松嫩草地兩種廣布豆科植物分布特征及其與土壤因子關系
2014-01-02 06:00:20李強周道瑋宋彥濤
草業學報 2014年1期
關鍵詞:物種
李強,周道瑋 ,宋彥濤
(1.中國科學院東北地理與農業生態研究所,吉林長春130102;2.中國科學院大學,北京100049)
豆科植物在草地植物資源中具有重要地位[1]。豆科植物具有低纖維、高蛋白含量特點,飼喂價值高,一般是優良的牧草[2],許多豆科植物還能為綠肥、蜜源、藥材所用[3]。更重要的,豆科植物根瘤特有的固氮特性能將大氣中的N2轉化為植物可利用的形態,不僅能減少自身生長對土壤氮素的依賴,其固定的氮還能通過根部的分泌、死亡根的分解、動物糞便等途徑轉移給鄰居植物利用[4-5]。有研究報道在豆科與禾本科混播草地中,豆科植物固氮最多可達到草地地上部氮積累總量的46%,這對維持草地系統的生產力起到了重要作用[6]。豆科植物還具有高的枯落物質量,能夠通過枯落物的快速分解加速養分循環,改善土壤養分狀況[7]。土壤狀況的改善可能提供更多的生態位給其他植物,對保護草地的物種多樣性和增加資源互補性是有利的[8]。發達的根系也是豆科植物的普遍特征,許多豆科植物的根系分布可深至5 m,這不僅能夠增加群體的水分利用和養分吸收效率,還能提高植被抗風、固土的作用[9]。鑒于豆科植物在草原系統中的重要價值,保護和利用豆科植物資源對發展草原經濟和生態有重要意義。
保護和發展草原豆科植物資源需要對豆科植物的分布和建植規律有充分的認知,如了解豆科物種的地理分布、豆科物種分布與環境因子關系、豆科分布區的植被特征等都是有效利用豆科植物資源的前提。國外早有學者在局域內對天然草地豆科植物分布特征開展了研究,如Ehrman和Cocks[10]研究了西亞一年生豆科飼草分布的地理、生態特征,Beale等[11]研究了非洲西北部草地一年生豆科植物的分布及其與氣候、土壤因子關系。國內對天然草原中豆科植物分布規律的研究較少,且多集中于豆科資源調查[9,12-13],也有一些研究針對區域自然條件從抗逆生理角度研究了一些豆科物種對干旱、鹽堿等環境的適應能力,間接地預測了這些豆科物種在該區域內分布的可能性和成功性[14-16]。與國外比較,國內對豆科植物分布規律的研究,尤其是天然草地豆科植物分布規律的研究相對薄弱。
松嫩草地位于歐亞草原區東緣,地理和水文條件決定了松嫩草地獨特的土壤特性——鹽堿化[17]。受土壤理化性質限制,以羊草(Leymus chinensis)占絕對優勢的鹽堿化草甸是松嫩草地的主要植被。在羊草草甸中廣泛分布著許多雜類草群落,這些雜類草群落以大小不一的斑塊狀鑲嵌在草甸中,相鄰的雜類草斑塊包裹羊草群落形成雜類草區。豆科植物就生長在這些雜類草區中。這些豆科植物不僅改善了松嫩草地牧草種類單一的局面,且對草地的飼草產量和質量、生態穩定性有極大貢獻[18]。保護和發展這些豆科物種要求我們了解這些豆科植物的分布特性,然而,很少有學者在該區域針對這個問題開展研究。盡管陶巖等[19]研究了該區域5種豆科植物總體的空間分布格局,但該研究并未在群落水平上具體揭示每種豆科植物分布規律,包括其與雜草區群落特征、土壤理化因子的對應關系。而這些信息恰是在天然草地中保護、引種利用豆科植物的關鍵背景。基于此,本研究通過野外調查實驗針對本區內分布最廣、數量最多,且產量和飼喂價值均較高的兩種豆科植物——五脈山黧豆(Lathyrus quinquenervius)和野豌豆(Vicia cracca)進行這些信息的揭示,以為本區域這兩種豆科植物資源利用提供關鍵理論支持。
