不同人為干擾下納帕海湖濱濕地植被及土壤退化特征
2013-12-21 05:00:06唐明艷楊永興
生態學報 2013年20期
唐明艷,楊永興
(同濟大學環境科學與工程學院污染控制與資源化國家重點實驗室,長江水環境教育部重點實驗室,上海 200092)
濕地是最具生產力的生態系統類型之一[1],但其水文依賴性也決定了其生態脆弱性[2]。近幾十年來,隨著人為干擾加劇,世界范圍內濕地發生不同程度退化甚至消失,濕地退化已經成為全球性現象,濕地及其自然資源持續減少[3],開展人為干擾下濕地生態系統退化過程與退化機理研究,已成為當前國際濕地科學研究熱點和前沿問題[4]。以往針對單一人為干擾方式下濕地結構與功能退化特征已有大量報道,其干擾方式涉及放牧[5]、圍墾、生物入侵、環境污染[6]等方面,其研究范圍涵蓋濕地水文[7]、土壤[8]和生物類群[9-10]等結構特征以及蓄水調洪、碳循環、降解污染物[11]等功能特征,然而,針對多重人為干擾下濕地退化特征和過程的綜合研究尚少有涉及。任何生態系統退化均是多種干擾因素綜合作用的結果,不同干擾因素之間存在拮抗或協同作用[12],對比和辨識不同人為干擾的作用方式和作用強度,有助于深入理解濕地生態系統退化的關鍵驅動因素及其退化機理問題。
納帕海濕地位于云南西北部高原區域,為喀斯特地貌上發育的高原濕地[13]。盡管與平原濕地和濱海濕地的顯著退化相比,高原濕地保存相對完整。然而,在過去幾年中,納帕海濕地仍受到嚴重人為干擾,尤以家豬拱地、牛羊過度放牧以及人為建造土路引起的濕地水源補給隔斷等干擾最為突出,引起濕地植被分布格局、土壤性質以及生物組成類群均發生顯著變化[14]。為此,本文分析了家豬拱地、牛羊過度放牧和人為隔斷水源補給3種干擾方式下納帕海濕地植被和土壤退化特征、過程及機理,目的在于:1)對比分析3種人為干擾方式下納帕海湖濱濕地植物群落特征、植物多樣性及土壤理化性質;2)引入土壤退化指數,比較3種人為干擾下濕地土壤特征和退化程度;3)確定納帕海湖濱濕地退化過程中的關鍵人為干擾方式和關鍵生態因子,為納帕海湖濱退化濕地生態恢復和科學管理提供參考。
1 研究區域地理位置概況
納帕海濕地地處滇西北橫斷山脈中段,香格里拉縣城西北部8 km處(27°49'—27°55'N,99°37'—99°43'E),海拔1460 m,行政上隸屬于云南省迪慶藏族自治州。2005年云南納帕海濕地被列入《國際重要濕地名錄》,是我國特有的國家Ⅰ級保護珍稀鳥類——黑頸鶴(Grus nigricollis)的保護區及其越冬生境。納帕海湖盆南北長10 km,東西寬2.3—2.4 km,積水面積660 km2,湖面面積31.25 km2。本區地質構造上屬滇西北槽褶皺系,古生界印支槽褶皺帶,中甸劍川巖相帶[15]。地貌為橫斷山系古夷平面的斷陷盆地,發育典型高原喀斯特地貌,納帕海濕地所處區域的氣候屬于亞熱帶西南季風氣候,同時疊加了海拔高度與高原地貌作用,形成冷涼濕潤的高原氣候,該區年均溫度5.4℃,最冷月平均溫度為-3.8℃,最熱月平均溫度為13.2℃,極端低溫-25.4℃,極端高溫24.5℃。年平均降水量828 mm,徑流量390 mm,年平均產水量2.5×108m3。納帕海濕地具有復雜的生物地理成分和高寒植物杉葉藻(Hippuris vulgaris)等水生植物群落。該區主要土壤類型為沼澤土、泥炭土和沼澤化草甸土[15]。
近年來,受家豬拱地、牛羊過度放牧、人為隔斷補給水源等人類活動干擾尤為突出,受人類不合理的社會經濟活動影響,納帕海濕地生態退化嚴重,引起政府部門關注,云南省于2007年將其列為濕地恢復工程(2007—2020年)優先項目[16]。在2011年7月納帕海退化湖濱濕地植被和土壤樣品采集過程中發現,湖濱濕地家豬放牧干擾尤為嚴重,隨處可見牧民放養的藏香豬,在覓食過程中,常呈條狀、帶狀或片狀拱翻土壤,形成深淺、大小不一的土坑,致使草根裸露,下覆泥炭層出露,有機質礦化加強,泥炭分解強烈,養分含量下降。據野外初步估算,家豬拱地破壞的濕地面積約占全部面積的5%左右。
2 研究方法
2.1 研究樣地設置
納帕海濕地距香格里拉縣城較近,周圍土地利用類型主要為農田和居民區,受人為活動干擾相當嚴重。本研究中,選擇3種對納帕海濕地影響最為嚴重的人為干擾方式:隔斷水源補給(人工在湖濱濕地區修建土路,切斷補給湖濱沼澤的湖水)、牛羊過度放牧及家豬拱地作為共同研究對象,3種干擾方式互相疊加,共同影響本區濕地發育。每種干擾方式各設置一條研究樣帶,進行不同人為干擾下納帕海湖濱濕地退化研究。3條樣帶具體描述如下:
