云南紅豆杉人工林萌枝特性
2013-12-20 03:05:58蘇建榮劉萬德李帥鋒
生態學報 2013年22期
蘇 磊,蘇建榮,劉萬德,李帥鋒
(中國林業科學研究院資源昆蟲研究所,昆明 650224)
萌枝現象普遍存在于許多古老植物的生長發育過程中,是它們在漫長的生存過程中進化出的生活史特征,萌枝在維持種群延續與穩定方面有著顯著的作用,是這些植物能夠經受不斷的環境改變、地質變化而不至滅絕的重要原因[1-2]。萌枝作為重要的植物功能特征[3],是一種重要的營養繁殖方式,在植被恢復和自然更新中有著重要的意義[4],是植物應對外界環境中各種干擾形成的有效適應策略。萌枝是持續生態位的占據者[5],也是高度進化的不穩定特征[6],以往對萌枝的研究集中于它們在群落演替[7]、物種保存[1]等方面的意義,影響萌枝能力的因素也有相關報道[8-9]。近些年在意識到萌枝的價值之后,一些與植物生長發育直接相關的研究工作也逐漸得到開展。李曉靖等通過冰雪災害后木荷的研究發現不同部位萌枝的光合生理特性存在差異[10];萌枝能力-高度權衡假說被學者提出并得到一定的認可,該假說認為萌枝生長消耗資源會限制高度生長[11-12],但有相反觀點認為萌枝不會帶來高度生長代價[13]。時至今日,萌枝是否會直接改變植物生長量的疑問仍未得到解答,萌枝能力-高度權衡假說的爭論也沒有一個明朗的結果。
云南紅豆杉(Taxus yunnanensis Cheng et L.K.Fu)是我國的國家一級保護植物,起源古老,與羅漢松科(Podocarpaceae)、粗榧科(Cephalotaxaceae)植物具有共同的祖先[14]。作為第三紀孓遺植物,云南紅豆杉具有明顯的萌枝現象[15]。瀕危植物云南紅豆杉富含天然抗癌藥物紫杉醇(Taxol),但是云南紅豆杉的資源量小、種群數量少,它的市場需求量卻非常大[15]。到目前為止,人工栽培紅豆杉是獲取紫杉醇最可行、最有效的手段[16]。云南紅豆杉是我國紫杉醇原料林的主要栽培樹種,在滇、川、渝、藏等省(市、自治區)紫杉醇原料林基地建設中有大規模種植[15]。鑒于云南紅豆杉既是重要的瀕危物種,又是價值很高的經濟植物,國內外學者圍繞它開展了大量的研究,內容涉及云南紅豆杉的種群生態學、細胞培養、組織培養、產紫杉醇內生真菌篩選、紫杉醇含量變異、苗木培育技術、原料林營建技術等諸多方面[16-23]。在這些研究中,尚未見云南紅豆杉的萌枝特性及萌枝對云南紅豆杉原料林產量和植株生長發育影響的研究報道。
枝葉生物量是云南紅豆杉藥用人工原料林的重要經濟性狀。生產上,一般采收云南紅豆杉枝、葉等提取紫杉醇及其前體以實現原料林的可持續利用[15],如何使云南紅豆杉人工林盡可能多的產出枝葉是一個迫切需要解決的問題。目前,云南紅豆杉萌枝與植株枝葉生物量的關系并不清楚,原料林培育是否需要區分萌枝類型進行選育、能否利用萌枝特性用于枝葉采收和原料林的持續經營等生產實際問題尚未解決。為此,本文通過云南紅豆杉人工林的萌枝情況、不同萌枝類型植株枝葉生物量等的調查、研究,探討云南紅豆杉人工種群萌枝性狀的發生率、萌枝的類型與數量分布;萌枝類型及數量與植株株高、地徑的關系;萌枝對植株枝葉生物量及其空間分布格局的影響等問題。以期為云南紅豆杉藥用原料林的良種選育、枝葉采收和可持續經營提供科學依據和理論支持。
1 材料與方法
1.1 研究地概況