1 材料與方法
1.1 研究區自然概況
研究地點位于中國科學院長嶺草地農牧生態研究站(44°33'N,123°31'E,海拔145 m),地處松嫩草地腹地。該區屬于溫帶半濕潤半干旱溫帶大陸性季風氣候,春季干旱多風且降水稀少,地表蒸發劇烈;夏季降水集中,冬季降雪較少。年均溫4.9℃,無霜期140~160 d。年降水量300~500 mm,集中在6-9月,占總降水量的70%。年蒸發量1600.2 mm,約是降水量的3.5倍。地帶性土壤為黑鈣土。主要植被類型為羊草草甸。以多年生根莖禾草羊草為優勢的草甸中,鑲嵌分布著一定數量的雜草群落聚集區,包括扁蓿豆(Medicago ruthenica)、興安胡枝子(Lespedeza davurica)、細葉胡枝子(Lespedeza hedysaroides)、五脈山黧豆、野豌豆等在內的豆科植物以零散或聚集的方式生長在這些雜草群落區內。一種豆科植物往往集中分布于特定的雜草群落區(如圖1中A、B、C)。其中,五脈山黧豆和野豌豆是數量最多、分布最廣的兩種豆科植物。
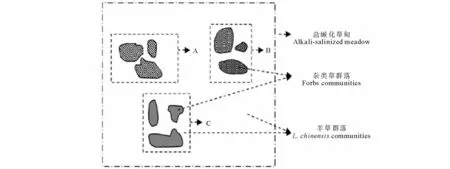
圖1 鹽堿化草甸雜類草群落分布特征Fig.1 The distribution feature of forb communities in alkaline meadow
1.2 研究方法
2009年8月在1個1000 hm2的天然羊草草甸中選取五脈山黧豆和野豌豆分布的典型雜草群落區,根據兩類雜草群落斑塊格局特點,各分別設置一個50 m×100 m的樣帶。每個樣帶均分為100個5 m×5 m的小區,每個小區中央設置一個1 m×1 m的樣方進行植物材料和土壤取樣。查數每種植物密度后,齊地面收獲地上生物量,目標豆科植物單獨收獲。取樣過程中未發現有目標以外其他種豆科植物生長在樣方內。地上植物材料在70℃下烘干48 h后稱取干重。地上部分取樣的同時,采用直徑3 cm的土鉆鉆取0~20 cm深度土壤樣品,對每層土壤,每個樣方內隨機鉆取3鉆混合為一份樣品,土壤樣品帶回室內陰干,待測土壤理化性質。
1.3 土壤樣品分析
每份風干土壤樣品的一部分過2 mm篩后,實驗室內使用PHS-3C pH儀和DDS-307電導率儀(上海,中國)測定土壤pH和電導率(水土質量比5∶1),另一部分過0.25 mm篩,采用重鉻酸鉀氧化法測定土壤有機質,土壤全氮用凱氏定氮儀(Kjeltec 2003 Analyzer Unit,FOSS TECATOR,H?gan?s,Sweden)測定,土壤全磷用鉬銻抗比色法測定[20]。
1.4 數據處理
采用物種豐富度指數(R)、Shannon-Wiener多樣性指數(H)和均勻性指數(E)表示每個樣方的群落結構特征,計算公式如下:

式中,S代表樣方中的物種數目,Pi表示第i個物種個體數目占總個體數目的比例。
