丹頂鶴春遷期覓食棲息地多尺度選擇
——以雙臺河口保護區為例
2013-12-21 04:59:54吳慶明鄒紅菲金洪陽馬建章
生態學報 2013年20期
吳慶明,鄒紅菲,金洪陽,馬建章
(東北林業大學野生動物資源學院,哈爾濱 150040)
最新研究表明:丹頂鶴(Grus japonensis)大陸種群西線群體的越冬種群數量急劇下降,每年以50—150只的速度從1100余只降到不足500只;并且越冬區域縮小為20世紀80年代的8%[1]。遼寧雙臺河口國家級自然保護區(以下簡稱雙臺河口保護區)是丹頂鶴大陸種群西線群體繁殖的南限[2]和越冬的北限,更是遷徙必經的主要停歇地;據1991—2012年間20余年的丹頂鶴遷徙種群監測,1995年種群數量最低為308只,2000年最高為808只,2011和2012年度均穩定在350—500只[3]。可見,雙臺河口保護區對于西線丹頂鶴群體種群數量的維持具有重要的生態意義。景觀監測結果顯示出[4-5]:自1987年以來的20年間,雙臺河口保護區內景觀斑塊破碎化趨勢明顯,蘆葦沼澤等天然濕地面積逐年減少;人為因素如油田開發建設等已成為保護區濕地景觀格局變化的主導因素。由此產生的管理不規范問題已引起中華人民共和國環境保護部的高度重視并掛牌督辦。在這種特殊環境情況下,非常有必要對丹頂鶴種群及其棲息地選擇情況進行監測,以便為丹頂鶴的科學保護提供理論參考。
在鳥類棲息地選擇方面,越來越多的研究證實[6-8]:鳥類的棲息地選擇具有多尺度特征,僅單一尺度的研究難以反映鳥類棲息地選擇的真實情況,多尺度研究能促進對鳥類及其棲息資源的認識和有效保護。關于多尺度的概念,最早源于1949年,Svardson認為鳥類的生境選擇模式應分為兩個過程:首先利用環境的一般特征并在不同的環境中進行粗放性選擇,然后再據某些精細的特征來選擇特定的生存環境[9];之后,多尺度的概念被廣泛應用于鳥類棲息地選擇研究中[10-15];我國的研究相對較少,主要針對丹頂鶴、白冠長尾雉、褐馬雞等瀕危特有鳥類進行了探討[16-20];其中,關于丹頂鶴,主要對大陸種群的主要繁殖地扎龍保護區丹頂鶴的繁殖棲息地選擇[16]和越冬地黃河三角洲保護區非繁殖期丹頂鶴覓食棲息地選擇[17-18]進行了多尺度特征的研究。雙臺河口保護區作為丹頂鶴大陸種群的特殊分布地——以遷徙停歇地為主、集越冬地和繁殖地于一體,從多尺度層面對春遷期丹頂鶴覓食棲息地選擇進行分析非常必要。
1 研究地概況
研究地位于遼寧省盤錦市境內雙臺河入海口處的雙臺河口保護區內(121°30'—122°00'E,40°45'—41°10'N),總面積12.8萬hm2[21];該保護區屬于海岸濕地,主要由蘆葦沼澤、濕草甸、人工濕地(主要為稻田和水庫)、河道、河口、泥灘、淺海海域等組成[3],成為我國高緯度地區面積最大的濱海蘆葦沼澤區[5]。保護區內大面積的翅堿蓬(Suaeda heteroptera)灘涂和淺海海域是環西太平洋遷徙的鳥類停歇的主要分布區[5];尤其是大面積的蘆葦沼澤,是春季遷徙期丹頂鶴停歇補充體能的重要棲息地,期間丹頂鶴也會偶爾選擇灘涂區域內的農田。關于蘆葦沼澤,為了利于翌年的生境管理與監測及葦草的萌發,蘆葦沼澤的利用者會通過火燒的方式對那些無法收割與處理的剩余蘆葦和草叢進行處理;每年春季,過火的葦草區會成為蘆葦沼澤內部或邊緣一種特殊的人為干預生境,這些區域內的根芽、種子等會成為丹頂鶴的主要食物。本文的調查主要發生在核心區內的黑嘴鷗繁殖地、四干北至向陽管區的南界、羅家分場、三道溝,緩沖區內四干南、向陽管區、孫家流子分場、營口垛場、八仙分場、酒壺嘴和南井子分場等區域。