研究地點設在云南省文山壯族苗族自治州馬關縣的金城林場。金城林場地處北緯22°59',東經104°26'。該林區海拔1600 m,為低緯度亞熱帶東部型山地季風氣候,雨季旱季界限明顯,且雨季氣溫較高,旱季氣溫較低。年均氣溫16.9℃,平均最高月氣溫21.7℃,平均最低月氣溫9.6℃。年平均降雨量1345 mm,相對濕度84%;年日照時數為1804 h,全年無霜期達300 d以上。金城林場于2005年開始在董亮營林區建設萬畝云南紅豆杉基地,現已建成30余hm2的云南紅豆杉采穗園,130余hm2的原料林基地[24]。
1.2 實驗材料與方法
1.2.1 試驗林概況
2011年10月初在金城林場云南紅豆杉基地選取了甲、乙、丙3塊樣地。所選3塊樣地的環境條件基本一致,具體情況見表1。樣地中的云南紅豆杉按臺地模式種植,種植密度為1株/(1.5 m×1.5 m),實生苗造林,定植時間2005年7月。整地、種植和撫育、管理過程中,所有樣地的營造林措施均采用同一技術標準。到2011年10月時,樣地內所種植云南紅豆杉的平均株高達2.17 m,平均地徑為44.61 mm,平均冠幅為1.2 m×1.2 m,發育良好,生長健壯,已達到可以進行商業采收的階段。
1.2.2 萌枝類型與數量的調查
從每塊樣地中分別選取植株長勢較接近,差異不大的180株云南紅豆杉作為樣株用于調查。選好樣株后,用標簽做好標記,然后逐株調查萌枝的類型和數量。萌枝的分類及判別采用Bellingham系統,該系統按以下標準把萌枝分為4種類型:(1)根萌枝,萌枝位置位于地下;(2)樹基萌枝,萌枝處于地上接近地面的位置;(3)樹干萌枝,萌枝位于樹冠,遠離地面;(4)枝萌枝,萌枝發生于非主干的枝條[2,25]。調查時,統一將地徑最粗、最高的莖干作為主干,分別用皮尺和游標卡尺測量每一植株的高度和地徑。
1.2.3 枝葉空間分布量的測定
研究萌枝對云南紅豆杉枝葉生長的空間分布時,在上述每塊樣地的180個樣株中分別隨機抽取15株用于生物量的測定。采收枝葉前,先按樹冠深度將樹冠等距離分為上層(Ⅰ)、中上層(Ⅱ)、中下層(Ⅲ)、下層(Ⅳ)4個層次并用油漆做上記號,然后采收各層次枝葉。采收后,迅速測定每個層次枝葉的重量。

表1 樣地概況Table 1 Outline of simple plots
1.3 數據處理
把樣地中有萌枝個體數/總個體數定義為有萌個體率;特定萌枝類型萌枝個體數/總個體數定義為特定萌枝類型的萌枝率[26]。研究萌枝數量與地徑、樹高的關系時,將3塊樣地的云南紅豆杉植株歸并為一個樣本,采用Spearman相關分析方法進行分析[26-27]。分析萌枝對枝葉生物量空間分布格局的影響時采用秩和檢驗法進行分析比較;不同萌枝類型的萌枝數量對枝葉空間分布的影響采用Kruskal-Wallis檢驗法分析。所有數據處理和統計分析均采用SPSS13.0軟件完成。
2 結果與分析
2.1 云南紅豆杉種群萌枝結構
云南紅豆杉有萌枝的比例則高達88.1%,萌枝現象十分普遍。在所調查的540株個體中,存在樹基萌枝的有432株,達到總數的80.0%;存在樹干萌枝的植株數量為214株,占總數的39.6%;兩種萌枝均有的是170株,占全部植株的31.5%(表2)。所調查的云南紅豆杉中,樹基萌枝最多的有8個,樹干萌枝最多的有5個,總萌枝數最多的有9個,總萌枝數集中于1—4個之間。無論是各類型的萌枝還是總體萌枝基本上呈現著萌枝數越多,植株數量越少的狀況(圖1)。