在每個豆科分布區內,根據調查樣方內目標豆科植物的地上生物量,將全部樣方進行分組。無目標豆科植物的樣方為一組,考慮到野豌豆和五脈山黧豆個體和群體地上生物量的差異,從>0始,五脈山黧豆地上生物量每增加50 g以內(含50 g)的樣方為一組,野豌豆地上生物量每增加100 g以內(含100 g)的樣方為一組。統計每組內的樣方數及在總樣方中的比例,在每組內,統計目標豆科地上生物量的最大值、最小值、均值。不同組別群落特征和土壤理化性質的差異通過單因素方差分析后多重比較(Duncan)實現,各組豆科地上生物量和群落特征、土壤因子均值間的關系采用一般線性回歸分析,顯著性水平均為0.05。所有統計采用SPSS軟件完成(SPSS 16.0 for windows,SPSS Inc.,Chicago,IL,USA)。采用 Sigma Plot 10.0(Systat software,Inc.,San Jose,CA,USA)完成作圖。
2 結果與分析
2.1 豆科植物生物量分布特征
選定的五脈山黧豆分布區樣帶內,包含目標豆科植物的樣方數為75個,占據了總調查樣方數的3/4(表1)。在含有五脈山黧豆的群落中,五脈山黧豆的最大地上生物量分布為286.25 g/m2,最小地上生物量為51.73 g/m2,未見有其地上生物量低于50 g/m2的樣方。全部的100個樣方按照規定方案分為6組(各組內豆科生物量均值見表1)。其中,五脈山黧豆地上生物量在100~150 g/m2和150~200 g/m2組的樣方數分別為24和21個,明顯高于其他五脈山黧豆分布組。野豌豆的地上生物量分布與五脈山黧豆相似。分布有山野豌豆的樣方數為75個,同樣占據了總調查樣方數的3/4(表1)。在含有野豌豆的群落中,野豌豆的最大地上生物量分布為494.77 g/m2,最小為61.19 g/m2。所有的100個樣方同樣被劃分為6組(各組內豆科地上生物量均值見表1),其中,野豌豆地上生物量在100~200 g/m2和200~300 g/m2組的樣方數分別為27和17個,明顯高于其他野豌豆分布組。
2.2 豆科植物生物量與群落特征關系
兩類雜草群落區中,豆科植物地上生物量和群落高度、植物密度、群落地上生物量之間均有顯著的線性相關關系(P<0.05;相關系數及方程見圖2)。其中,隨著豆科植物地上生物量的增加,群落高度和群落地上生物量持續增加(圖2A、圖2C),而群落的植物密度逐漸降低(圖2B)。五脈山黧豆地上生物量與群落密度、群落地上生物量關系斜率絕對值較野豌豆更大,表明二者隨五脈山黧豆地上生物量增加的變化速率更大。按豆科植物地上生物量由低到高,不同五脈山黧豆地上生物量組對應的群落高度均值范圍為37.8~57.8 cm;植物密度均值范圍為896~507株/m2;群落地上生物量均值范圍為335.03~593.25 g/m2。不同野豌豆地上生物量組對應的群落高度均值范圍為43.2~75.9 cm;植物密度均值范圍為902~526株/m2;群落地上生物量均值范圍為381.95~733.35 g/m2(圖2)。在相鄰低豆科地上生物量組別間,群落高度、植物密度、群落地上生物量差異明顯,尤其是無豆科植物組和相鄰組間,幾乎都呈現顯著性差異,而在高豆科地上生物量組間,這些指標的變化相對較小(圖2)。
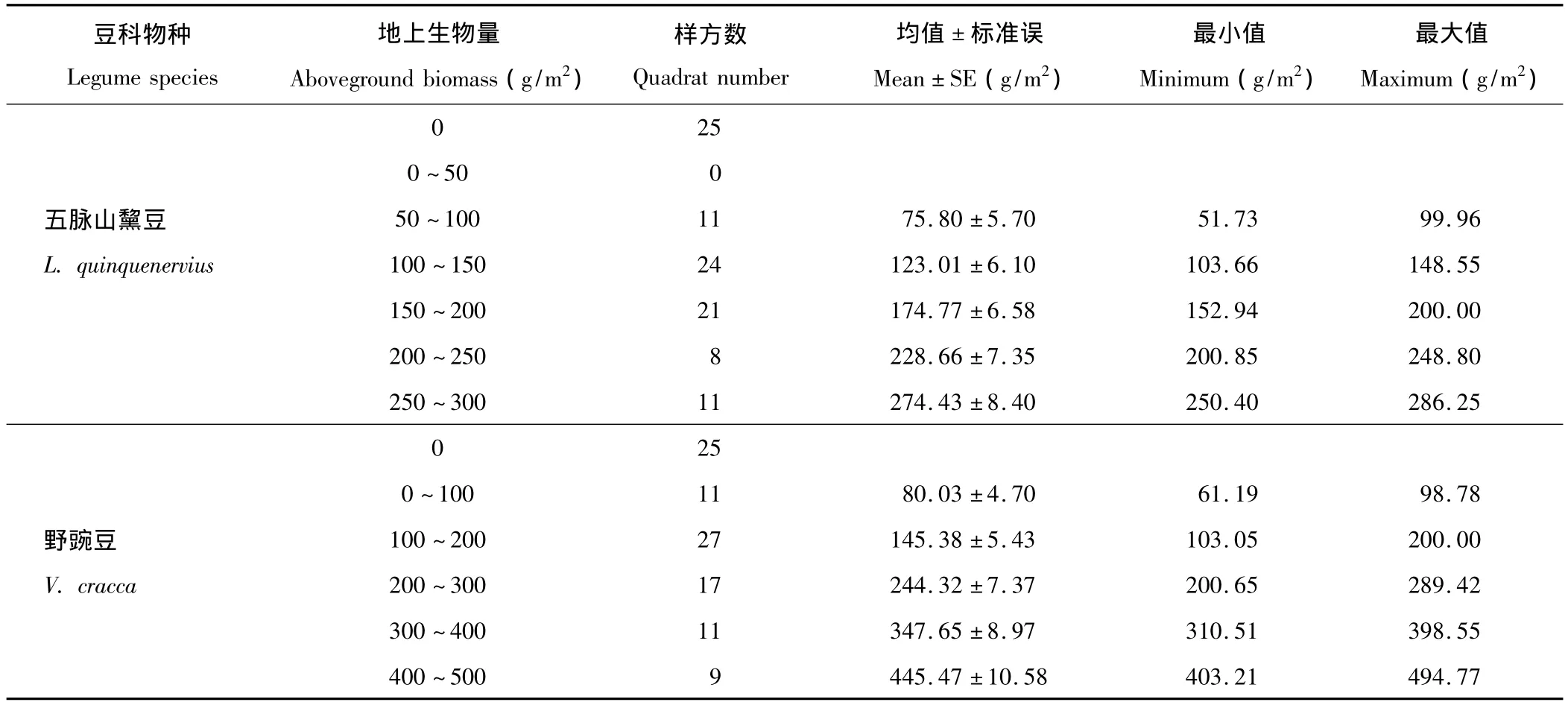
表1 豆科植物地上生物量在雜草群落區中的分布特征Table 1 The distribution feature of aboveground biomass of legumes in forb community area
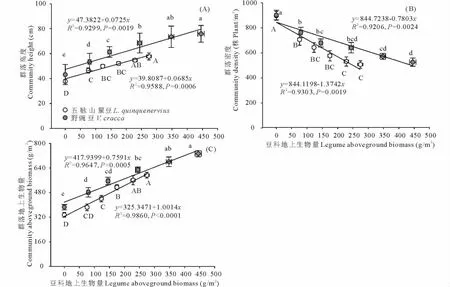