2 研究方法
野外觀察時間為2008—2010年3—4月。
首先,憑借摩托車和自行車等交通工具采用路線法和定點觀察法通過10倍雙筒和20—60倍單筒望遠鏡對保護區內不同類型生境進行瀏覽式觀察,搜索該時期遷來停歇的丹頂鶴,觀察到丹頂鶴后記錄其晝間停歇棲息的生境類型和區域;然后,針對監測到的丹頂鶴晝間停歇區,連續2—5d進行觀察以確定其覓食棲息地的固定性,并記錄生境類型和大致范圍;之后,通過丹頂鶴足跡、糞便和自然標識物及激光測距儀等實地確認丹頂鶴覓食區的準確范圍,并用GPS定位,若發現GPS定位點不重復則在確定的覓食區范圍內,通過二維坐標法和樣方法進行樣方布設和環境因素測量[21]。
具體的樣方布設方法[22-23]為:在丹頂鶴覓食區范圍內,隨機選取一點O,過O點作任意的兩條垂線,在2條垂線上距離O點0、5、10m、15 m處分別設1 m×1 m樣方各1個,記錄各種環境因素,取其均值。3a間共確認丹頂鶴春季覓食區90個(其中,蘆葦沼澤覓食區81個),總樣方1170個,有效樣方1067個。
環境因素[21,24]主要包括植被因素和距離因素。其中,植被因素主要包括:植被密度(X1,根/m2,1 m×1 m樣方內植被的密度,主要為蘆葦植被的密度)、植被高度(X2,cm,1 m×1 m樣方內植被的平均高度,主要為蘆葦植被的高度)、植被直徑(X3,mm,1 m×1 m樣方內植被的平均直徑,主要為蘆葦植被的直徑)。距離因素主要包括:明水面距離(X4,m,距最近的且面積大于1 m2的明水面距離)、堤壩距離(X5,m,距最近的堤壩距離)、道路距離(X6,km,距最近的道路距離)、火燒地距離(X7,m,距最近的火燒地距離)、油井距離(X8,km,距最近的油井距離)、居民區距離(X9,km,距最近的村屯距離)。
通過SPSS 13.0中的One-way ANOVA、Factor Analysis對數據進行分析。
3 結果與分析
3.1 春遷期丹頂鶴覓食棲息地類型及群體分布
調查期間,共累積觀察確定丹頂鶴覓食棲息地90處,統計結果顯示出(圖1):在雙臺河口保護區,春遷期丹頂鶴的覓食棲息地類型多樣化,包括蘆葦沼澤、玉米地、泥灘、草甸四種,蘆葦沼澤為偏好覓食生境(90.00%)。種群分布結果顯示出(圖2):蘆葦沼澤中覓食分布的丹頂鶴種群數量最大(92.29%)。綜上所述,雙臺河口保護區,蘆葦沼澤是春遷期丹頂鶴偏好選擇利用的主要覓食棲息地。

圖1 春遷期丹頂鶴覓食棲息地類型Fig.1 Feeding habitat type of Red-crowned crane during spring migration
3.2 春遷期丹頂鶴覓食棲息地選擇