表2 各種萌枝方式的植株數量與比例Table 2 Number and rate of different sprouting types
2.2 萌枝數量與樹高和地徑的關系
云南紅豆杉的樹基萌枝數量與樹高之間呈正相關關系(R=0.52,P=0.00),樹干萌枝數量和樹高之間不存在顯著的相關關系(R=-0.04,P>0.05);樹基萌枝數量與地徑呈負相關關系(R= -0.46,P=0.00),樹干萌枝數量與地徑沒有顯著相關關系(R=-0.03,P>0.05);總萌枝數量與樹高和地徑之間的關系與它們和樹基萌枝數量的關系大致一樣,只是相關性低一些。簡而言之,云南紅豆杉隨著樹基萌枝或總萌枝數量的增多,樹干有增高而地徑有減小的趨勢(表3)。
2.3 萌枝數量與枝葉產出空間分布的關系
由表4可見,有無樹基萌枝的云南紅豆杉總體枝葉著生量并無顯著差異(P>0.05),同樣的情況也發生在有無樹干萌枝的植株中。盡管云南紅豆杉萌枝和總體枝葉生長量并無太大關系,但是萌枝對枝葉的空間分布格局有著較大的影響。樹基萌枝的植株其上層生長的枝葉極顯著多于無樹基萌枝的植株(P<0.01),在中上層的枝葉顯著多于無樹基萌枝的(P<0.05),但是它們的中下層枝葉量卻顯著小于無樹基萌枝的個體(P<0.05),下層的枝葉兩者之間沒有顯著的差異(P>0.05)。有樹干萌枝的植株上層、中上層和下層的分布的枝葉都稍微多于無樹干萌枝的,但它們的差異都未達到顯著水平,唯有中下層的枝葉分布量達到了顯著差異,有樹干萌枝的植株顯著少于沒有樹干萌枝的(P<0.05)。

圖1 云南紅豆杉的萌枝分布狀況Fig.1 Sprouting distribution status of Taxus yunnanensis

表3 云南紅豆杉不同萌枝類型的數量與樹高及地徑的關系Table 3 Relationship between different sprouting types and height&collar diameter in Taxus yunnanensis
從圖2可以看出,不管云南紅豆杉的萌枝方式怎樣,枝葉分布比例均以中下層和下層的居多。上層的枝葉分配比例都是有樹基萌枝或樹干萌枝時最多,無樹干萌枝時次之,無樹基萌枝時最少。沒有樹基萌枝的植株中上層枝葉也是最少的,但是它們的中下層和下層的枝葉卻都是四種萌枝狀況中最多的。
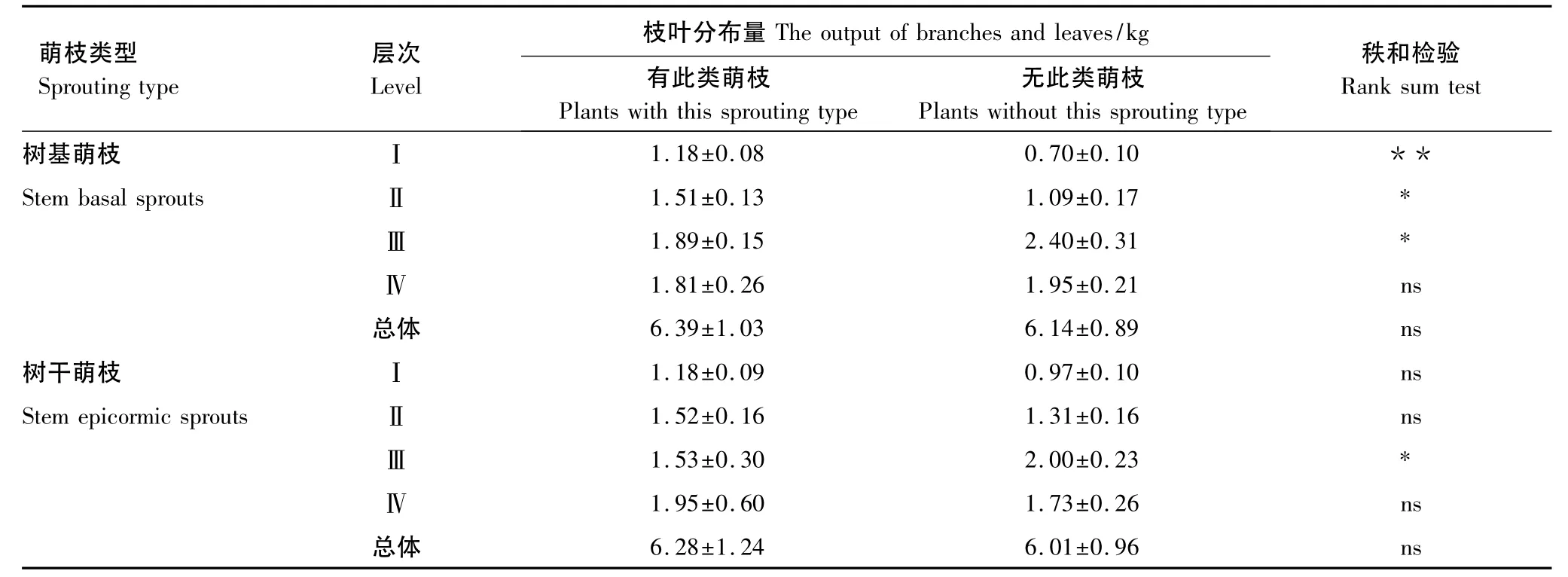
表4 云南紅豆杉各層次枝葉著生量與萌枝的關系Table 4 Relationship between growth amount of branch leaves and sprouts in Taxus yunnanensis
從圖1和表2可以看出云南紅豆杉樹基萌枝率很高,有1個、2個和3個樹基萌枝的植株最多,而且個體數也比較接近,故而對這3類萌枝的植株再單獨進行比較,以探究樹基萌枝數量對枝葉產出空間分布的影響,圖3為其Kruskal-Wallis檢驗結果,可以看出,云南紅豆杉的上層和中上層的枝葉量呈現著隨著樹基萌枝數量增加而增多的趨勢,3個樹基萌枝的云南紅豆杉的上層和中上層的枝葉產出顯著多于1個和2個樹基萌枝的植株(H=5.52,n=3,P<0.05),中上層的枝葉產出也顯著多于1個和 2個樹基萌枝的(H=4.60,n=3,P<0.05)。中下層的枝葉是1個樹基萌枝的植株最多,顯著高于2個和3個樹基萌枝的(H=10.59,n=3,P<0.05)。下層的枝葉分布相差不大,差異都未達到顯著程度(H=3.06,n=3,P=0.217)。