3條樣帶均為溝谷濕地,其中樣帶A谷地長約750 m(沿溝谷中線方向),谷口較寬,地表稍有起伏,主要受人為隔斷水源補給干擾,盡管部分樣點存在家豬拱地和牛羊放牧干擾,但影響范圍相當有限。本樣帶共設置6個樣點,編號依次為A1、A2、A3、A4、A5、A6;樣帶B溝谷地勢較緩,主要是牛羊過度放牧干擾,共設置5個樣點,編號依次為B1、B2、B3、B4、B5,其中樣點B1、B2位于典型的牛羊過度放牧區域;樣帶C溝谷寬廣,集中連片分布家豬拱翻地,受家豬干擾最嚴重,共設置7個樣點,編號依次為C1、C2、C3、C4、C5、C6、C7,其中樣點C2、C4位于嚴重的家豬拱地破壞地區,為裸地,濕地退化最為嚴重。3條樣帶均按人為干擾程度遞減方向設置采樣點,最后采樣點為原生濕地。樣地環境特征及干擾方式見表1,其中樣點C2,C4草根層與腐殖質層被家豬擾動形成擾動層,深約15cm,下層為腐殖質層-潛育層。
2.2 植被調查和土壤樣品采集
于2011年7—8月植物生長旺盛期采用樣方法對滇西北高原納帕海湖濱濕地進行野外研究。每個樣點內隨機設置1個1 m×1 m樣方和2個0.5 m×0.5 m小樣方,記錄植物物種組成,確定該梯度上植物群落的優勢物種,測量植物的密度、蓋度、高度等群落特征。在樣方附近同步設置土壤探坑采集土壤樣品,取樣時根據土壤發育特征和人為干擾情況確定探坑深度和取樣層次,即根據土壤發生層分層取樣,每層各取1份樣品用以室內分析化驗。同時,分別用環刀和小鋁盒取表層和亞表層原狀土壤進行土壤物理性質的測定。帶回的土壤樣品在室內陰涼通風處風干,粉碎研磨,過直徑0.15 mm尼龍篩,裝入聚乙烯密封袋貯存待測。
2.3 土壤樣品分析
在實驗室進行土壤容重(BD)、含水率(WC)和毛管孔隙度(P)的測定,采用環刀法測定容重,采用烘箱法測定含水率,采用浸水法測定毛管孔隙度。按中國科學院南京土壤研究所的分析方法[17],采用半微量開氏法測定全氮(TN);采用擴散法測速效氮(AN);采用NaOH熔融-鉬銻抗比色法測定全磷(TP);采用紫外分光光度法測速效磷(AP);采用NaOH熔融-火焰光度法測定全鉀(TK);采用火焰光度法測速效鉀(AK);采用灼燒法測有機質(OM);采用電位法測定土壤pH值。各類樣品分析化驗均采用平行樣分析測試,平行樣之間誤差在標準允許范圍內。

表1 納帕海湖濱濕地樣地特征Table 1 Characteristics of the plots in lakeside wetlands,Napahai
2.4 數據分析
2.4.1 植物群落退化特征分析
植物群落退化特征分析主要包括群落蓋度、高度和密度等數量特征、植物群落水分類型結構特征以及物種多樣性組成特征3方面。在數量特征分析中,群落蓋度為各種分蓋度之和。計算密度時,將0.25 m2小樣方內植株密度擴大4倍,轉換為1m2樣方,然后再參與分析。群落高度未作處理,依據以上3個指標計算物種重要值(IV),IV=(相對密度+相對高度+相對蓋度)/3;在分析群落水分類型組成特征時,按照金振洲在云南省濕地植物研究中定義的植物水分生態類型[18],將納帕海濕地植物劃分為水生植物(Ap)、沼生植物(He)、濕生植物(Hy)和中生植物(Mesophyte,Me)四大類群,分別計算各類群重要值,考察群落水分生態類型組成結構;物種多樣性組成特征計算了樣地內總物種數(St)、單位樣方內物種豐富度(Sq)、Shannon-Wiener指數(H')和 Whittaker指數(βw)[19]。
依據種重要值數據,進行植物群落TWINSPAN聚類分析和植被-土壤的典范對應分析(CCA)。CCA分析中總計包括38種植物和11個土壤變量,以相關性最高的第一、二軸作物種-土壤因子和樣地-土壤因子二維排序圖,反映群落分布和物種組成沿土壤因子梯度變化情況。其中,樣帶C中樣點C2和C4已退化為裸地,不存在地上植被,為極度退化濕地,未納入以上分析。
2.4.2 土壤退化特征分析
由于人為干擾主要影響濕地表層土壤理化性質,導致土壤養分衰減,土質變劣,削弱土壤亞系統生態功能[15],因此,本文僅對不同人為干擾下退化濕地表層土壤理化性質進行了比較分析。依據野外調查和TWINSPAN聚類結果,分析各樣帶土壤退化特征,并采用Pearson相關性分析檢驗濕地退化過程中各土壤理化指標間的相關性,顯著水平為P=0.05或P=0.01。
采用Adejuwon[20]提出的人為干擾下土壤退化指數定量刻畫土壤退化程度,其公式為:

由于原式中變量xi不易解釋,結合本研究實際情況,引用時對原式作等價轉換,改寫為:

式中,SDI為土壤退化指數,xij是第i個樣地第j個土壤理化指標值,xcj是第j個土壤理化指標參照值,n為納入計算的土壤指標數。本研究中樣帶A、C以樣帶A中樣點A5的土壤理化指標值作為參照值,因樣帶C受豬拱干擾,未發現原生濕地,樣點A5為常年淹水地段,發育典型水蔥群落,受直接人為干擾最小,可視為原生濕地;樣帶B以B5為土壤理化指標值作為參照值。計算時土壤pH以及速效磷和速效鉀含量由于受成土母巖性質影響較大,對人為干擾程度指示性差,未納入分析,實際選取的有土壤含水率、容重、孔隙度、有機質、速效氮、全氮、全磷、全鉀總計8個指標,其中土壤容重由于與土壤退化程度呈負相關[5,21],計算時對其作相反數轉換處理,其他未作處理。據此,SDI值是8個土壤指標的綜合計算結果,可較全面反映人為干擾下濕地土壤退化狀況,SDI<0,代表土壤退化程度高于參照點,其值越小,退化越重,SDI>0則反之。