圖2 豆科植物地上生物量與群落高度(A)、密度(B)、總地上生物量(C)的關系Fig.2 The relationships between legumes aboveground biomass and height(A),density(B),aboveground biomass(C)of community
兩類雜草群落區中,群落物種豐富度、多樣性、均勻性和豆科植物地上生物量之間均有顯著的線性正相關關系(P<0.05;相關系數及方程見圖3),并且三者均隨群落中豆科地上生物量的遞增而持續增加。五脈山黧豆地上生物量與三者關系斜率較野豌豆更大,表明3個變量隨五脈山黧豆地上生物量增加的變化更快。同樣的,按豆科植物地上生物量由低到高,不同五脈山黧豆地上生物量組對應的群落物種豐富度指數均值范圍為5.08~11.27;Shannon-Wiener指數均值范圍為0.88~2.36;均勻性指數均值范圍為0.52~0.98。不同野豌豆地上生物量組對應的群落物種豐富度指數均值范圍為4.63~9.89;Shannon-Wiener指數均值范圍為0.68~2.20;均勻性指數均值范圍為0.45~0.96(圖3)。與群落高度等相似,在相鄰低豆科地上生物量組別間,群落物種豐富度、多樣性、均勻性差異明顯,尤其是無豆科植物組和相鄰組間,均呈現顯著性差異(P<0.05),而在高豆科地上生物量組間,這些指標的變化相對較小(圖3)。
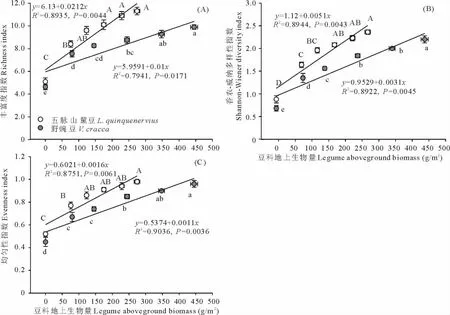
圖3 豆科植物地上生物量與群落物種豐富度(A)、多樣性(B)、均勻性(C)的關系Fig.3 The relationships between legumes aboveground biomass and species richness(A),diversity(B),evenness(C)of community
2.3 豆科植物生物量與土壤因子的關系
兩類雜草群落區中,群落豆科植物地上生物量和土壤有機碳含量、總氮含量、土壤pH、土壤電導率之間均有顯著的線性相關關系(P<0.05;相關系數及方程見圖4)。其中,土壤有機碳含量和總氮含量隨群落豆科植物地上生物量的增加而增加(圖4A、圖4B),而土壤pH和電導率隨群落豆科植物地上生物量增加而降低(圖4D、圖4E)。土壤總磷含量與群落豆科植物生物量間未發現有顯著相關關系(P>0.05;圖4C)。除土壤總磷外,五脈山黧豆地上生物量與其他土壤因子關系斜率絕對值較野豌豆更大,表明這些因子隨五脈山黧豆地上生物量增加的變化更快。按豆科植物地上生物量由低到高,不同五脈山黧豆地上生物量組對應的土壤有機碳含量、土壤總氮含量、土壤 pH、土壤電導率均值范圍為10.33 ~16.44 g/kg、1.36 ~1.99 g/kg、9.77 ~8.56、524 ~232 μS/cm。不同野豌豆地上生物量組對應的土壤有機碳含量、土壤總氮含量、土壤pH、土壤電導率均值范圍為10.08~15.73 g/kg、1.31 ~2.14 g/kg、9.71 ~8.51、516 ~216 μS/cm(圖4)。相鄰低豆科地上生物量組別間,土壤有機碳含量、總氮含量、土壤pH和電導率差異明顯,尤其是無豆科植物組和相鄰組間,均呈現顯著性差異(P<0.05),而在高豆科生物量組間,這些土壤因子的變化相對較小(圖4)。對土壤總磷含量,僅發現野豌豆分布區0豆科生物量組與其他組間的顯著性差異(P<0.05;圖4C)。五脈山黧豆分布區中,0豆科生物量組的土壤總磷含量均值為0.30 g/kg,其他組維持在0.32 g/kg左右;野豌豆分布區中,0豆科生物量組的土壤總磷含量均值為0.29 g/kg,而其他組維持在0.33 g/kg左右。