3 結論與討論
3.1 云南紅豆杉萌枝狀況剖析
紅豆杉屬植物已在地球上生存繁衍超過了250萬年之久[15],是第三紀孓遺植物。具備萌枝能力許多古老植物的普遍特征[3],它們在漫長的生存史中經歷了多次地質構造運動和劇烈的環境變化,各種嚴酷的干擾因素時常會在其尚未完成有性繁殖時就已經到來,植物無法通過有性繁殖產生后代來度過危機,萌枝更新是植物有性繁殖受阻時種群延續的一個有效的補充方式,許多古老植物正是得以依賴自身進化出的萌枝能力通過更新瓶頸期而始終不至于滅絕[1]。云南紅豆杉枝干受到外界環境強烈的干擾脅迫后,根部成為了最重要的營養和水分儲存場所,萌枝的發生需要母株的營養供給,樹基部分最接近根部,新生萌枝必然會最大程度的從此處發出,因此樹基萌枝便成為云南紅豆杉最主要的萌枝方式。樹冠是植物生長活躍的部位,但其距離根部較遠,且主干也可能由于病蟲害等外界干擾運輸能力大大減弱,因此樹冠處發出萌枝的概率會比較低。云南紅豆杉的萌枝情形正是其祖先長期生活在干擾脅迫環境下而進化出的適應能力的一個具體表達。
3.2 云南紅豆杉萌枝與植株的生長發育

圖2 不同萌枝狀況的云南紅豆杉枝葉空間分配格局Fig.2 Branches and leaves spatial distribution pattern of Taxus yunnanensis with different sprouting status
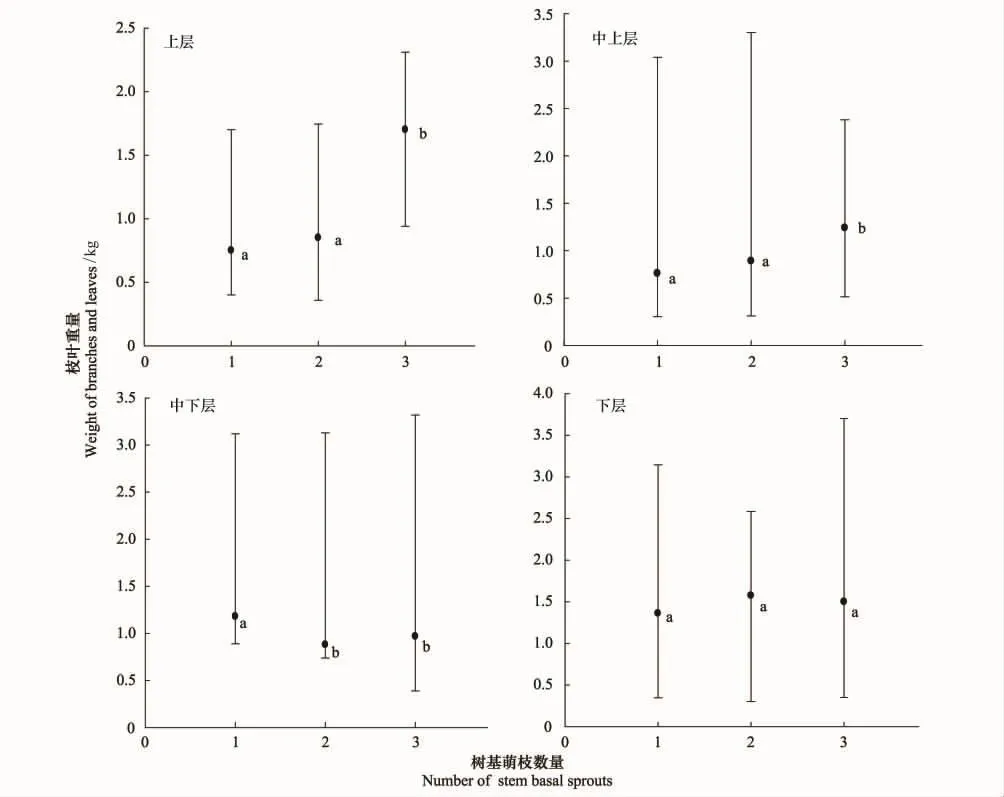
圖3 不同樹基萌枝程度的枝葉空間分布比較Fig.3 Branches and leaves spatial distribution comparison of different stem basal sprouting levels
根據萌枝能力-高度權衡假說,萌枝的出現過程是一個消耗物質和能量的過程,萌枝生長的投入會限制植物其他方面的生長,一個最顯著的例子就植物的高度生長受到制約,于是就造成了沒有萌枝的植物往往高于有萌枝的植物,萌枝少的植物高于萌枝多的植物[11]。但在本研究中卻是萌枝數量多的云南紅豆杉植株高于萌枝數量少的,這與趙睿等人對米心水青岡(Fagus engeriana)的研究結果相近[13]。萌枝能力-高度權衡假說是建立在不同植物的比較的基礎上的,且只考慮到了萌枝生長對資源的消耗,因此必然存在著一定的局限性。萌枝的發生無疑會給主干帶來一定的生長代價,造成主干資源向外轉移。云南紅豆杉樹基萌枝的枝干與主干空間重疊部分最多,與主干的資源競爭也最激烈,故而它們對主干的消耗尤為明顯,這就造成了樹基萌枝越多,主干越細的現象。調查時發現萌枝與主干幾乎呈平行狀態,各個萌枝都與主干一起競爭利用光資源,植株的萌枝越多,主干與萌枝間的競爭就愈激烈,為了占據有利的光環境,主干和萌枝都采取了盡可能增加高度的生存策略[28]。