上述分析中,采用BioDiversity pro軟件計算Shannon-Wiener多樣性指數;采用Win TWINS 2.3軟件進行TWINSPAN聚類分析;采用SPSS13.0軟件進行Pearson相關性分析;利用Canoco 4.5進行植被-土壤CCA分析;采用Microsoft Excel 2007對數據進行基本統計分析及Whittaker多樣性指數計算。
3 結果與分析
3.1 納帕海湖濱濕地植被退化特征分析
3.1.1 納帕海湖濱濕地研究樣點TWINSPAN分類
納帕海退化濕地18個樣點可分為5個退化等級(表2)。從聚類結果看,樣帶A的退化梯度為:原生濕地→輕度退化→中度退化→重度退化;樣帶B的退化梯度為:原生濕地→輕度退化→重度退化,無中度退化,由輕度退化直接跳躍到重度退化;樣帶C的退化梯度為:重度退化→極度退化,這與野外調查結果相一致。總體上,3條樣帶退化程度為:樣帶C>樣帶B>樣帶A。

表2 納帕海湖濱退化濕地分類與分級Table 2 Classification and grade of degraded lakeside wetlands in Napahai
3.1.2 不同人為干擾下納帕海湖濱濕地植物群落組成結構特征變化
納帕海濕地樣帶A包括6種群落類型,分別是水蔥群落(Com.Scirpus validus)、劉氏荸薺群落(Com.Eleocharis liouana)、小花燈心草群落(Com.Juncus articulatus)、華扁穗草群落(Com.Blysmus sinocompressus)、云生毛茛群落(Com.Ranunculus longicaulis)和木里薹草群落(Com.Carex muliensis)。主要伴生種有三穗薹草(Carex tristachya)、稗草(Echinochloa crusgalli)、水蓼(Polygonum hydropiper)、睡菜(Menyanthes trifoliate)和竹葉眼子菜(Potamogeton malaianus)等29種植物,隸屬于17科25屬。樣帶B包括5種群落類型,分別是三穗薹草群落、車前群落(Com.Plantago asiatica)、華扁穗草群落、木里薹草群落和睡菜群落。主要伴生種有云生毛茛、鵝絨委陵菜(Potentilla anseriana)、細葉苦菜(Ixeris chinensis var.intemdia)、高原毛茛 (Ranunculus tanguticus)、小花燈心草、杉葉藻、黑三棱 (Rhizoma scirpi)、水蔥和蔥狀燈心草(Juncus allioide s)等23種植物,隸屬于14科19屬。樣帶C包括4種群落類型,分別是木里薹草群落、鵝絨委陵菜群落、華扁穗草群落和水蓼群落。主要伴生種有白花碎米薺(Cardamine leucantha)、三穗薹草、發草 (Deschampsia caespitosa)、遏藍菜(Thaspi arvense)、竹葉眼子菜和黑藻(Hydrilla verticillata)等16種植物,隸屬于10科14屬。可見,在不同人為干擾下,植物群落類型以及群落組成結構均發生相應改變,在群落類型多樣性上,樣帶A>樣帶B>樣帶C,這與樣帶A常年淹水,受人為干擾程度相對最輕,樣帶B受過度放牧影響,草甸發育廣泛,而樣帶C受家豬拱地干擾,地表干燥緊實,大面積表土裸露密切相關。
3條樣帶植物群落蓋度差異不明顯(圖1),而植物群落高度差異明顯,其中樣帶C群落高度明顯低于樣帶A和樣帶B;植物群落密度以樣帶B最低,沿退化梯度(與采樣順序相反)群落密度整體上呈遞增趨勢(圖1)。3條樣帶均以沼生植物最多,但樣帶C各水分類群重要值明顯偏低,尤其水生植物發育明顯較樣帶A和樣帶B差,而樣帶B中生植物明顯較樣帶A和樣帶C多,比較而言,樣帶A水分生態類型組成最為合理。從物種多樣性變化看,總物種數、物種豐富度和Shannon-Wiener指數以樣帶C最低,以樣帶A最高,Whittaker指數則以樣帶C最高。綜合以上結果,納帕海湖濱濕地植物群落退化程度為:樣帶C>樣帶B>樣帶A(表3)。

圖1 納帕海湖濱濕地植被退化數量特征Fig.1 Quantitative characteristics of vegetation degradation in lakeside wetlands,Napahai

表3 納帕海湖濱濕地植物群落水分生態類型及物種多樣性組成特征Table 3 Composition characteristics of water ecological types and plant species diversity in plant communities in lakeside wetlands,Napahai
3.2 不同人為干擾下納帕海湖濱濕地土壤理化性質變化特征
