju Y W,Kumar S,Stohlgren T J.Modelling potential habitat for alien species of Dreissena polymorpha in the continental USA.Acta Ecologica Sinica,2008,28(9):4253-4258.
[31]Swets J A.Measuring the accuracy of diagnostic systems.Science,1988,240:85-93.
[32]Xi Q.Research on Potential Distribution of the White-headed Langur Based on GISTechnology[D].NanJing:Nanjing Forestry University,2010.
[33]Oli M K.Seasonal patterns in habitat use of blue sheep Pseudoisnayaur(Artiodactyla,Bovidae)in Nepal.Mammalia,1996,60(2):187-193.
[34]Liu Z S,Cao L R,Wang X M,Li T,Li Z G.Winter bed-site selection by blue sheep(Pseudois nayaur)in Helan Mountains,Ningxia,China.Acta Theriologica Sinica,2005,25(1):1-8.
[35]Wang X M,Schaller GB.Status of large mammals in Inner Mongolia,China.Journal of East China Normal University:Special Issue of Mammals.1996,6(1):94-104.
[36]Wang X M,Liu ZS,Li X Q,Li Z G.Comparison of age structures of male blue sheep(Pseudois nayaur)on Helan Mountain in two periods,1995 and 2004.Zoological Research,2005,26(5):467-472.
參考文獻:
[1]王金亮,陳姚.3S技術在野生動物生境研究中的應用.地理與地理信息科學,2004,20(6):44-47.
[2]王秀磊.普氏原羚生境的景觀動態與適宜性評價研究[D].北京:中國林業科學研究院,2004.
[4]劉振生.巖羊 (Pseudois nayaur)生態學研究[D].上海:華東師范大學,2006.
[5]盛和林.毛皮動物手冊.上海:上海辭書出版社,1994:218-219.
[7]劉國庫,周材權,楊志松,龍帥,潘立,王維奎,游于群,曾國偉,耿山山.竹巴籠矮巖羊晝間行為節律和時間分配.生態學報,2011,31(4):972-981.
[8]昶野,劉振生,張明明,李志剛,胡天華.賀蘭山同域分布巖羊和馬鹿的夏季食性.生態學報,2010,30(6):1486-1493.
[10]王小明,劉志霄,徐宏發,李明,李元廣.賀蘭山巖羊種群生態及保護.生物多樣性,1998,6(1):1-5.
[12]劉振生,王小明,李志剛,崔多英,李新慶.賀蘭山巖羊冬春季取食生境的比較.動物學研究,2005,26(6):580-589.
[13]劉振生,王小明,李志剛,翟昊,胡天華.賀蘭山巖羊的數量與分布.動物學雜志,2007,42(3):1-8.
[15]徐衛華,歐陽志云,李宇,劉建國.基于遙感和GIS的秦嶺山系大熊貓生境評價.遙感技術與應用,2006,21(3):239-242.
[18]李紅梅,韓紅香,薛大勇.利用GARP生態位模型預測日本松干蚧在中國的地理分布.昆蟲學報,2005,48(1):95-100.
[20]李明陽,席慶,徐海根.基于WEB數據庫和Maxent模型的白頭葉猴潛在生境評價研究.林業資源管理,2010,(4):31-36.
[21]齊增湘,徐衛華,熊興耀,歐陽志云,鄭華,甘德欣.基于MAXENT模型的秦嶺山系黑熊潛在生境評價.生物多樣性,2011,19(3):343-352.
[22]徐衛華,羅翀.MAXENT模型在秦嶺川金絲猴生境評價中的應用.森林工程,2010,26(2):1-4.
[23]劉振生,李新慶,王小明,李志剛,胡天華,翟昊.賀蘭山巖羊 (Pseudois nayaur)集群特征的季節變化.生態學報,2009,29(6):2782-2788.
[24]崔多英,劉振生,王小明,翟昊,胡天華,李志剛.賀蘭山馬鹿冬季食性分析.動物學研究,2007,28(4):383-388.
[25]刑丁亮,郝占慶.最大熵原理及其在生態學研究中的應用.生物多樣性,2011,19(3):295-302.
[27]羅翀,徐衛華,周志翔,歐陽志云,張路.基于生態位模型的秦嶺山系林麝生境預測.生態學報,2011,31(5):1211-1229.
[30]李明陽,居云為,Kumar S,Stohlgren T J.美國大陸外來入侵物種斑馬紋貽貝 (Dreissena polymorp ha)潛在生境預測模型.生態學報,2008,28(9):4253-4258.
[32]席慶.基于GIS技術的白頭葉猴潛在分布研究[D].南京:南京林業大學,2010.
[34]劉振生,曹麗榮,王小明,李濤,李志剛.賀蘭山巖羊冬季對臥息地的選擇.獸類學報,2005,25(1):1-8.
[36]王小明,劉振生,李新慶,李志剛.賀蘭山雄性巖羊兩個時期生命表的比較.動物學研究,2005,26(5):467-472.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
石油瀝青(2021年4期)2021-10-14 08:50:44
世界科學技術-中醫藥現代化(2021年10期)2021-03-02 05:52:06
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
中國教育技術裝備(2015年19期)2015-03-01 02:43:07
中國工程咨詢(2015年2期)2015-02-14 02:59:26
俄羅斯問題研究(2012年1期)2012-03-25 09:54:51