對雙臺河口保護區內丹頂鶴春遷期覓食棲息地選擇因素進行因子分析,結果表明:前4個因子的特征值均大于1(表1),相關性較小;其余因子的特征值均小于1,相關性較大;前4個因子的累積貢獻率為84.069%(表1),超過60%,基本包含了全部9個環境因素所具有的信息,能夠反映影響春遷期丹頂鶴覓食棲息地選擇的絕大部分信息,故本文取前4個因子并計算其相應的特征向量(表2),將其分別定義為微生境因子、宏生境因子、強干擾因子和弱干擾因子。
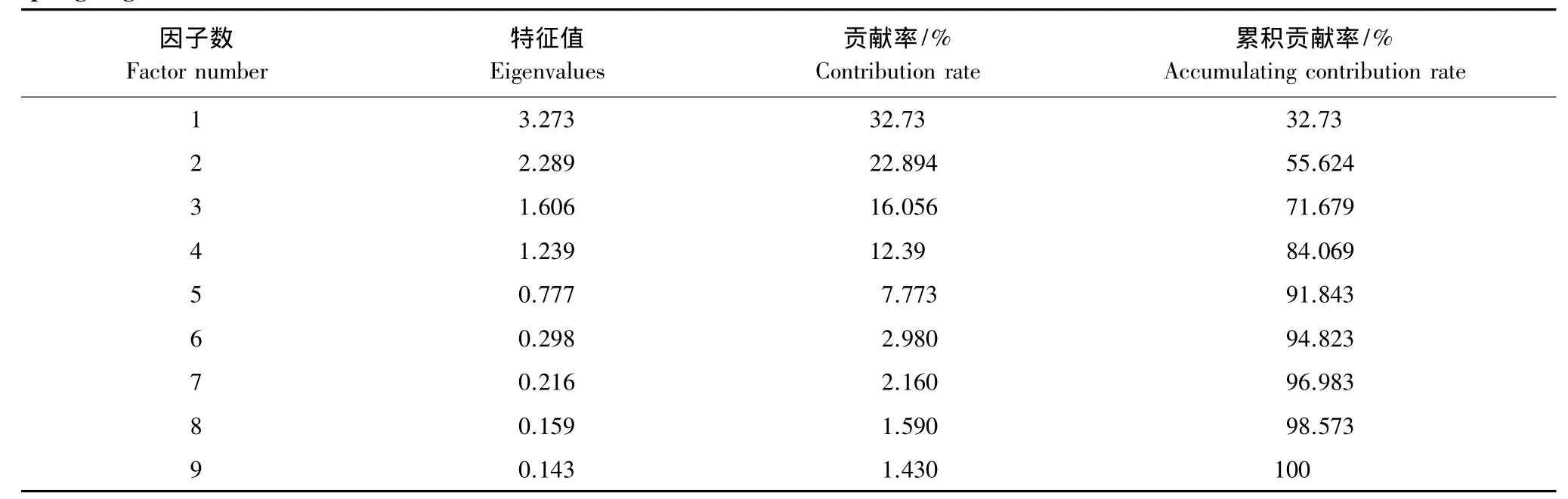
表1 丹頂鶴春遷期覓食棲息地選擇因素因子分析的累積貢獻率Table 1 Accumulating contribution rate of factor analysis on environment factor for feeding habitat selection of Red-crowned crane during spring migration
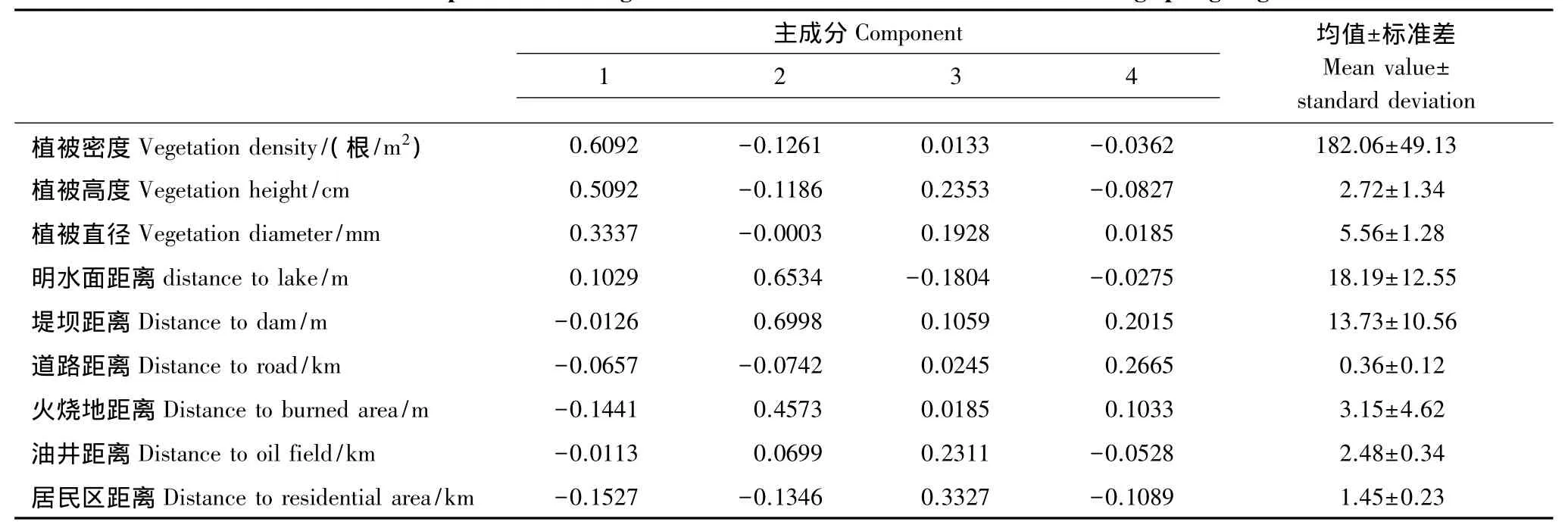
表2 春遷期丹頂鶴覓食棲息地選擇的主成分分析Table 2 Main component of feeding habitat selection for Red-crowned crane during spring migration
4 討論
4.1 覓食棲息地類型
關于春遷期丹頂鶴覓食棲息地類型,與扎龍保護區的研究結果[25]存有異同;相似之處在于春遷期丹頂鶴偏好選擇的覓食棲息地類型一致,均為蘆葦沼澤;相異之處在于丹頂鶴擴展選擇的覓食棲息地類型不同,扎龍保護區的丹頂鶴僅擴增為苔草沼澤,而雙臺河口保護區的丹頂鶴擴增為玉米地、泥灘、草甸3種。
兩個保護區的植被類型均以蘆葦沼澤為優勢植被,這也是丹頂鶴該時期階段偏好選擇的植被類型,不同區域的主要研究結果一致符合丹頂鶴該時期的行為方式和區域優勢植被特征。由于地理位置的差異,雙臺河口保護區內存有一定面積的灘涂,灘涂分布有許多土壤動物[26],這也是鳥類的主要食物來源[27];但由于此時的灘涂尚未解凍,不適宜作為春遷期丹頂鶴偏好選擇的覓食棲息地,故只有少數量的丹頂鶴偶爾選擇該區域進行擇食。關于玉米地,兩個保護區的調查范圍也存在差異;扎龍保護區的調查范圍并非整個保護區,而是以保護區局址為核心的一定半徑的區域內,該區域內玉米地較少且面積也較小,因此扎龍保護區的調查結果只意味著其區域內未記錄玉米地這一覓食棲息地類型,并不意味著玉米地在丹頂鶴春遷期的覓食棲息地選擇范圍之外,該方面還需要通過對整個區域進行全面調查才能得出科學的結論。
4.2 覓食棲息地選擇主成分的判定
第一主成分中,植被密度、植被高度、植被直徑的系數較大。由統計結果可以看出,雙臺河口保護區,春遷期丹頂鶴覓食棲息地的植被特征(植被密度均為182.06根/m2、植被高度均為2.72 cm、植被直徑均為5.56 mm)具有密度大、高度小、直徑小的特征。野外觀察發現,該時期,保護區內植被上仍覆蓋著部分的冰雪,只有部分區域因陽光輻射、地表積溫高或處于風口位置而呈少量冰、雪、水混合體或冰、水混合體或完全無雪狀態,這種自然狀況的區域,植被的根基位置以一定面積裸露出來,且植被高度的特征小于丹頂鶴的平均喙長(16.3 cm),這樣的環境便于丹頂鶴識別并尋覓植被根基或土壤中的食物,丹頂鶴覓食時留下的足跡與啄跡等印痕特征便是丹頂鶴利用類似生境的一個佐證。植被密度和植被直徑直接關系著植被蓋度(草本植物以植被的根蓋度為主,為植被密度與植被根面積二者均值的乘積,植被根面積由植被直徑求出),若以二者為變量進行計算,春遷期丹頂鶴覓食棲息地的植被蓋度約為(0.442±0.007)%。綜上所述,該主成分包括的植被密度、植被高度、植被直徑3個環境因素直接關系丹頂鶴該時期具體覓食點的信息,屬于覓食棲息地利用的小尺度也即微環境特征,將其定義為微生境因子。