整體而言,樣帶A和樣帶B土壤含水率隨采樣順序均呈增加趨勢(圖2),樣帶A和樣帶B含水率均明顯高于樣帶C,而土壤容重變化趨勢與含水率完全相反,以樣帶C最高,在1.0—1.3 g/cm3之間波動;3條樣帶土壤毛管孔隙度變化總體上均略呈增加趨勢;土壤pH值受人為干擾影響明顯,3條樣帶土壤pH總體上均呈增加趨勢,但變化格局略有不同,其中樣帶A土壤pH在樣點A1、A2、A3、A4走勢平緩,而在樣點A5、A6處明顯增加,這與樣點A5、A6土壤存在大量小螺殼有關,樣帶B則表現為在樣點B3、B4、B5處明顯增加,樣帶C表現為在樣點C6、C7處明顯增加,這與該5處樣點大量牲畜排泄物帶入的氨氮有關;土壤表層有機質及全氮含量變化規律一致,3條樣帶間分布規律為樣帶A>樣帶B>樣帶C,而土壤全鉀含量在3條樣帶間變化趨勢與全氮變化趨勢完全相反;土壤全磷含量在樣帶A中呈單調降低變化格局,在樣帶B中呈U型變化格局,而在樣帶C中波動性較大,變化趨勢不明顯;土壤速效性養分在3條樣帶中均呈較大波動性,其中土壤速效氮含量在3條樣帶間總體表現為樣帶A>樣帶B>樣帶C,而速效磷和速效鉀波動相當大,且在3條樣帶間亦無明顯差異,暗示可能存在其他關鍵生態變量影響土壤速效性養分含量(圖2)。
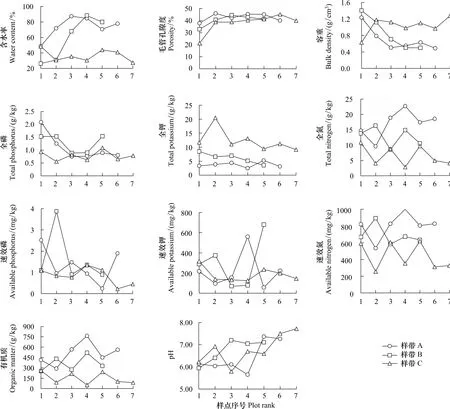
圖2 納帕海湖濱濕地土壤理化性質動態Fig.2 Dynamics of the soil physical and chemical characteristics of lakeside wetlands,Napahai
3.3 不同人為干擾下納帕海湖濱濕地土壤理化指標相關性變化
Pearson相關性分析表明,3條樣帶各土壤指標之間的相關性質及相關強度差異較大,這種相關性變化與人為干擾方式有關。如土壤含水率和容重在樣帶A和樣帶B中均呈極顯著負相關(P<0.01),而在樣帶C中無顯著相關性(P>0.05),表明家豬拱地對土壤物理性質影響較大,又如土壤全氮與有機質在樣帶A和樣帶C中均呈極顯著正相關(P<0.01),而在樣帶B中無顯著相關性(P>0.05),表明牛羊放牧對土壤氮含量和有機質含量影響較大;在相關性質變化上,在樣帶A和樣帶B土壤孔隙度與含水率表現為正相關,與容重為負相關,而在樣帶C中相關性質完全相反,該結果同樣表明家豬拱地對土壤物理性質的影響。因此,土壤指標間相關性質和相關強度的變化,體現出不同干擾方式對濕地土壤的作用特點(表4)。

表4 不同人為干擾下納帕海湖濱退化濕地土壤指標相關性分析Table 4 Pearson correlation analysis for soil indices under different human disturbance in lakeside wetlands,Napahai
3.4 納帕海湖濱濕地植物群落與土壤理化性質的關系
物種與土壤環境的CCA排序統計結果表明,第一、二排序軸特征值分別是0.811和0.613;物種與土壤環境因子顯著正相關,且相關系數均在0.950以上,表明環境變量能很好地解釋植物群落結構變化(表5)。

表5 納帕海湖濱濕地植物群落CCA排序統計結果Table 5 Summary statistics results for the four CCA ordinations in lakeside wetlands plant communities,Napahai
物種排序圖主要反映不同植物種類對土壤條件的相似適應性以及植物群落在土壤理化因子梯度上的分布,通過土壤因子與排序軸的相關分析可以找出與排序軸顯著相關的因子,這些因子就是影響群落分布格局的主要因素。從排序結果看,土壤含水率和容重與第一軸相關性最大,其中含水率與之呈正相關,容重與之呈負相關,土壤全磷含量與第二軸相關性最大,表明土壤含水率和全磷含量是影響納帕海湖濱濕地植物群落種類組成和分布的關鍵因素(圖3)。
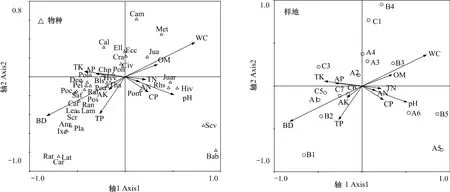
圖3 納帕海湖濱濕地物種、樣地與土壤因子的CCA排序圖Fig.3 CCA biplots between plant species and plots and soil factors in lakeside wetlands,Napahai
環境梯度和人為干擾可以顯著影響植物生態分布[22],排序圖中各植物物種的空間聚集性反映其在土壤環境梯度上的分布情況。沿排序圖逆時針方向,植物水分生態類型呈明顯旱化趨勢,水毛茛(Batrachium bungei)、水蔥、睡菜、黑三棱等水生植物分布在第一象限和第四象限,說明這些種類更適應含水率高的土壤;鵝絨委陵菜、云生毛茛、高原毛茛等中生植物分布在第三象限,說明它們更適應土壤含水率低,全磷、全鉀含量高的區域(圖3)。在群落分布上,原生濕地樣點主要分布在第四象限,輕度退化濕地樣點主要分布在第一、四象限,中度退化樣點主要分布在第二軸附近,重度退化濕地樣點主要分布在第二、三象限,即沿逆時針方向,濕地退化程度逐漸增加,該結果與TWINSPAN分類結果一致(圖3)。