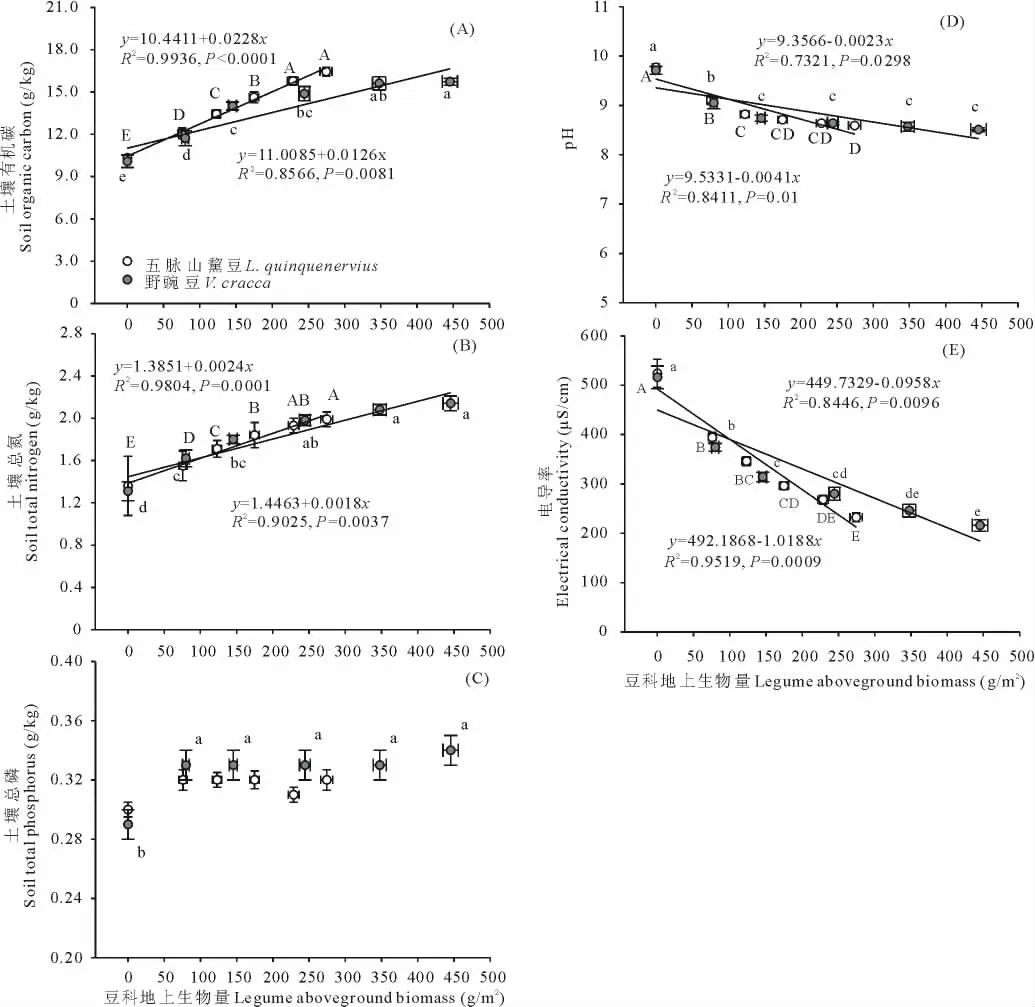
圖4 豆科植物地上生物量與土壤因子的關系Fig.4 The relationships between legumes aboveground biomass and soil factors
3 討論
3.1 五脈山黧豆和野豌豆的分布特點
陶巖等[19]的研究中已表明五脈山黧豆和野豌豆在松嫩草地中表現聚集分布的空間格局,但該研究未具體給出兩種豆科植物種群的聚集程度。而本文通過研究得出兩種豆科植物在單位面積上的分布數量和各數量級所占面積比例。結果表明:五脈山黧豆和野豌豆種群聚集程度主要在100~200 g/m2和100~300 g/m2水平。依據兩種豆科植物在雜草區中被發現的最大單位面積地上生物量,這兩個范圍都處于兩種豆科植物的中等聚集程度上。在兩種豆科植物各自的調查樣帶內,含有目標豆科植物的樣方均為75個,這可能意味著在相同大小的雜草區域內,兩種豆科植物的分布面積相近。種群大小能夠影響物種在群體中占據的生態位,進而影響物種的生存、繁殖以及環境變化下的適應性[21]。我們的研究結果發現,五脈山黧豆和野豌豆的最小單位面積聚集量都高于50 g/m2,分別為51.73和61.19 g/m2,這可能是本區域內兩種豆科植物在群落中維持種群延續的最小單位面積地上生物量。
3.2 兩種豆科植物生物量與群落特征關系
豆科植物地上生物量和群落特征的關系信息能夠為我們調研和保護豆科植物資源、認識豆科植物功能提供直觀的依據。我們的研究結果發現,兩種豆科植物的地上生物量與群落高度、群落地上生物量間有顯著的正相關關系。含野豌豆的群落具有更高的群落高度和更大的群落地上生物量。分析其原因可能包括,1)與兩種豆科植物的性狀有關。兩種豆科植物一般較禾本科植物葉片繁密、多枝、個體地上生物量更大。一方面,豆科植物在群落中數量越多可能增加取樣效應(sampling effect)對群落地上生物量的影響[22-23]。另一方面,豆科植物在群落中存在能夠增加群落冠層的密閉度和光資源的截獲,而其他植物可能通過增加個體高度的方式實現對光資源的競爭,間接增加群體高度和地上生物量[24-25]。尤其野豌豆,不僅分枝更多、個體地上生物量更大,且具有蔓生的特點,能夠依附在其他植物的莖上不斷延伸個體,其主莖長度可達1 m以上,這增加了對光的截獲強度,促使了其他植物群體高度和地上生物量的增加。2)兩種豆科植物存在的群落中往往伴生有牛鞭草(Hemarthria sibirica)、箭頭唐松草(Thalictrum simples)等植物,這些植物本身具有較高的高度,也能通過光競爭間接促進群體生長。3)豆科植物地上生物量和土壤養分呈正相關關系,和土壤鹽堿程度呈負相關關系(圖4),高的土壤肥力和低的鹽堿脅迫可能直接促進豆科和鄰居植物的生長。研究結果顯示,豆科植物地上生物量和群落植物密度之間有顯著負相關關系。這主要是因為在豆科植物數量增加情況下,群落的物種組成發生了明顯變化,原初密度較大的禾草群體不斷被個體更大的豆科植物和雜草物種取代,導致總體密度的下降。