在第二主成分中,明水面距離、堤壩距離、火燒地距離的系數較大。由于油氣建設和旅游開發的需要,雙臺河口保護區內修建了481.2 km的穿越核心區、緩沖區、實驗區的163條油田專用道路和旅游觀光道路(中華人民共和國環境保護部,2012年2月24日),這些道路使得保護區內呈現出規則斑塊分布的景觀格局特征。春遷期丹頂鶴的覓食棲息地也均分布于這些斑塊中,這些斑塊多被道路、堤壩和堤壩下的水渠所包圍,堤壩與水渠相鄰,二者與道路有一定的距離。由于冬季收割的不便性,堤壩上較高的蘆葦和雜草在不同年度的冬季均被保留下來,這些葦草常在翌年春季通過火燒的方式被處理。每年春季,堤壩周邊及水渠周邊區域便成了火燒地,該區域的地面裸露度較高,土壤動物復蘇較早,葦草萌芽也較早,干擾較少的類似區域常成為春遷期丹頂鶴首選的覓食地,調查的結果也印證了這一點,90.00%的覓食區和94.59%的鶴群均位于火燒地內或火燒地邊緣。本文的分析結果與此相吻合,丹頂鶴覓食棲息地距明水面距離、堤壩距離和火燒地距離均較近(30 m內),距火燒地最近(10 m內)。可見,明水面、堤壩、火燒地成為春遷期丹頂鶴從大尺度角度即宏生境尺度選擇覓食棲息地必不可少的環境特征,對于具備這3個景觀條件的棲息地,丹頂鶴可以根據干擾情況進行酌情的覓食選擇。基于此,將第二主成分定義為宏生境因子。
任何野生動物,都對干擾保持一定的敏感度,都有一定的安全距離,不同的干擾強度有不同的安全距離,丹頂鶴也不例外。在保護區內,丹頂鶴春季遷來期間,主要的干擾為非自然干擾,包括人的干擾和動物的干擾。動物的干擾主要包括狗等家畜和狐貍、獾等哺乳類的干擾,雙臺河口保護區內,該類干擾基本沒有。人的干擾主要包括居民區和油田作業區等強度較大的干擾和道路上的行人、行車等強度較小的干擾。居民區和油田作業區無季節性,常年有活動,對丹頂鶴的擾動較大;道路上行人、行車具有季節性,不同季節內行人和行車的頻率不同,春季屬于人車活動較少的季節,且人車存留時間短。本文的分析結果與此一致,油田作業區和居民區被判定為第三主成分,道路被判定為第四主成分。分析結果顯示出,春遷期丹頂鶴覓食棲息地距居民區距離平均保持在1 km以上,距油井距離平均保持在2 km以上,距道路距離300—500 m。基于此,將第三主成分定義為強干擾因子,將第四主成分定義為弱干擾因子。
4.3 覓食棲息地選擇分析
針對處于遷徙狀態的候鳥而言,遷徙過程中停歇點的唯一貢獻是提供充足的能量補充以利于候鳥繼續遷徙順利到達繁殖地或遷徙的終點[28-29],丹頂鶴也如此。針對春遷期丹頂鶴,雙臺河口保護區只是個別幾對丹頂鶴的繁殖地,更是絕大多數丹頂鶴的遷徙停歇地,該時期丹頂鶴選擇該區域停歇的唯一目的是補充體能繼續遷徙,因此能否有效覓食直接關系著丹頂鶴的后續活動,覓食棲息地的選擇也成為有效覓食的關鍵,無干擾或少干擾和高效覓食并攝食將成為丹頂鶴有效覓食補充體能的必要因素。