3.5 不同人為干擾下納帕海湖濱濕地土壤退化程度定量分析
土壤退化指數較好地定量反映出3條樣帶土壤退化狀況。在每條樣帶內,隨退化程度增加,土壤退化指數均表現為增加趨勢;不同樣帶間比較表明,樣帶C土壤退化程度最重,總土壤退化指數達-45.01%,樣帶A最輕,總土壤退化指數為-7.40%,二者相差6倍之多,反映出家豬拱地導致納帕海湖濱濕地土壤發生嚴重退化(表6)。

表6 納帕海湖濱濕地不同退化階段土壤退化指數變化Table 6 Changes of soil degradation index in different degradation stage in lakeside wetlands,Napahai
4 討論
4.1 不同人為干擾下納帕海湖濱濕地植被退化特征
濕地植物群落是濕地健康良性發展的基礎,也是濕地監測應用最廣泛的生物指標[23]。濕地退化結果之一是加速水生植物群落向陸生植物群落演替過程。在此過程中,中生植物侵入,增加群落密度和物種組成多樣性,在本研究和其他濕地區的相關研究已證實該結果[24-27]。然而,在不同人為干擾下,這一過程發生程度會有所不同。本研究發現,在3種人為干擾方式中,隔斷水源補給干擾樣帶水分生態類型組成以水生和沼生種類重要值最高,中生植物重要值最低,牛羊放牧干擾樣帶也具有較多的水生和沼生種類,但比較而言,中生植物明顯增加。而家豬拱地干擾樣帶各類群重要值均明顯降低,尤其水生植物降低明顯,受影響最大,這有以下幾個原因:隔斷水源補給干擾樣帶和牛羊放牧干擾樣帶均存在一定比例的積水區,生長水蔥、竹葉眼子菜、睡菜等植物,因此,兩個樣帶的水生、沼生植物均較多,但在沼澤化草甸區,樣帶A受干擾程度小,仍以沼生、濕生植物為主,而樣帶B在過度放牧干擾下,沼澤化草甸的旱生化過程加快[5],極大增加中生植物的多度和分布。家豬拱地主要發生在沼澤化草甸和草甸區域,取食植物地下根莖,尤其殘留的水生、沼生植物根莖,含水量大,質地鮮嫩,尤其喜食,其結果之一就是影響水生、沼生植物種群的更新能力,使得原來有可能自然恢復的水生、沼生植被徹底喪失。因此,納帕海湖濱濕地植物群落退化特征既遵循一般性退化規律,又在具體人為干擾下有其自身退化特征。
納帕海濕地退化是在自然因素和人為因素共同作用的結果,巖溶地貌發育的落水洞與溶洞使湖水外泄,構成沼澤疏干退化自然背景,人為排水、家豬拱地、牛羊過度放牧和無序旅游等人類活動的強烈干預使得自然因素提供的可能性成為現實。在不同人為干擾下,濕地植物群落物種組成、結構向旱生化方向演替,導致群落功能發生退化[28]。根據3種人為干擾下群落高度、密度、蓋度、植物組成多樣性以及群落水分生態類型結構變化。本文初步認為:對納帕海濕地植物群落結構和功能影響最大的是家豬拱地干擾,但是植物分布格局、多度和豐富度與不同時空尺度上的競爭、環境因素、干擾(天然的或人為的)諸因素有關,如何定量刻畫諸因素是未來深入揭示濕地植被退化機理的關鍵[26],有待深入研究。
4.2 不同人為干擾下納帕海湖濱濕地土壤退化特征
濕地土壤理化特征能較好反映人為干擾情況[29],研究不同干擾方式下退化濕地土壤的理化性質變化,不僅有助于我們了解納帕海湖濱濕地退化的真正原因,還為進一步保護濕地生態系統提供了理論依據。
納帕海湖濱濕地土壤在不同人為干擾下其退化特征不同。土壤含水率和毛管孔隙度在3種干擾方式間退化規律為樣帶C<樣帶B<樣帶A,土壤容重為樣帶C>樣帶B>樣帶A,因此,3個指標均受家豬拱地干擾影響最大而受人為隔斷水源補給影響最小。這與樣帶A地表積水面積大、土壤受家畜直接干擾程度小,樣帶B受大型家畜采食踐踏干擾,而樣帶C大面積土壤被拱翻,植被覆蓋率低甚至形成裸地,土壤結構變緊實的干擾特征相一致。其變化機理為:隨著人為干擾強度增加,土壤壓實,土壤顆粒緊密,單位體積內的土壤固相組分增加,而土壤總孔隙度減少,土壤間隙中的毛管孔隙也隨之減少。同時由于容重的增大,導致土壤滲透性能減弱,水分滲入率變小,從而自然含水率也隨之減少。
土壤有機質和全氮含量在3種干擾方式間退化規律均為樣帶C<樣帶B<樣帶A。自然厭氧條件下,土壤有機質礦化率低,有利于有機質積累和保存。在人為干擾下,土壤淹水狀態下的厭氧條件發生改變,通氣性改善,加快原生沼澤積累的有機質分解過程,人為干擾程度越重,有機質分解程度越大。土壤中的氮素很大部分儲藏在土壤有機質的有機態含氮化合物中,全氮含量主要取決于有機質的積累和分解作用的相對強度,因此土壤全氮含量的消長規律與土壤有機質含量的變化相一致。在人為干擾下,土壤全氮含量同樣表現出隨土壤通氣性增加而衰減的變化規律。因此,土壤有機質和全氮含量的變化直接反映出人為干擾下濕地土壤退化過程[15]。土壤全磷含量退化規律為樣帶C<樣帶A<樣帶B。這是由于樣帶A淹水程度重,土壤還原作用較強,加之石灰巖母質導致的較高pH值,使原來固定的磷得以釋放,致使土壤全磷下降,從而略低于過度放牧干擾的樣帶B。但土壤全鉀含量變化規律與全氮完全相反,這是由于土壤中的氮磷主要是以有機態存在,而鉀是以無機態存于礦物中,極難被植物吸收利用,從而土壤中積累較高的濃度。綜合土壤有機質、全氮、全磷和全鉀含量在3條樣帶間的退化規律看,家豬拱地干擾對納帕海濕地土壤影響最大,這從土壤退化指數的定量分析中也可以得到佐證。但本研究中,土壤速效性養分含量無論在樣帶內還是樣帶間均無明顯變化趨勢,由于土壤速效性養分含量與全量養分含量、植物群落類型、土壤微生物組成、環境因子等因素密切相關,推測可能存在本研究未涉及的其他關鍵生態變量,關于其變化機制有待于專門設計土壤學控制實驗進行更嚴謹的解釋。