伴隨豆科植物地上生物量不斷增加,群落中禾草的絕對優勢被打破,加之與豆科植物量協同增加的土壤養分,這些因素可能在群落內為更多物種提供生態位[8],導致群落物種豐富度、多樣性指數、均勻性指數的增加。在結果中我們也發現:群落密度、物種豐富度、多樣性和均勻性隨五脈山黧豆生物量增加較隨野豌豆地上生物量增加的變化速率更大,且相同兩種豆科地上生物量下,上述變量在含五脈山黧豆的群落中具有更高值。這表明伴隨五脈山黧豆數量的改變,群落物種替換和結構變化的速率更大。而且,從土壤數據中發現:在低豆科地上生物量下,五脈山黧豆生境的土壤條件并未優于野豌豆。盡管當前的研究還無法斷定其背后的機理,但我們推定單位質量的五脈山黧豆較野豌豆對其他植物建植的促進作用(facilitation effect)更強。
眾所周知土壤本底條件可能直接影響群落特征,包括豆科植物在群落中的數量[10-11],而豆科植物也可通過對土壤的改善作用或資源的修飾作用間接地影響植物的建植和生長[6-8,24-25]。盡管我們對兩種豆科植物地上生物量與雜草群落特征的關系給予了解釋,但據當前野外研究的結果,我們仍無法判定土壤條件和豆科植物二者對雜草群落特征的決定作用,這需要進一步的因子控制實驗來驗證。
3.3 兩種豆科植物生物量與土壤因子關系
我們的研究結果發現,土壤有機碳、總氮、土壤pH、電導率與豆科地上生物量存在顯著相關關系。但土壤條件和豆科植物數量之間相互的決定關系卻很難確定。因為自然生態系統中,土壤和植物存在相互作用,即土壤條件能夠影響植物的定居,而定居的植物又將通過生命活動和物質循環途徑反饋作用于土壤[26],尤其豆科植物,其對土壤條件可能有顯著的改善作用。在鹽堿化草地中,土壤鹽堿程度是制約植物建植的首要因子[27-28],土壤的高度鹽堿化能通過抑制植物種子的萌發和幼苗的生長影響植物的建植[29]。然而,楊光[30]利用兩種豆科植物在堿性鹽脅迫下的發芽研究發現,即使土壤pH達到11.5以上,兩種豆科植物種子的發芽率仍能達到83%以上,而在包括裸堿斑在內的整個松嫩草地,土壤的鹽堿程度都要低于pH=11.5的水平[31]。因此,松嫩草地的土壤鹽堿化狀況可能不會、或至少不會單獨構成兩種豆科植物建植的限制因素。但考慮自然界中環境因子的協同變化,如在研究本區域內,表土鹽堿化程度往往匹配著土壤干旱、養分貧瘠程度,因此并不排除鹽堿脅迫協同其他因子共同限制兩種豆科植物分布。豆科植物本身具有固氮的特性,因此土壤氮不可能成為限制豆科植物分布的因素,也未見有相關的報道。對于土壤磷水平,李強等[32]和宋彥濤[33]在本區域內的研究報道:未退化羊草草地的土壤0~20 cm層總磷含量均低于0.20 g/kg,這個數值要低于本研究中雜草區的最低土壤磷含量,因此土壤磷可能是這兩類豆科植物分布的限制因子之一。但對任意豆科植物分布區,在含豆科植物的各組樣方中,土壤總磷含量并未表現顯著差異,這可能意味著土壤磷對兩種豆科植物的分布限制有個閾值,而超過這個閾值,土壤磷即不構成對兩種豆科植物生長數量的限制。而根據我們的研究結果,如果磷確是兩種豆科植物分布限制因子,這個閾值應該是有五脈山黧豆和野豌豆分布的最小土壤磷水平,分別是0.29和0.27 g/kg。綜上,對本研究結果中土壤有機碳、總氮、土壤pH、電導率隨兩種豆科植物地上生物量增加的變化,可能有兩個解釋:1)盡管這些變量不會或至少不會單獨構成限制兩種豆科植物分布的因素,但它們附和了其他限制因子的變化而變化,譬如土壤水分或其他因子。