植物的冠層結構不是一成不變的,而是可以發生動態變化的。植株內部主干與萌枝的競爭過程也是枝葉空間格局重新調整和分配的過程。萌枝與主干的競爭使得有萌枝的云南紅豆杉的部分枝葉向上轉移,同時,萌枝會影響到枝葉所處的光照環境,冠層上半部分枝葉濃密時,中下層的蔭蔽程度高,這一部位的枝條自然修剪的幾率高,枝條壽命相對較短[29]。中下層的枝葉在上述因素共同作用下分布有所減少。這些現象在不同的萌枝狀況下會略有差異。幾種萌枝狀況中有樹干萌枝的植株中下層枝葉產出是最少的,這是因為樹干萌枝最開始從主干的上部發出,隨著樹體的生長,其原始發生區基本上成了植冠的中間部位,樹干萌枝的生長極大地限制了樹冠中下層的養分供給,阻礙了這一區域枝葉的生長發育。沒有樹基萌枝這一主要萌枝方式的云南紅豆杉個體枝干間競爭壓力最小,主干發育最為良好,樹冠也更接近錐體形,因此相對于其他植株,它們的冠幅較寬,枝葉量的空間分布也更接近上少下多的金字塔形。
萌枝影響了云南紅豆杉幼樹的枝葉空間分布格局,但是卻沒有顯著改變總體的枝葉產出。究其原因,一方面是由于萌枝消耗了主干的養分資源,減少了主干著生更多枝葉的幾率,但是萌枝上也有枝葉可以制造光合產物,并不是單純的資源消耗者,萌枝上的枝葉恰恰彌補了主干所減少的枝葉;另一方面,主干高度隨著萌枝數量的增多而有一定的增大,但是由于地徑與樹高的權衡關系,較小的地徑成了限制枝葉產量增加的重要因素。本文的調查的對象是尚未受干擾或是只是受到輕微干擾的云南紅豆杉幼樹,而萌枝是植物適應外界干擾而進化出的營養繁殖策略,萌枝的巨大價值應該會在受到較強烈干擾的云南紅豆杉群體中有更大的體現。
3.3 云南紅豆杉萌枝的應用展望
許多的古老植物如水青岡屬 (Fagus)[13,30-33]、樺木屬 (Betula)[34-35]、櫟屬(Quercus)[36-39]、紅豆杉屬(Taxus)[40]都具有較強的萌枝能力。在植株的主干遭受破壞之后,萌枝在受損的植株恢復過程中發揮著至關重要的作用,存活的萌枝有機會生出新的根系,逐漸占據一定的空間并獨立成活。萌枝有著廣泛的實踐應用前景,有研究發現對遼東櫟萌生灌叢的萌枝數量進行調控會顯著影響植株的高生長和有性繁殖[39],哀牢山中的居民在選擇殼斗科植物作為薪柴時也不自覺的利用了它們所具有萌枝能力[41]。萌枝與云南紅豆杉的生理發育有著密切的聯系,筆者認為若是對云南紅豆杉的萌枝狀況進行合理控制,例如使用機械方法讓萌枝與主干產生一定的分離,使樹冠擴展而獲取較原來更多的光資源,可能會使植株的枝葉著生量有明顯的提升;或者是設計采收方案時有意保留不同數量和類型的萌枝跟蹤調查其生長發育情況以選取枝葉產出最多的萌枝形式。采收枝葉后的云南紅豆杉的萌枝情況也可能發生改變,它們對以后枝葉產出的影響也需要認真研究。采收人工栽培的云南紅豆杉的枝葉作為原材料是獲取紫杉醇的主要來源,然而云南紅豆杉的生長卻是十分緩慢,紫杉醇原料來之不易,萌枝存在恢復種群穩定的潛能力[2],如何合理利用這一特性進行調控以便最大量的持續收獲枝葉是一個亟待開展的新課題。
[1]Zhang S,Tang Y,Wang J,Ahou L Y,Jia Y H.Propagation of Magnolia sargentiana Rehder& Wilson(Magnoliaceae)by sprouting and its role in biodiversity conservation.Acta Botanica Boreali-Occidentalia Sinica,2010,30(4):769-775.
