油茶油體觀察及其與含油量的相關性
2014-03-13 08:54:41羅麗萍杜尚廣江香梅付宇新郭夏麗
中國糧油學報 2014年11期
羅麗萍 肖 萍 杜尚廣 江香梅 付宇新 郭夏麗
(南昌大學生命科學與食品工程學院1,南昌 330031)
(江西省林業科學院2,南昌 330031)
脂類是種子貯藏能量最有效的形式[1]。在種子發育過程中,脂類主要以三酰甘油(triacylglycerols,TAGs)的形式在內質網上合成后被運輸到胞內的貯油細胞器-油體(oil bodies)或稱為脂肪體(lipid bodies)中,為種子萌發和幼苗生長提供能量[2-3]。油體是植物細胞中最小的細胞器,其直徑為0.5~2.5μm,且因植物種類、組織來源等不同[4-5]。油體十分穩定,在干燥種子細胞內或經離心分離,油體間均不發生融合或聚合[6]。近年來,對油體形態結構、蛋白質成分、結構功能和分子水平以及利用油體構建生物反應器生產外源蛋白和發展生物柴油等方面有較多研究報道[4,7]。
油茶(Camellia oleifera)是我國特有的木本油料樹種,與油棕、油橄欖和椰子并稱為世界四大木本食用油料植物[8]。成熟的油茶種子經榨取所得油稱為茶油,茶油色清味香、營養豐富,被譽為“東方橄欖油”,是高級烹調食用油[9],也是醫藥和化妝品等的主要原料。含油量是油料植物的主要農藝性狀和育種指標,提高含油量一直是油茶育種中最重要的問題。
種子中油體與含油量關系一直受到人們的關注。Ting等[10]研究發現,高油玉米品種具有較大的油體和較高的TAG/油質蛋白比。Peng等[11]對芝麻油體進行了人工合成,合成的油體大小與TAG/油體蛋白成正比。Siloto等[12]通過在擬南芥中調控油質蛋白基因的表達水平,證明了油體大小的變化會影響種子內脂肪酸的累積,從而影響到種子的含油量。Hu等[13]研究歐洲油菜油體結構與含油量之間的關系,發現顯著的大油體與低含油量之間存在高度相關。董勁松等[14]觀察成熟甘藍型油菜種子子葉細胞內油體發現,油菜種子細胞中油體的總面積與含油量之間存在正相關。少數研究者對油茶油體進行了報道,包括對油茶種子的油體蛋白進行了分離和鑒定,對其基因的分離和生物信息學進行了分析,還研究了油體蛋白的原核融合表達載體的構建[15],而有關油茶油體的形態、大小以及與種子含油量之間的關系均鮮見報道。
為觀察油茶油體的形態大小,研究油茶油體與種子含油量之間的關系,本試驗將冰凍切片、熒光染料尼羅紅染色和激光掃描共聚焦顯微鏡(laser scanning confocalmicroscope,LSCM)觀察技術相結合,建立了油茶油體快捷觀察與鑒定技術體系;并對不同含油量的油茶品系的油體進行觀察和統計分析,探討油茶種子含油量與油體的關系。研究結果可為油茶油體改造,高油油茶品系選育等提供理論依據。