4.3 濕地植物群落與土壤因子之間的關系
植物群落種類組成和分布受植物區系和地貌、氣候、水文等環境因子的影響,而在小尺度上,土壤性質是植物群落類型的最終決定因子[30]。CCA分析結果表明,納帕海退化湖濱濕地群落物種組成和分布主要受土壤含水率和全磷含量影響。濕生植物主要分布在水分條件好、有機質豐富和全磷含量較低的生境,而中生植物分布在土壤全磷含量高的生境,表明土壤磷含量增加會提高中生雜類草的競爭能力[31]。土壤含水率、容重、有機質、全磷含量差異是形成納帕海湖濱濕地植物分布格局的主要因素,這與若爾蓋高原的研究結果一致[5]。
4.4 人為干擾下納帕海湖濱濕地退化機制
人為干擾導致濕地生態系統退化是當前濕地科學熱點研究領域。納帕海濕地作為《國際重要濕地名錄》收錄的國際濕地,其嚴重的退化現狀已引起各界關注。濕地區牛羊過度放牧、放養野香豬、人為建造土路等人為干擾無不對濕地發育產生深刻影響[32]。本研究也證實家豬拱地是本區最嚴重的干擾方式之一,但生態系統退化絕非單一因素作用的結果,在當前3種干擾方式中,人為隔斷水源補給對濕地發育的影響是漸變、長遠的,其結果加速濕地旱生化過程,向沼澤化草甸方向演替,牛羊過度放牧的踐踏、采食作用下,表層土壤有機質分解加速,耐牧種類得以保留,濕地生境進一步向草甸化演替,而在草甸化濕地,家豬的拱土采食破壞植物根系以及根莖、塊莖等繁殖體,導致亞表層土壤裸露,影響水生和沼生植物群落的自然恢復。因此,3種干擾方式中,人為隔斷水源補給影響面積大,作用強度緩和,而牛羊放牧和家豬拱地作用面積小,但強度大,在3種干擾協同作用下,原生濕地徹底退化為中生草甸甚至次生裸地。因此,在濕地恢復過程中,應依據不同干擾方式制定相應對策,但應首先加強牛羊、豬等大型家畜的科學管理,減緩對濕地的進一步破壞,是當務之急,而對于人為隔斷水源補給需要系統規劃,從長遠著手,深入研究沼澤補水技術和水資源管理技術體系,既滿足農業灌溉、水利工程用水需要,又滿足沼澤發育所需的水文條件,是實現納帕海濕地資源可持續利用的途徑之一。
5 結論
(1)經過比較分析,3種干擾方式下納帕海湖濱濕地植物群落類型組成、物種豐富度、沼生植物重要值、物種數、Shannon-Wiener多樣性指數和土壤有機質、全氮、含水率、速效氮、孔隙度變化規律為:人為隔斷水源>牛羊過度放牧>家豬拱地,而土壤全鉀、容重變化規律則完全相反,但土壤速效磷、速效鉀含量變化規律不明顯。
(2)3條樣帶的各土壤指標間相關強度在不同干擾樣帶間波動較大,甚至相關性質改變,體現出不同干擾方式對濕地土壤的作用特點。
(3)應用CCA分析對土壤理化因子和植被重要值數據進行排序,初步揭示了植物群落、植物物種對土壤理化性質梯度變化的響應情況,納帕海湖濱濕地植被特征與土壤理化性質表現出顯著的相關性,退化濕地植被群落主要受到土壤含水率、全磷含量的影響。
(4)本研究選取的3種干擾方式,通過TWINSPAN分類法、植物群落特征變化分析、土壤理化性質變化分析和土壤退化指數,揭示人為干擾類型與強度的差異對濕地植物群落特征、土壤理化性質特征及其相互關系具有不同的生態作用,當前對納帕海湖濱濕地退化是3種干擾方式協同作用的結果,其影響程度順序為:家豬拱地>牛羊過度放牧>人為隔斷水源補給。
[1]Spohn M,Babka B,Giani,L.Changes in soil organic matter quality during sea-influenced marsh soil development at the North Sea coast.Catena,2013,107:110-117.
[2]Amores M J,Verones F,Raptis C,Juraske R,Pfister S,Stoessel F,Anton A,Castells F,Hellweg S.Biodiversity Impacts from Salinity Increase in a Coastal Wetland.Environmental science & technology,2013,47(12):6384-6392.