因子分析的4個因子中,強干擾因子與弱干擾因子與干擾相關,微生境因子和宏生境因子與高效覓食并攝食相關。
關于干擾方面,4個因子中有兩個因子與此相關,可以將二者合并為干擾因子。從數量的角度能看出干擾因子在該時期丹頂鶴覓食棲息地選擇過程中的重要性;從因子貢獻率的角度考慮也能體現出干擾因子的重要性,強干擾因子和弱干擾因子的累積貢獻率為28.45%,超過宏生境因子(22.89%),小于微生境因子(32.73%),占總貢獻率的33.84%。針對干擾因子,丹頂鶴采取回避的措施,但對兩個干擾因子的關注度存在差異,強干擾因子的貢獻率(16.06%)高于弱干擾因子(12.39%),所采取的回避距離也不同,強干擾因子的回避距離(對居民區的回避距離在1 km以上,對油田作業區的回避距離在2 km以上)大于弱干擾因子(對道路的回避距離在300—500 m)。
關于高效覓食并攝食方面,另外兩個因子即宏生境因子和微生境因子與此相關。由因子分析可知,宏生境因子主要關系著該時期覓食棲息地的大尺度選擇,從宏觀的角度判斷某區域是否適合作為覓食棲息地,這是丹頂鶴在空中巡飛尋覓覓食棲息地的大尺度特征,便于識別且具備食物的表征是兩個關鍵的因素。本文的分析結果包括明水面、道路、火燒地。眾所周知,該時期雙臺河口保護區景觀色主要為冰雪的白色,間雜葦草的黃色,而過火區域則為黑色,這樣的區域非常便于環境識別。野外考察發現,過火后的區域,不僅地表積溫高,部分冰雪也融化為冰水且冰層或雪被也變薄,部分區域土壤一定程度解凍,這樣的區域減少或避免了丹頂鶴覓食過程中用于破土、破冰所花費的能量,符合能量最優理論[30-31];而且,類似的區域葦草萌芽早,解凍的明水面也能提供魚蝦等水生高蛋白食物。因此,從某種角度考慮,具備宏生境因子的環境能為丹頂鶴發出該區域有食物的信號。
微生境因子主要關系著該時期丹頂鶴覓食棲息地的小尺度選擇,從微觀的角度判斷某區域是否適合作為具體的覓食點,植被環境的特征有著決定性的作用。本文選取的植被特征為植被高度、植被密度、植被直徑3個因素,從丹頂鶴的喙長和植被蓋度兩個角度對丹頂鶴覓食的便利性進行了分析,結果表明,植被蓋度小(<5%)且植被高度小于丹頂鶴喙長的環境是丹頂鶴首選的覓食點。
4.4 覓食棲息地選擇的尺度
按宏生境和微生境尺度進行模擬,雙臺河口保護區春遷期丹頂鶴覓食棲息地選擇的大生境尺度包括兩個選擇:(1)大的地理分布區的選擇,即雙臺河口保護區(松遼平原亞區遼河平原)為本文的研究區;(2)地理分布區內生境類型的選擇,即以蘆葦沼澤為偏好輔以玉米地、泥灘和草甸。小生境尺度包括兩個選擇:(1)生境類型內覓食區的選擇,該選擇通過宏生境因子(包括明水面、堤壩、火燒地3個環境因素)和干擾因子(包括居民區和油田等強干擾因子和道路等弱干擾因子)來實現,對宏生境因子采取接觸的方式、對干擾因子采取保持一定距離的回避方式;(2)利用棲息地內覓食點的選擇,該層次的選擇通過微生境因子(主要包括植被高度、植被密度、植被直徑等植被因素)來實現,選擇蓋度小(5%)且植被高度小于喙長的微生境。