1 材料與方法
1.1 試驗材料
選擇成熟干燥的油茶種子,7個油茶品系名及含油量分別為:遂巾子52.82%;珍珠子51.32%;二水桃50.20%;石柿紅皮50.13%;觀音桃48.73%;贛萍茶47.88%;茅崗大果46.50%,均由江西省林業科學院提供。
1.2 試驗方法
1.2.1 冰凍切片最適厚度以及尼羅紅染液最適濃度的篩選
選取外觀正常、飽滿的油茶種子,去除種皮后,切取子葉中間1.0 cm×0.5 cm×0.5 cm大小,放入福爾馬林-醋酸-酒精混合液(FAA液)于4℃冰箱中固定15 h后,在-20℃下,用冰凍切片機(Leica CA 1950)切片,切片放入尼羅紅染液[16]中避光孵育15 min,期間用槍頭吹打幾次,促使染料與細胞內油脂充分結合。切片厚度分別設定為15、20、25、30、35、40μm。尼羅紅染液配制方法為:稱取5 mg尼羅紅(J&K百靈威科技)溶于 5 mL丙酮中,得到1 mg/mL的母液,分裝后在4℃避光保存[17]。用雙蒸水將尼羅紅母液分別稀釋到濃度為1、2、5、10、20、40μg/mL現配現用。
1.2.2 制片、觀察
染色后用0.1 mol/L PBS緩沖液漂洗3次,洗去未結合的染料。制片,用浸鏡油封片。在 LSCM(ZEISSLSM 710)下使用20倍和40倍物鏡掃描觀察并拍照,掃描參數為HeNe激光,激發波長為483 nm,吸收波長為566~670 nm。
1.2.3 圖片處理與數據統計
測定油茶品系遂巾子、珍珠子、二水桃、石柿紅皮、觀音桃、贛萍茶、茅崗大果共7個品系單個子葉貯藏細胞內油體截面積。利用ZEN 2008 Light Eddition軟件的測量功能,每個品系隨機選取30個油體測定其面積、周長,推算出油體直徑;再每個品系隨機選取30個細胞進行單個細胞內油體截面積計算與統計,設定名詞:含油體率=單個細胞內油體截面積÷整個細胞面積大小×100%,通過量化,方便比較各個品系間的油體的含量的差異。
2 結果與分析
2.1 冰凍切片與染色的探討
經過對不同厚度切片進行不同濃度尼羅紅染液的染色比較觀察,最終確定適合油茶種子子葉細胞內油體觀察的切片厚度為25μm,尼羅紅染液的濃度為10μg/mL,染色時間為15 min。由于細胞內油體的完整觀察需要合適的細胞厚度,太厚油體過分重疊,太薄則油體可能損失。而用熒光染料尼羅紅對切片進行染色目的是使其與細胞內油脂物質相結合并發出熒光檢測信號,染液濃度太低染色時間太短,則染液無法與細胞內脂類物質結合完全。因此在保證細胞內脂類物質完全染色的情況下縮短時間,以達到快捷準確的目的。在制片過程中,用浸鏡油進行封片,可達到減少尼羅紅熒光染料淬滅以及延遲子葉貯藏細胞失水收縮的效果[14]。