但同時,兩種豆科植物本身一定程度上也在改善土壤條件。因此,這些土壤因子在不同豆科植物數量的群落中的差異是土壤條件自身的變化和豆科植物改良土壤的共同作用結果;2)本區域內任一土壤因素均不構成對兩種豆科植物分布的限制,兩種豆科植物聚集程度的變化完全取決于其種子散布的隨機性[34],而土壤性質的變化則源于不同數量豆科植物改良土壤作用的差異。盡管土壤性質與兩種豆科植物生物量之間關系的機理還有待更深入的研究,但本文直觀地給出了兩者的對應關系,這對我們保護和發展這兩種豆科植物提供了重要參考。
4 結論
野外調查結果表明,五脈山黧豆和野豌豆種群聚集程度主要在100~200 g/m2和100~300 g/m2水平,五脈山黧豆和野豌豆的最小單位面積聚集量分別為51.73和61.19 g/m2,這可能是兩種豆科植物在群落中維持個體生存和種群延續的最小單位面積地上生物量。群落高度、地上生物量、物種多樣性隨兩種豆科植物地上生物量的增加而增加,這可能與兩種豆科植物的性狀和促進效應有關。五脈山黧豆存在下,群落物種替換和結構變化的速率更大。結合土壤數據,我們推定單位質量的五脈山黧豆較野豌豆對其他植物建植的促進作用更強。土壤有機碳、總氮、土壤pH、電導率與兩種豆科植物地上生物量存在顯著相關關系。而參考以往研究,我們得出這些土壤因子不會或至少不會單獨構成對兩種豆科植物分布的限制。磷可能是豆科植物分布的限制因子之一。因此,本研究中涉及的土壤因子隨豆科植物數量的變化可能是協同了其他豆科植物分布限制因子的變化而出現的,或由豆科植物對土壤的改良作用決定的。
[1]牛書麗,蔣高明.豆科植物在中國草原生態系統中的地位及其生理生態研究[J].植物學通報,2004,21(1):9-18.
[2]Buxton D R,Redfearn D D.Plant limitations to fiber digestion and utilization[J].The Journal of Nutrition,1997,127(5):814-818.
[3]賈麗,曲式曾.豆科錦雞兒屬植物研究進展[J].植物研究,2001,21(4):515-518.
[4]Lory J A,Russelle M P,Heichel G H.Quantification of symbiotically fixed nitrogen in soil surrounding alfalfa roots and nodules[J].Agronomy Journal,1992,84(6):1033-1040.
[5]Paynel F,Murray P J,Cliquet J B.Root exudates:a pathway for short-term N transfer from clover and ryegrass[J].Plant and Soil,2001,229(2):235-243.
[6]Cadisch G R,Schunke M,Giller K Z.Nitrogen cycle in monoculture grassland and Legume-grass mixture in Brazil Red soil[J].Trop Grasslands,1994,28:43-52.