與扎龍保護區同時期的類似研究結果相比[27],二者的覓食棲息地選擇因素有異同。兩個保護區區內的人為環境存在極大的差異,扎龍保護區內沒有油田建設路,只有村屯間連通的自然路和村村通道路,蘆葦沼澤相對完整,覓食棲息地選擇的影響因素在干擾方面的關注度相對較低,更側重自然因素,如食物因素、隱蔽因素、水因素、標識因素等。二者覓食棲息地選擇的因子雖有差異,但核心因素相似,均直接關系著該時期的主要行為即覓食補充體能。可見,保護區區內人為環境直接影響著丹頂鶴春遷期覓食棲息地的選擇利用。
此外,劉春悅等[16]通過遙感影像以小尺度等級的米為單位對扎龍保護區的丹頂鶴繁殖棲息地特征進行了8個尺度(30—240)的分析,結果表明:植被密度、植被蓋度和長勢、水分條件、人為干擾分別體現在30 m、60—120 m、90 m、>120 m的尺度上。關于丹頂鶴棲息地的多尺度選擇,也有學者采用3S技術以遙感影像為主要數據來源基于不同尺度等級進行了多尺度的分析。其中,曹銘昌等[17-18]以大尺度等級的公頃為單位對黃河三角洲保護區丹頂鶴非繁殖期棲息地選擇及生境適宜性評價進行了分析,范圍從10 hm2至2000 hm2劃分了12個等級,結果表明:丹頂鶴生境需求既體現在小尺度范圍內也體現在大尺度上,在景觀尺度(以翅堿蓬灘涂百分比和水分百分比為主)、斑塊尺度(斑塊類型更重要)和微生境(植被蓋度和水深為限制因子)尺度存在明顯差異。
可以看出,相似研究雖在方法、數據、結果上有所差異,但均表明一點,丹頂鶴的棲息地選擇具有多尺度特征,只不過多尺度表達的方式不同,本文體現的是丹頂鶴由宏觀至微觀的多尺度選擇過程,劉春悅等的研究表明了不同環境因子在小尺度米等級上的多尺度表達,曹銘昌等的研究綜合了多尺度選擇過程與不同環境因子在不同尺度上的表達這兩者的內涵。
基于上述分析,為了有效保護丹頂鶴及其棲息的環境資源,建議雙臺河口保護區在春遷期應更多的關注人為活動的強度和頻率、火燒地的情況。
[1]Su L Y,Zou H F.Status,threats and conservation needs for the continental population of the Red crowned Crane.Chinese Birds,2012,3(3):147-164.
[2]Hu Y M,Shu Y,Li X Z,Wang L,LI Y X,Yang Y C.Change of red crowned crane breeding habitat and the analysis of breeding capacity in Shuangtaihekou national nature reserve.Chinese Journal of Ecology 2004,23(5):7-12.
[3]Li Y X,Song C Z,Yang Y C,Li X J,Li F L,Huang J,Huang X L.Monitoring population dynamics of the migratory Red crowned crane(Grus japonensis)at Shuangtaihekou national nature reserve,Northeastern China,from 1991 to 2012.Chinese Birds,2012,3(3):225-229.