[2]Bellingham P J,Sparrow A D.Resprouting as a life history strategy in woody plant communities.Oikos,2000,89(2):409-416.
[3]Loehle C.Strategy spaceand thedisturbance spectrum:a life-history model for tree species coexistence.The American Naturalist,2000,156(1):14-33.
[4]Chen M,Cao M,Lin X L.Research advances in regeneration of woody plants by sprouting.Chinese Journal of Ecology,2007,26(7):1114-1118.
[5]Bond W J,Midgley J J.Ecology of sprouting in woody plants:the persistence niche.Trends in Ecology and Evolution,2001,16(1):45-51.
[6]Bond W J,Midgley J J.The evolutionary ecology of sprouting in woody plants.International Journal of Plant Sciences,2003,164(3):S103-S114
[7]Miller P M,Kauffman J B.Seedling and sprout response to slash-and-burn agriculture in a tropical deciduous forest.Biotropica,1998,30(4):538-546.
[8]Espelta J M,Retana J,Habrouk A.Resprouting patterns after fire and response to stool cleaning of two coexisting Mediterranean oaks with contrasting leaf habits on two different sites.Forest Ecology and Management,2003,179(1/3):401-414.
[9]Hoffman W A,Bazzaz FA,Chatterton N J,Harrison P A,Jackson R B.Elevated CO2enhances resprouting of a tropical savanna tree.Oecologia,2000,123,(3):312-317.
[10]Li X J,Zhou B Z,Cao Y H,Wang G,Hong Y F.Photosynthetic physiology of Schima superba sprouts after the ice and snow disaster in 2008.Chinese Journal of Ecology,2011,30(12):2753-2760.
[11]Midgley JJ.Why the world's vegetation is not totally dominated by resprouting plants;because resprouters are shorter than reseeders.Ecography,1996,19(1):92-94.
[12]Falster D S,Westoby M.Tradeoffs between height growth rate,stempersistenceand maximum height among plant species in a post-firesuccession.Oikos,2005,111(1):57-66.
[13]Zhao R,Zhou X F,Xu N N,Zhao M S,Liu L,Chen X Y.Sprouting and height in a Fagus engeriana population.Acta Ecologica Sinica,2009,29(7):3665-3669.
[14]Su J R,Zhang ZJ,Deng J,Li GS.Relationships between Geographical Distribution of Taxuswallichiana and Climate in China.Forest Research,2005,18(5):510-515.
[15]Wang W B,Wang D M.Taxus yunnanensis.Kunming:Yunnan University Press,2006.
[16]Su JR,Miao Y C,Zhang Z J.RAPD Markers Related to Taxol Content of Taxus yunnanensis.Scientia Silvae Sinicae,2009,45(7):16-20.
[17]Su J R,Zhang Z J,Deng J,Chen Z Y.Population structure and life table of the endangered population of Taxus yunnanensis.Forest Research,2005,18(6):651-656.
[18]Zhang C H,Wu J Y,He JY.Effects of inoculum size and age on biomass growth and paclitaxel production of elicitor-treated Taxus yunnanensis cell cultures.Applied Microbiology and Biotechnology,2002,60(4):396-402.
[19]Wang S,Jia Y J,Wei F,Lu H Z,Wu SF.Tissue culture and plant regeneration of Taxus Yunanensis.Acta Botanica Yunnanica,1997,19(4):407-410.