[7]Spehn E M,Scherer-Lorenzen M,Schmid B,et al.The role of legumes as a component of biodiversity in a cross-European study of grassland biomass nitrogen[J].Oikos,2002,98(2):205-218.
[8]Van Ruijven J,Berendse F.Positive effects of plant species diversity on productivity in the absence of legumes[J].Ecology Letters,2003,6(3):170-175.
[9]譚志遠.陜西及甘,寧部分地區豆科植物根瘤菌資源調查[J].西北植物學報,1996,15(2):189-196.
[10]Ehrman T,Cocks P S.Ecogeography of annual legumes in Syria:distribution patterns[J].Journal of Applied Ecology,1990,27(2):578-591.
[11]Beale P E,Lahlou A,Bounejmate M.Distribution of wild annual legume species in Morocco and relationship with soil and climatic factors[J].Australian Journal of Agricultural Research,1991,42(7):1217-1230.
[12]王一峰,楊文璽,王春霞,等.甘肅豆科飼用植物資源[J].草業科學,2006,23(3):12-16.
[13]劉國道,羅麗娟,白昌軍,等.海南豆科飼用植物資源及營養價值評價[J].草地學報,2006,14(3):254-260.
[14]李鴻雁,李志勇,師文貴,等.內蒙古扁蓿豆葉片解剖性狀與抗旱性的研究[J].草業學報,2012,21(3):138-146.
[15]劉晶,才華,劉瑩,等.兩種紫花苜蓿苗期耐鹽生理特性的初步研究及耐鹽性比較[J].草業學報,2013,22(2):250-256.
[16]陸嘉惠,呂新,吳玲,等.三種藥用甘草種子對鹽漬環境的萌發響應及適宜生態種植區[J].草業學報,2013,22(2):195-202.
[17]羅金明,鄧偉,張曉平,等.蘇打鹽漬土的微域特征以及水分的遷移規律探討[J].土壤通報,2009,(3):482-486.
[18]宋彥濤.松嫩草地豆科植物群落與羊草群落特征比較研究[D].長春:東北師范大學,2008.
[19]陶巖,周道瑋,江源,等.松嫩平原鹽堿化草甸中5種豆科植物的分布格局研究[J].北京師范大學學報(自然科學版),2008,44(4):539-544.
[20]中國科學院土壤研究所.土壤理化分析方法[M].上海:上海科技出版社,1978.
[21]Leimu R,Mutikainen P I A,Koricheva J,et al.How general are positive relationships between plant population size,fitness and genetic variation[J].Journal of Ecology,2006,94(5):942-952.
[22]Hector A,Bazeley-White E,Loreau M,et al.Overyielding in grassland communities:testing the sampling effect hypothesis with replicated biodiversity experiments[J].Ecology Letters,2002,5(4):502-511.
[23]江小雷,張衛國.一年生植物群落內取樣效應和互補效應的分離[J].生態學報,2006,26(6):1896-1902.
[24]Adams T P,Purves D W,Pacala S W.Understanding height-structured competition in forests:is there an R*for light[J].Proceedings of the Royal Society B:Biological Sciences,2007,274:3039-3048.
[25]武高林,陳敏,杜國禎.三種高寒植物幼苗生物量分配及性狀特征對光照和養分的響應[J].生態學報,2010,30(1):60-66.
[26]Kardol P,Martijn Bezemer T,Van Der Putten W H.Temporal variation in plant-soil feedback controls succession[J].Ecology Letters,2006,9(9):1080-1088.
[27]藺吉祥,李曉宇,張兆軍,等.溫度與鹽堿脅迫交互作用對羊草種子萌發與幼苗生長的影響[J].草地學報,2011,19(6):1005-1009.
[28]管博,周道瑋,田雨,等.鹽堿及變溫條件對花苜蓿種子發芽的影響[J].中國草地學報,2009,32(1):58-63.
[29]李曉宇,藺吉祥,李秀軍,等.羊草苗期對鹽堿脅迫的生長適應及Na+、K+代謝響應[J].草業學報,2013,22(1):201-209.
[30]楊光.鹽堿脅迫對豆科牧草種子萌發及其生理的影響[D].長春:東北師范大學,2009.
[31]周道瑋,李強,宋彥濤,等.松嫩平原羊草草地鹽堿化過程[J].應用生態學報,2011,22(6):1423-1430.
[32]李強,劉延春,周道瑋,等.松嫩退化草地三種優勢植物群落對封育的響應[J].東北師范大學學報(自然科學版),2009,41(2):139-144.
[33]宋彥濤.松嫩草地植物功能生態學研究[D].長春:東北師范大學,2012.
[34]Nathan R,Muller-Landau H C.Spatial patterns of seed dispersal,their determinants and consequences for recruitment[J].Trends in Ecology and Evolution,2000,15(7):278-285.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