[4]Wan JH,Li M,Zhang J,Ma Y,Liu SW.Study on the changes in wetland landscape pattern of Shuangtai estuary reserve.Geomatics& Spatial Information Technology,2012,35(8):8-12.
[5]Chen A L,Zhu B Q,Chen L D,Wu Y H,Sun R H.Dynamic changes of landscape pattern and eco disturbance degree in Shuangtai estuary wetland of Liaoning province,China.Chinese Journal of Applied Ecology,2010,21(5):1120-1128.
[6]Barbaro L,Couzi L,Bretagnolle V,Nezan J,Vetillard F.Multi-scale habitat selection and foraging ecology of the Eurasian hoopoe(Upupa epops)in pine plantations.Biodiversity and Conservation,2008,17(5):1073-1087.
[7]Grand J,Cushman SA.A multi-scale analysis of species-environment relationships:breeding birds in a pitch pine-scrub oak(Pinusrigida-Quercus ilicifolia)community.Biological Conservation,2003,112(3):307-317.
[8]Sergio F,Pedrini P,Marchesi L.Adaptive selection of foraging and nesting habitat by black kites(Milvus migrans)and its implications for conservation:a multi-scale approach.Biological Conservation,2003,112(3):351-362.
[9]Svardson G.Competition and habitat selection in birds.Oikos,1949,1(2):157-174.
[10]Revermann R,Schmid H,Zbinden N,Spaar R,Schr?der B.Habitat at the mountain tops:how long can Rock Ptarmigan(Lagopusmuta helvetica)survive rapid climate change in the Swiss Alps?A multi-scale approach.Journal of Ornithology,2012,153(3):891-905.
[11]Schipper A M,Koffijberg K,van Weperen M,Atsma G,Ragas A M J,Hendriks A J,Leuven R SE W.The distribution of a threatened migratory bird species in a patchy landscape:a multi-scale analysis.Landscape Ecology,2011,26(3):397-410.
[12]Ciudad C,Robles H,Matthysen E.Postfledging habitat selection of juvenile middle spotted woodpeckers:a multi-scale approach.Ecography,2009,32(4):676-682.
[13]Coreau A,Martin JL.Multi-scale study of bird species distribution and of their response to vegetation change:a Mediterranean example.Landscape Ecology,2007,22(5):747-764.
[14]Kristan W B III.Sources and expectations for hierarchical structure in bird-habitat associations.The Condor,2006,108(1):5-12.
[15]Cushman S A,McGarigal K.Hierarchical analysis of forest bird species-environment relationships in the Oregon Coast Range.Ecological Applications,2004,14(4):1090-1105.
[16]Liu CY,Jiang H X,Zhang SQ,Hou Y Q,Lu J.Breeding habitat characteristics of red-crowned crane at Zhalong of Northeast China:A multiscale approach based on TM and ASAR image data.Chinese Journal of Applied Ecology,2012,23(2):491-498.
[17]Cao M C,Liu GH,Xu H G.A multi-scale analysis of red-crowned crane's habitat selection at the Yellow River Delta Nature Reserve,Shandong,China.Acta Ecologica Sinica,2011,31(21):6344-6352.
[18]Cao M C,Liu G H,Shan K,Hou Y X,Wang M C,Li D L,Shen WM.A multi-scale assessment of habitat suitability of red-crowned crane at the Yellow River Delta Nature Reserve,Shandong,China.Biodiversity Science,2010,18(3):283-291.
[19]Xu J L,Zhang X H,Zhang Z W,Zheng GM,Ruan X F,Zhu JG,Xi B.Multi-scale analysis on wintering habitat selection of Reeves's Pheasant(Syrmaticus reevesii)in Dongzhai National Nature Reserve,Henan Province,China.Acta Ecologica Sinica,2006,26(7):2061-2067.
[20]Zhang G G,Zheng G M,Zhang Z W,Guo J R,Wang J P,Gong SL.Scale-dependent wintering habitat selection by brown-eared pheasant in Luyashan Nature Reserve of Shanxi,China.Acta Ecologica Sinica,2005,25(5):952-957.
[21]Sun M,Zou H F,Jin H Y,Wu Q M,Ma J Z.Environment characters of feeding ground by Red crowned crane during spring stopover in Shuangtaihekou nature reserve,China.Journal of Northeast Forestry University,2012,40(10):108-110.
[22]Wu Q M,Zou H F.Nest site selection pattern of Grus japonensis in Zhalong nature reserve of northeast China.Journal of Forestry Research,2011,22(2):281-288.