[20]Chen Y J,Zhang Z,Wang Y,Su Y,Zhang R.Screening endophytic fungus to produce taxol from Taxus Yunnanensis.Biotechnology.2003,13(2):10-11.
[21]Qing W Y,Diao Y G,Yuan Z M.Raising seedlings by cuttage of Taxus yunnanensis.Journal of Sichuan Forestry Science and Technology,2005,26(1):76-77.
[22]Wang C W,Peng SL,Li M G,Li W J,Lin F L.Review of Factors Affecting the taxoids content of Taxus Spp.Acta Ecologica Sinica,2006,26(5):1583-1590.
[23]Zhou Z C,Su JR,Cheng G Y.Medicinal Plantation Cultivation Practical Technology of Taxus.Beijing:China Forestry Press,2010.
[24]Jin G M,Wang W B,Zhang J F,Yuan F.Growth rate and biomass comparison of four years Taxus yunnanesis propagated from seeding and cutting.Shaanxi Forest Science and Technology,2009,(4):1-5.
[25]Su JR,Liu WD,Zhang Z J,Li SF.Sprouting characteristic in restoration ecosystems of monsoon evergreen broadleaved forest in south-central of Yunnan Province.Acta Ecologica Sinica,2012,32(2):805-814.
[26]Nanami S,Kawaguchi H,Tateno R,Li C H,Katagiri S.Sprouting traits and population structure of co-occurring Castanopsis species in an evergreen broad-leaved forest in southern China.Ecological Research,2004,19(3):341-348.
[27]Yang Y C,Mu JP,Tang CQ,Yang K.Community structure and population regeneration in remnant Ginkgo biloba stands.Acta Ecologica Sinica,2011,31(21):6396-6408.
[28]Graciaa M,Retana J.Effect of site quality and shading on sprouting patterns of holm oak coppices.Forest Ecology and Management,2004,188(1/3):39-49.
[29]Zhan F,Yang D M.Relationships among light conditions,crown structure and branch longevity:a case study in Osmanthus fragrans and Metasequoia glyptostroboides.Acta Ecologica Sinica,2012,32(3):984-992.
[30]Ohkubo T,Tanimoto T,Peters R.Response of Japanese beech(Fagus japonica Maxim.)sprouts to canopy gaps.Vegetatio,1996,124(1):1-8.
[31]He JS,Liu F,Chen W L,Chen L Z.History of disturbance and regeneration strategies of Fagus engleriana and Quercus aliena var.acuteserrata forests in Shennongjia,Hubei Province.Acta Botanica Sinina,1999,41(8):887-892.
[32]He J S,Liu F,Chen W L.Study on the sprouting process of Fagus engleriana in Shennongjia Mountains.Acta Phytoecologica Sinica,1998,22(5):385-391.
[33]Yu S,Wang R,Liu M,Yang S Z,Luo Y,Chen X Y.Species composition and sprouting regeneration of Fagus engleriana community in Tianmushan Mountain.Chinese Journal of Ecology,2009,28(2):182-187.
[34]Johansson T.Sprouting ability and biomass production of downy and silver birch stumps of different diameters.Biomass and Bioenergy,2008,32(10):944-951.
[35]Liu Z L,Fang SZ,Liu D,Yu M K,Tang L Z.Influence of thinning time and density on sprout development,biomass production and energy stocks of sawtooth oak stumps.Forest Ecology and Management,2011,262(2):299-306.
[36]Fang SZ,Liu Z L,Cao Y D,Liu D,YU M K,Tang L Z.Sprout development,biomass accumulation and fuelwood characteristics from coppiced plantations of Quercus acutissima.Biomass and Bioenergy,2011,35(7):3104-3114.
[37]Pausas JG.Resprouting of Quercus suber in NE Spain after fire.Journal of Vegetation Science,1997,8(5):703-706.
[38]Gao X M,Wang W,Du X J,Ma K P.Size structure,ecological significance and population origin of Quercus wutaishanica forest in Beijing mountainous area.Acta Phytoecologica Sinica,2001,25(6):673-678.
[39]Meng L B,Bao W K,Pang X Y,Sun F.Effects of ramets adjustment on Quercus liaotungensis growth and seed generation.Chinese Journal of Applied Ecology,2006,17(10):1771-1776.
[40]Li X K,Xiang W S,Su Z M.Structure and dynamics of Taxuschinensis var.mairei clonal population.Chinese Journal of Applied Ecology,2004,15(2):177-180.
[41]He Y T,Cao M,Tang Y,Li GC.A preliminary study on sprouting of canopy trees in middle mountain moist evergreen broad-leaved forest of Ailao mountain,Yunnan.Journal of Wuhan Botanical Research,2000,18(6):523-527.