[3]Keddy PA,Fraser L H,Solomeshch A I,Junk W J,Campbell D R,Arroyo M T K,Alho CJR.Wetland wonderful:the world's largest wetlands are conservation priorities.Bioscience,2009,59(1):39-51.
[4]Hang D Y,Yang Y X,Yang Y,Li K.Recent advances in wetland degradation research.Acta Ecologica Sinica,2012,32(4):1293-1307.
[5]Li K,Yang Y X,Yang Y,Hang D Y.Characteristics and influence factors of the swamp degradation under the stress of grazing in the Zoige Plateau.Acta Ecologica Sinica,2011,31(20):5956-5969.
[6]Yang Y X.Ecological environment deterioration,mire degeneration and their formation mechanism in the Zoige Plateau.Journal of Mountain Science,1999,17(4):318-323.
[7]Woo M K,Young K L.Hydrogen morphology of patchy wetlands in the high Arctic,polar desert environment.Wetlands,2003,23(2):291-309.
[8]Sigua G C,Griffin J,Kang W J,Coleman S W.Wetland conversion to beef cattle pasture:changes in soil properties.Journal of Soils and Sediments,2004,4(1):4-10.
[9]Lehtinen R M,Galatowitsch SM,Tester JR.Consequences of habitat loss and fragmentation for wetland amphibian assemblages.Wetlands,1999,19(1):1-12.
[10]Seilheimer T S,Wei A H,Chow-Fraser P,Eyles N.Impact of urbanization on the water quality,fish habitat,and fish community of a Lake Ontario marsh,Frenchman's Bay.Urban Ecosystem,2007,10(3):299-319.
[11]Brito E M S,Duran R,Guyoneaud R.A case study of in situ contamination in a mangrove swamp(Rio De Janeiro,Brazil).Marine Pollution Bulletin,2009,58:418-423.
[12]Chen L D,Fu B J.Ecological signif icance,characteristics and types of disturbance.Acta Ecologica Sinica,2000,20(4):581-586.
[13]Tian K,Mo J F,Lu M,Chang FL,Yang Y X.Human disturbances on the ecological environment degradation of Napahai Wetland in the upstream of Yangtze River.Resources and Environment in the Yangtze Basin,2004,13(3):292-295.
[14]Xiao D R,Tian K,Zhang L Q.Relationship between plant diversity and soil fertility in Napahai wetland of Northwestern Yunnan Plateau.Acta Ecologica Sinica,2008,28(7):3116-3124.
[15]Tian K.Mechanism and process of soil degradation in Napahai wetland on Yunnan Plateau[D].Changchun:Northeast Institute of Geography and Agroecology,Chinese Academy of Sciences,2004.