2.2 油茶種子中油體觀察
從7個油茶品系成熟種子子葉貯藏細胞內油體的LSCM觀察圖片(圖1)中可以看出油茶種子子葉貯藏細胞內油體為球形,未見橢圓或不規則形狀,與董勁松等[14]報道的甘藍型油菜油體的形態有所不同。

圖1 7個油茶品系成熟種子子葉貯藏細胞內油體的LSCM觀察(Bar=10μm)
經測定,每個品系的平均直徑見表1。7個品系油茶油體平均直徑為2.52μm,標準差為1.99μm,圖2中顯示了油茶種子油體大小的分布情況,從圖2可以看出多數油體直徑分布在2.00~3.00μm之間。Tzen等[5]報道,油菜、玉米、花生、芝麻的油體直徑分別為0.65、1.45、1.95、2.00μm,由此可看出不同植物種子油體的大小不同。
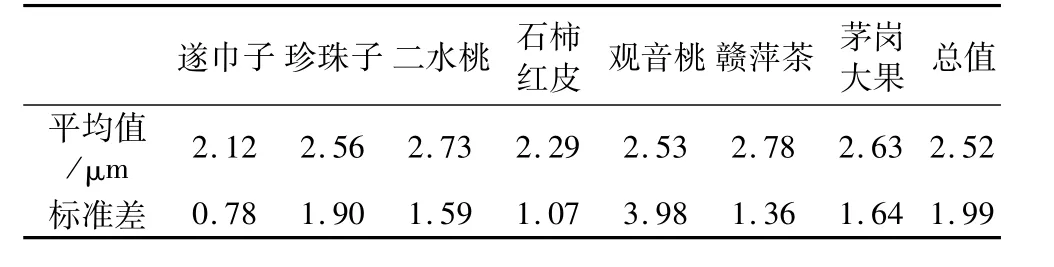
表1 7個油茶品系成熟種子子葉貯藏細胞內油體直徑
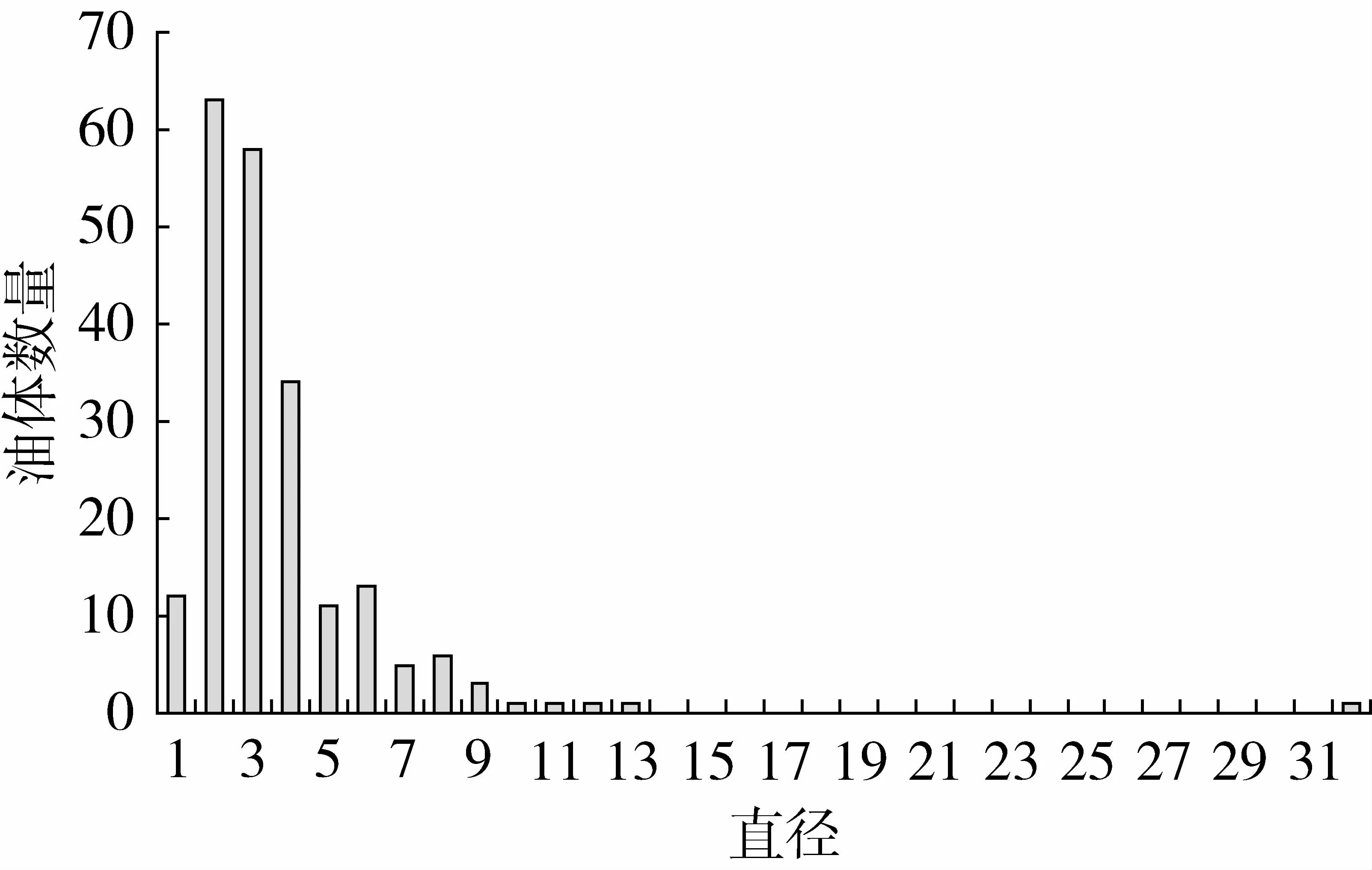
圖2 油茶種子油體大小分布
迄今為止,國內外植物油體的形態觀察已報道的植物有擬南芥[12]、甘藍型油菜[13-14]、玉米[10]、大豆[18]等。通過對所報道的植物種類的油體大小進行比較[5],發現不同物種的種子中油體的大小不同,直徑大小由小到大依次是:油菜、芥菜、棉花、亞麻、玉米、花生、芝麻、油茶。除油茶為木本植物外,前面7種均為草本植物,而油茶油體在其中為最大的,由此推測油體的大小可能與植物的類型也存在一定關系。
2.3 不同含油量的油茶種子子葉貯藏細胞內油體的分布觀察
從7個油茶品系成熟種子子葉貯藏細胞內油體的LSCM觀察圖片(圖1)中可以看出含油量越高的油茶品系細胞染色越深,細胞內油體排列更緊密,油體數量越多,并集中堆疊分布于細胞中央,不與細胞壁接觸,邊緣清晰;而含油量越低的品系細胞染色越淺,細胞內油體或分布于細胞壁邊緣,或稀疏分散于細胞內,油體數量越少且不連續。此結果與相關文獻所報道的甘藍型油菜油體的描述結果類似[14,19]。
2.4 不同含油量油茶種子子葉貯藏細胞內含油體率統計分析
為準確反映不同含油量的油茶品系種子子葉細胞內油體含量的差異,從獲取的圖片中選取有代表性的細胞進行細胞面積與細胞內油體截面積之和的測量,并計算出含油體率(表2)。從表2中可以看出,7個油茶種子子葉細胞內油體量隨著含油量的增加而增加,對油茶種子含油量和含油體率繪制標準曲線(圖3)。可知油茶種子含油量越高,種子子葉細胞內含油體率越高,種子含油量與含油體率成正相關,與 Ting等[10]、Peng等[11]、董勁松等[14]所報道的結果一致。