參考文獻:
[1]張松,唐亞,王靜,阿侯拉葉,賈銀輝.凹葉木蘭萌枝更新及其在物種保存中的意義.西北植物學報,2010,30(4):769-775.
[4]陳沐,曹敏,林露湘.木本植物萌生更新的研究進展.生態學雜志,2007,26(7):1114-1118.
[10]李曉靖,周本智,曹永慧,王剛,洪奕豐.南方冰雪災害后受害木荷萌枝光合生理特性.生態學雜志,2011,30(12):2753-2760.
[13]趙睿,周學峰,徐娜娜,趙明水,劉亮,陳小勇.米心水青岡種群萌條更新與高度生長.生態學報,2009,29(7):3665-3669.
[14]蘇建榮,張志鈞,鄧疆,李國松.云南紅豆杉的地理分布與氣候關系.林業科學研究,2005,18(5):510-515.
[15]王衛斌,王達明.云南紅豆杉.昆明:云南大學出版社,2006.
[16]蘇建榮,繆迎春,張志鈞.云南紅豆杉紫杉醇含量變異及其相關的RAPD分子標記.林業科學,2009,45(7):16-20.
[17]蘇建榮,張志鈞,鄧疆,陳志勇.云南紅豆杉種群結構與生命表分析.林業科學研究,2005,18(6):651-656.
[19]王水,賈勇炯,魏峰,盧慧宗,伍順鳳.云南紅豆杉的組織培養及植株再生.云南植物研究,1997,19(4):407-410.
[20]陳毅堅,張灼,王艷,蘇源,張睿.云南紅豆杉(Taxus Yunnanensis)內生真菌中產紫杉醇真菌的篩選.生物技術,2003,13(2):10-11.
[21]卿文英,刁陽光,袁志明.云南紅豆杉的扦插育苗方法.四川林業科技,2005,26(1):76-77.
[22]王昌偉,彭少麟,李鳴光,李文建,林發亮.紅豆杉中紫杉醇及其衍生物含量影響因子研究進展.生態學報,2006,26(5):1583-1590.
[23]周志春,蘇建榮,程廣有.紅豆杉藥用林高效栽培實用技術.北京:中國林業出版社,2010.
[24]金桂梅,王衛斌,張勁峰,袁鋒.四年生云南紅豆杉實生苗與扦插苗生長量和生物量比較研究.陜西林業科技,2009,(4):1-5.
[25]蘇建榮,劉萬德,張志鈞,李帥鋒.云南中南部季風常綠闊葉林恢復生態系統萌生特征.生態學報,2012,32(2):805-814.
[27]楊永川,穆建平,Tang C Q,楊軻.殘存銀杏群落的結構及種群更新特征.生態學報,2011,31(21):6396-6408.
[29]占峰,楊冬梅.光照條件、植株冠層結構和枝條壽命的關系——以桂花和水杉為例.生態學報,2012,32(3):984-992.
[31]賀金生,劉峰,陳偉烈,陳靈芝.神農架地區米心水青岡林和銳齒槲櫟林群落干擾歷史及更新策略.植物學報,1999,41(8):887-892.
[32]賀金生,劉峰,陳偉烈.神農架地區米心水青岡萌枝過程的研究.植物生態學報,1998,22(5):385-391.
[33]于碩,王嶸,劉敏,楊淑貞,羅遠,陳小勇.天目山米心水青岡群落物種組成特點與萌條更新.生態學雜志,2009,28(2):182-187.
[38]高賢明,王巍,杜曉軍,馬克平.北京山區遼東櫟林的徑級結構、種群起源及生態學意義.植物生態學報,2001,25(6):673-678.
[39]孟令彬,包維楷,龐學勇,孫凡.萌蘗調控對遼東櫟留存萌生株生長與結實的影響.應用生態學報,2006,17(10):1771-1776.
[40]李先琨,向悟生,蘇宗明.南方紅豆杉無性系種群結構和動態研究.應用生態學報,2004,15(2):177-180.
[41]何永濤,曹敏,唐勇,李貴才.云南省哀牢山中山濕性常綠闊葉林萌生現象的初步研究.武漢植物學研究,2000,18(6):523-527.
猜你喜歡
世界科學技術-中醫藥現代化(2022年2期)2022-05-25 13:17:14
昆明醫科大學學報(2021年12期)2021-12-30 07:00:10
實用口腔醫學雜志(2017年6期)2017-09-19 02:51:06
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中外醫療(2016年15期)2016-12-01 04:25:50
哈爾濱醫藥(2015年2期)2015-12-01 03:57:41
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