[23]Wu Q M,Zou H F.Forage habitat selection of White-naped crane during its incubation period in Zhalong wetland.Chinese Journal of Applied Ecology,2009,20(7):1716-1722.
[24]Zou H F,Wu Q M,Niu M G.Comparing of feeding habitat selection between the wild and semi domestic White-naped crane during the pre breeding period in Zhalong wetland.Chinese Journal of Zoology,2005,40(4):45-50.
[25]Zou H F,Wu QM.Feeding habitat of Red crowned crane and white-napped crane during their courtship period in Zhalong wetland.Chinese Journal of Applied Ecology,2006,17(3):444-449.
[26]Zhu J,Jing K,Gan X J,Ma Z J.Food supply in intertidal area for shorebirds during stopover at Chongming Dongtan,China.Acta Ecologica Sinica,2007,27(6):2149-2159.
[27]Weber L M,Haig S M.Shorebird diet and size selection of nereid polychaetes in South Carolina coastal diked wetlands.Journal of Field Ornithology,1997,68(3):358-366.
[28]Jiang K Y,Wu M,Shao X X.Community composition,seasonal dynamics and interspecific correlation of waterbirds in the Qiantangjiang river estuary and Hangzhou bay.Zoological Research,2011,32(6):631-640.
[29]Ma Z J,Li B,Chen JK.Study on the utilitizaion of stopover sitesand migration strategies of migratory birds.Acta Ecologica Sinica,2005,25(6):1404-1412.
[30]Ma Z J.The research methods and study advances of bird migration.Bulletin of Biology,2009,44(3):5-9.
[31]Ma Z J,Wang Y,Chen J K.Physiological ecology of migratory birds during the stopover periods.Acta Ecologica Sinica,2005,25(11):3067-3075.
參考文獻:
[2]胡遠滿,舒瑩,李秀珍,王凌,李玉祥,楊玉成.遼寧雙臺河口自然保護區丹頂鶴繁殖生境變化及其繁殖容量分析.生態學雜志,2004,23(5):7-12.
[4]萬劍華,厲梅,張杰,馬毅,劉善偉.雙臺河口保護區濕地景觀格局變化研究.測繪與空間地理信息,2012,35(8):8-12.
[5]陳愛蓮,朱博勤,陳利頂,吳艷華,孫然好.雙臺河口濕地景觀及生態干擾度的動態變化.應用生態學報,2010,21(5):1120-1128.
[16]劉春悅,江紅星,張樹清,侯韻秋,陸軍.基于TM與ASAR遙感數據的扎龍丹頂鶴繁殖棲息地多尺度特征.應用生態學報,2012,23(2):491-498.
[17]曹銘昌,劉高煥,徐海根.丹頂鶴多尺度生境選擇機制——以黃河三角洲自然保護區為例.生態學報,2011,31(21):6344-6352.
[18]曹銘昌,劉高煥,單凱,侯銀蓄,王明春,李東來,申文明.基于多尺度的丹頂鶴生境適宜性評價——以黃河三角洲自然保護區為例.生物多樣性,2010,18(3):283-291.
[20]張國鋼,鄭光美,張正旺,郭建榮,王建平,宮樹龍.山西蘆芽山褐馬雞越冬棲息地選擇的多尺度研究.生態學報,2005,25(5):952-957.
[21]孫猛,鄒紅菲,金洪陽,吳慶明,馬建章.雙臺河口保護區丹頂鶴春遷期覓食地環境特征.東北林業大學學報,2012,40(10):108-110.
[23]吳慶明,鄒紅菲.扎龍濕地白枕鶴孵化期覓食生境選擇.應用生態學報,2009,20(7):1716-1722.
[24]鄒紅菲,吳慶明,牛茂剛.扎龍濕地野生與散養白枕鶴繁殖前期覓食生境選擇對比分析.動物學雜志,2005,40(4):45-50.
[25]鄒紅菲,吳慶明.扎龍濕地丹頂鶴和白枕鶴求偶期覓食生境對比分析.應用生態學報,2006,17(3):444-449.
[26]朱晶,敬凱,干曉靜,馬志軍.遷徙停歇期鸻鷸類在崇明東灘潮間帶的食物分布.生態學報,2007,27(6):2149-2159.
[29]馬志軍,李博,陳家寬.遷徙鳥類對中途停歇地的利用及遷徙對策.生態學報,2005,25(6):1404-1412.
[30]馬志軍.鳥類遷徙的研究方法和研究進展.生物學通報,2009,44(3):5-9.
[31]馬志軍,王勇,陳家寬.遷徙鳥類中途停歇期的生理生態學研究.生態學報,2005,25(11):3067-3075.