表2 7個油茶品系含油量與含油體率的比較/%
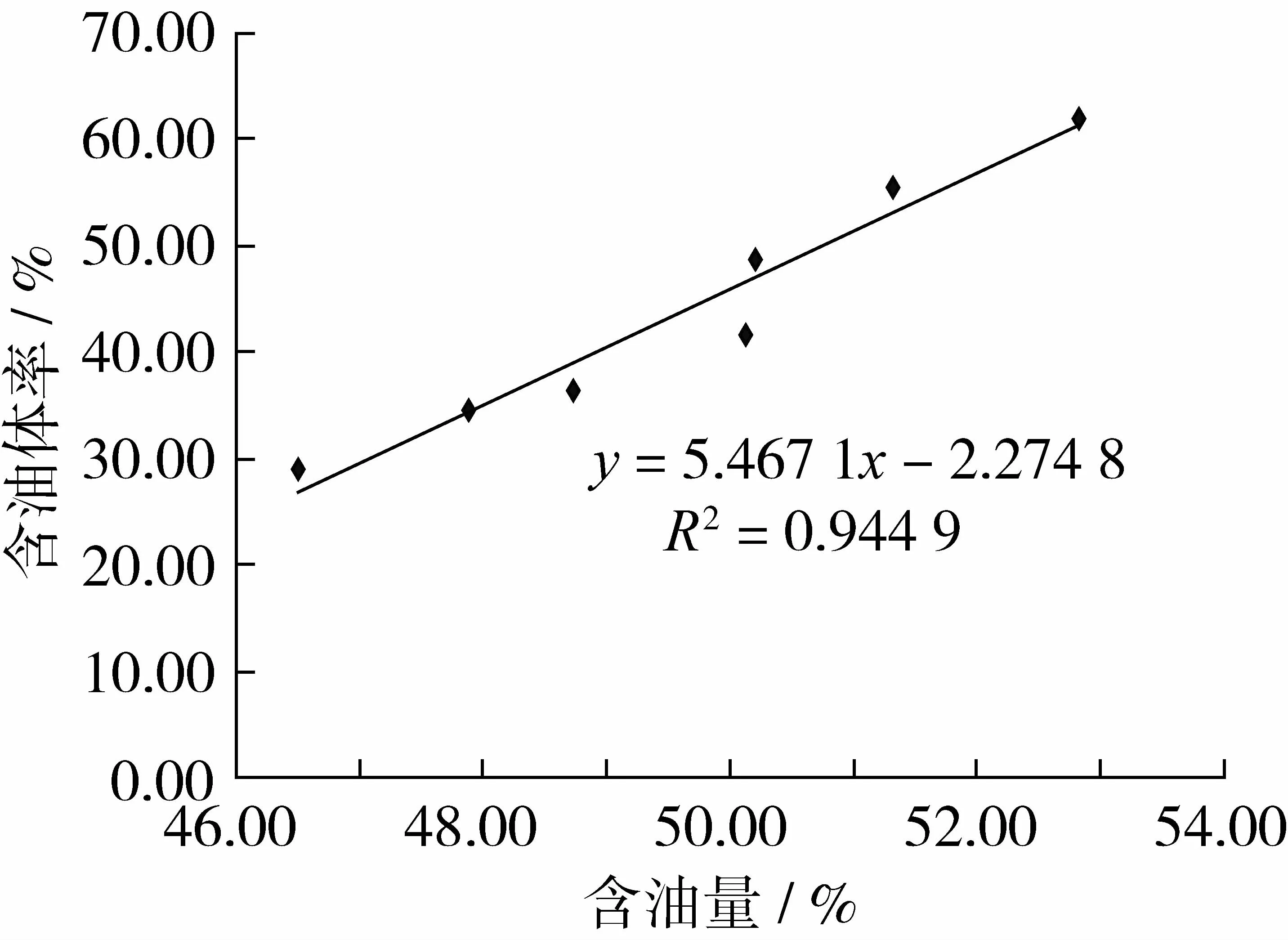
圖3 油茶種子含油量和含油體率標準曲線
3 結論
通過測量不同含油量的油茶品系種子子葉細胞內的油體直徑,未發現有明顯差異,但是油體在細胞內的分布與截面面積差異較明顯。油茶種子含油量越高,油體排列越緊密,截面面積越大。
采用冰凍切片機切片、熒光染料尼羅紅染色和LSCM顯微觀察相結合技術,建立了油茶油體準確快捷觀察與鑒定的研究體系,可為油茶油體的研究提供技術支持。利用含油量與含油體率的相關性以及不同含油量油茶種子子葉細胞內油體形態、分布的差異,可快速準確地判斷油茶種子的含油量高低,為油茶高油品系的選育提供理論依據。
[1]仇鍵,譚曉風.植物種子油體及相關蛋白研究綜述[J].中南林學院學報,2005(25):96-100
[2]Huang A H C.Oil bodies and oleosins in seeds[J].Annual Review of Plant Physiology and Plant Molecular Biology,1992,43:177-200
[3]Huang A H.Oleosins and Oil Bodies in Seeds and Other Organs[J].Plant Physiology,1996,110:1055-1061
[4]程紅焱,宋松泉.種子的貯油細胞器—油體及其蛋白[J].植物學通報,2006(23):418-430
[5]Tzen J T C,Cao Y Z,Laurent P,et al.Lipids,proteins,and structure of seed oil bodies from diverse species[J].Plant Physiology,1993,101:267-276
[6]Frandsen G I,Mundy J,Tzen JTC.Oil bodies and their associated proteins,oleosin and caleosin[J].Physiologia Plantarum,2001,112:301-307
[7]趙傳志,盧金東,蘇磊,等.以油體作為生物反應器的研究進展[J].生物技術通報,2008(2):73-76
[8]莊瑞林.中國油茶(第2版)[M].北京:中國林業出版社,2008:54-127
[9]馬力.茶油與橄欖油營養價值的比較[J].糧食與食品工業,2007,14:42-44