[16]Hu JM,Dong Y X,Yuan H,Li J,Ma B B.Variation of soil nitrogen componentsunder different degradation condition in Napahai wetland reserve.Chinese Journal of Soil Science,2012,43(3):690-695.
[17]The Institute of Soil Science,CAS.Analysis of soil physical and chemical characteristics.Shanghai:Shanghai scientific & Technical Publishers,1978.
[18]Jin Z Z.Complication on systematic,geographical and ecological features of wetland plants in Yunnan Plateau.Beijing:Science Press,2009.
[19]Hang D Y,Yang Y X,Yang Y.Changes of plant species diversity and interspecific correlation in a degraded swamp community along drainage gradients on the ZoigêPlateau of China.Chinese Journal of Plant Ecology,2012,36(5):411-419.
[20]Adejuwon JO,Ekanade O.A comparison of soil properties under different land use types in a part of the Nigerian Cocoa Belt.Catena,1988,15(3/4):319-331.
[21]Shang W,Yang Y X.Degradation characteristics,patterns,and processes of lakeside wetland in Napahai of northwest Yunnan Plateau,Southwest China.Chinese Journal of Applied Ecology,2012,23(12):3257-3265.
[22]Sharpe P J,Baldwin A H.Wetland plant species richness across estuarine gradients:The role of environmental factors and the mid-domain effect.Aquatic Botany,2013,107:23-32.
[23]Wilson M J,Bayley SE.Use of single versus multiple biotic communities as indicators of biological integrity in northern prairie wetlands.Ecological Indicators,2012,20:187-195.
[24]Moran J,Skeffington SM,Gormally M.The influence of hydrological regime and grazing management on the plant communities of a karst wetland(Skealoghan turlough)in Ireland.Applied Vegetation Science,2008,11(1):13-24.
[25]Capon SJ.Flood variability and spatial variation in plant community composition and structure on a large arid floodplain.Journal of Arid Environ,2005,60(2):283-302.
[26]Tousignant M E,Pellerin S,Brisson J.The relative impact of human disturbances on the vegetation of a large wetland complex.Wetlands,2010,30(2):333-344.
[27]Syrbe R U,Michel E,Walz U.Structural indicators for the assessment of biodiversity and their connection to the richness of avifauna.Ecological Indicators,2013,31(S1):89-98.
[28]Lu X G,Liu X H.Wetland research progresses in China.Scientia geographica sinica,2008,28(3):301-308.
[29]San-JoséJ,Montes R,Buendía C.Response of terrestrial-aquatic palm ecotone(morichal)to anthropogenic disturbance in the Orinoco Lowlands.Folia Geobot,2012,47(2):153-178.
[30]Burnside N G,Joyce CB,Puurmann E,Scott D M.Use of vegetation classification and plant indicators to assess grazing abandonment in Estonian coastal wetlands.Journal of Vegetation Science,2007,18(5):645-654.
[31]King R S,Richardson CJ,Urban D L,Romanowicz E A.Spatial dependency of vegetation environment linkages in an anthropogenically influenced wetland ecosystem.Ecosystems,2004,7(1):75-97.
[32]Reiss K C,Brown M T,Lane C R.Characteristic community structure of Florida's subtropical wetlands:the Florida wetland condition index for depressional marshes,depressional forested and flowing water forested wetlands.Wetlands Ecology and Management,2010,18(5):543-556.
參考文獻:
[4]韓大勇,楊永興,楊楊,李珂.濕地退化研究進展.生態學報,2012,32(4):1293-1307.
[5]李珂,楊永興,楊楊,韓大勇.放牧脅迫下若爾蓋高原沼澤退化特征及其影響因子.生態學報,2011,31(20):5956-5969.
[6]楊永興.若爾蓋高原生態環境惡化與沼澤退化及其形成機制.山地學報,1999,17(4):318-323.
[12]陳利頂,傅伯杰.干擾的類型、特征及其生態學意義.生態學報,2000,20(4):581-586.
[13]田昆,莫劍鋒,陸梅,常鳳來,楊永興.人為活動干擾對納帕海濕地環境影響的研究.長江流域資源與環境,2004,13(3):292-295.
[14]肖德榮,田昆,張利權.滇西北高原納帕海濕地植物多樣性與土壤肥力的關系.生態學報,2008,28(7):3116-3124.
[15]田昆.云南納帕海高原濕地土壤退化過程及驅動機制[D].長春:中國科學院東北地理與農業生態研究所,2004.
[16]胡金明,董云霞,袁寒,李杰,馬彬斌.納帕海濕地不同退化狀態下土壤氮素的分異特征.土壤通報,2012,43(3):690-695.
[17]中國科學院南京土壤研究所.土壤理化分析.上海:上海科學技術出版社,1978.
[18]金振洲.云南高原濕地植物的分類與地理生態特征匯編.北京:科學出版社,2009.
[19]韓大勇,楊永興,楊楊.若爾蓋高原退化沼澤群落植物多樣性及種間相關性沿排水梯度的變化.植物生態學報,2012,36(5):411-419.
[21]尚文,楊永興.滇西北高原納帕海湖濱濕地退化特征、規律與過程.應用生態學報,2012,23(12):3257-3265.
[28]呂憲國,劉曉輝.中國濕地研究進展.地理科學,2008,28(3):301-308.