[10]Ting JT L,Lee K,Ratnayake C,et al.Oleosin genes in maize kernels having diverse oil contents are constitutively expressed independent of oil contents[J].Planta,1996,199:158-165
[11]Peng CC,Lin IP,Lin CK,et al.Size and stability of reconstituted sesame oil bodies[J].Biotechnology Progress,2003,19:1623-1626
[12]Siloto R M P,Findlay K,Lopez-villalobos A,et al.The accumulation of oleosins determines the size of seed oilbodies in Arabidopsis[J].The Plant Cell,2006,18:1961-1974
[13]Hu Z Y,Wang X F,Zhan GM,etal.Unusually large oilbodies are highly correlated with lower oil content in Brassica napus[J].Plant Cell Reports,2009,28:541-549
[14]董勁松,石東喬,高建芹,等.甘藍型油菜油體數量及面積之和與含油量的相關性[J].植物學報,2009(44):79-85
[15]仇鍵.油茶種子油體蛋白基因的分離克隆及其原核表達[D].長沙:中南林業科技大學,2006
[16]Greenspan P,Mayer E P,Fowler SD.Nile red:a selective fluorescent stain for intracellular lipid droplets[J].The Journal of Cell Biology,1985,100:965-973
[17]董勁松.甘藍型油菜(Brassica napus L.)油體發育機制及其與含油量相關性研究[D].南京:南京農業大學,2008
[18]SchmidtM A,Hermanb EM.Suppression of soybean oleosin producesmicro-oil bodies that aggregate into oil body/ER complexes[J].Molecular Plant,2008(1):910-924
[19]代柳亭.不同含油量甘藍型油菜種子油脂分布、生理生化特性以及化學調控的研究[D].重慶:西南大學,2008.
