食欲素與血糖和能量代謝的調節
2014-03-27 00:50:18綜述陸穎理審校
醫學綜述 2014年3期
翁 盼(綜述),陸穎理(審校)
(1.上海交通大學醫學院,上海 200011; 2.上海交通大學醫學院附屬第九人民醫院內分泌代謝科,上海 200011)
食欲素是在1998年首先被發現的,包括食欲素A(含33個氨基酸殘基)和食欲素B(含28個氨基酸殘基),是由同一前體蛋白裂解產生的一對興奮性神經肽激素,食欲素由下丘腦中特定的神經元產生,主要位于外側下丘腦區、穹窿周圍區,以及下丘腦后部[1]。在過去的十年里,許多實驗已經證實,食欲素在調控一系列生理活動中扮演重要角色,包括調節進食、睡眠、覓食和能量血糖動態平衡等。該文就食欲素與血糖、能量代謝調節和肥胖的研究進展予以綜述。
1 食欲素與血糖調節和胰島素抵抗
1.1血糖的來源與調節因素 血糖的主要來源有食物和肝臟的內源性葡萄糖生成,前者提升血糖的程度主要由食物成分決定,后者主要受胰島素和胰高血糖素的調節,但也有一部分直接受自主神經支配,經下丘腦控制。胰島素和副交感神經增加肝臟葡萄糖攝取和糖原合成,而胰高血糖素和交感神經促進糖原分解、糖異生和葡萄糖釋放。血糖水平也受肌肉和脂肪組織攝取葡萄糖的影響。
1.2食欲素及受體的分布與作用特征 食欲素是下丘腦血糖調節的重要因子,食欲素纖維分布于整個腦干,包括迷走神經背核和孤束核。食欲素神經元受谷氨酸、促腎上腺皮質素釋放因子,神經降壓素,血管加壓素、催產素和促甲狀腺素釋放激素的興奮,而γ-氨基丁酸(GABA)、5-羥色胺、去甲腎上腺素、多巴胺和神經肽Y對食欲素神經元有抑制作用[1]。外周的代謝信號也影響食欲素神經元的活性,葡萄糖和瘦素抑制食欲素神經元的活性,而生長激素促分泌素受體內源性配體則促進其活性。
食欲素主要通過兩個G蛋白偶聯受體,即食欲素受體1(orexin receptor 1,OXR1)和食欲素受體2(orexin receptor 2,OXR2)發揮作用,OXR1與食欲素A結合的親和力比食欲素B高100倍,而OXR2與兩者結合的親和力基本相同。下丘腦腹內側核是OXR1 mRNA最豐富的區域,而OXR2 mRNA高表達于下丘腦結節乳頭核、室旁核、弓狀核以及外側下丘腦區。
1.3食欲素與血糖穩態的中樞性調節機制
1.3.1食欲素與血糖的雙重調節 下丘腦食欲素系統參與調節血糖穩態:在這個系統中,食欲素神經元精確感知外周營養狀態,調節自主神經平衡,控制外周組織產生和利用葡萄糖。下丘腦食欲素神經元能刺激交感神經節前纖維和副交感神經元,通過改變自主神經平衡來對血糖穩態進行雙重調節(圖1)[2]。

圖1 食欲素與血糖穩態雙重調節
研究發現,食欲素A可直接激活腹內側下丘腦神經元,促進交感神經系統激活,然后經由骨骼肌細胞內的β2腎上腺素受體信號介導,增加骨骼肌攝取葡萄糖,促進胰島素誘導的葡萄糖攝取和肌糖原合成[3-4]。
Ramadori等[5]通過應用一種在下丘腦腹內側核神經元SF1缺乏食欲素代謝傳感器煙酰胺腺嘌呤二核苷酸(NAD+)依賴的脫乙酰酶1(sirtuin type 1,SIRT1)的小鼠(Sf1-Cre;Sirt1loxP/loxP mice)來研究經腹內側核調控血糖平衡的機制。當給予高熱量飲食后,與正常小鼠相比,Sf1-Cre;Sirt1loxP/loxP小鼠體質量增加更顯著,能量消耗減少,瘦素抵抗增加。另外,高熱量飲食喂養的基因突變小鼠(Sf1-Cre;Sirt1loxP/loxP mice)在出現嚴重肥胖之前就已經呈現出高胰島素血癥、糖耐量受損和胰島素抵抗。對于高熱量飲食喂養的對照組小鼠,持續側腦室注射食欲素 A(0.5 nmol/d)能改善高血糖和高胰島素血癥,但在上述基因突變的小鼠中未觀察到這種改善現象。這些實驗結果提示,下丘腦腹內側核是食欲素調控肌肉葡萄糖利用的主要部位。
另一方面,食欲素對肝臟葡萄糖生成的調節可能一部分是通過調控人體生物鐘完成的。生物鐘位于下丘腦視交叉上核,而下丘腦室旁核控制著交感和副交感神經的活動,后者是生物鐘信號輸出的一個重要靶點,視交叉上核直接控制著血漿葡萄糖濃度的日常節律改變,且不受進食的影響。空腹大鼠室旁核內應用GABA受體拮抗劑進行干預后發現,僅在白天可引起高血糖癥,提示從視交叉上核傳遞到室旁核的GABAergic神經元信號的晝夜改變會引起肝臟葡萄糖生成的相應改變[6]。視交叉上核的生物鐘可受到由食欲素神經元產生的促覺醒信號的重置,而食欲素神經元的激活很大程度上又受到生理節奏的控制,提示兩者之間存在互惠關系[7]。
1.3.2食欲素、胰島素、瘦素和血糖調節的關系 在胰島素對肝臟葡萄糖生成的抑制效應中,下丘腦信號作用是必須的。下丘腦-肝臟通路的任何改變都可能會造成糖尿病高血糖癥。外周胰島素抵抗相關的高血糖癥抑制下丘腦食欲素的表達,而食欲素有助于維持下丘腦胰島素敏感性,食欲素表達受到抑制之后便產生了下丘腦和外周胰島素抵抗的惡性循環(圖2)[8]。正常情況下,胰島素在下丘腦的作用可促進胰島素的降糖效應,特別是通過抑制肝臟葡萄糖生成。這個過程由下丘腦ATP敏感性鉀通道(KATP)通道介導的。另外,下丘腦蛋白激酶C(可能是蛋白激酶Cδ亞型)的活化對于激活下丘腦KATP,從而降低肝臟葡萄糖生成是非常必要的。瘦素也可通過信號轉導與轉錄激活因子3通路調節肝臟葡萄糖生成。另一方面,年齡或食源性胰島素抵抗相關的高血糖癥的患者,其下丘腦食欲素的表達減少。反過來,食欲素的缺乏可導致下丘腦胰島素受體、瘦素受體以及食欲素受體信號中斷(具體機制不明)。結果就是調節能量(葡萄糖)代謝的下丘腦/肝臟通路受損,從而進一步加劇外周組織的胰島素抵抗。
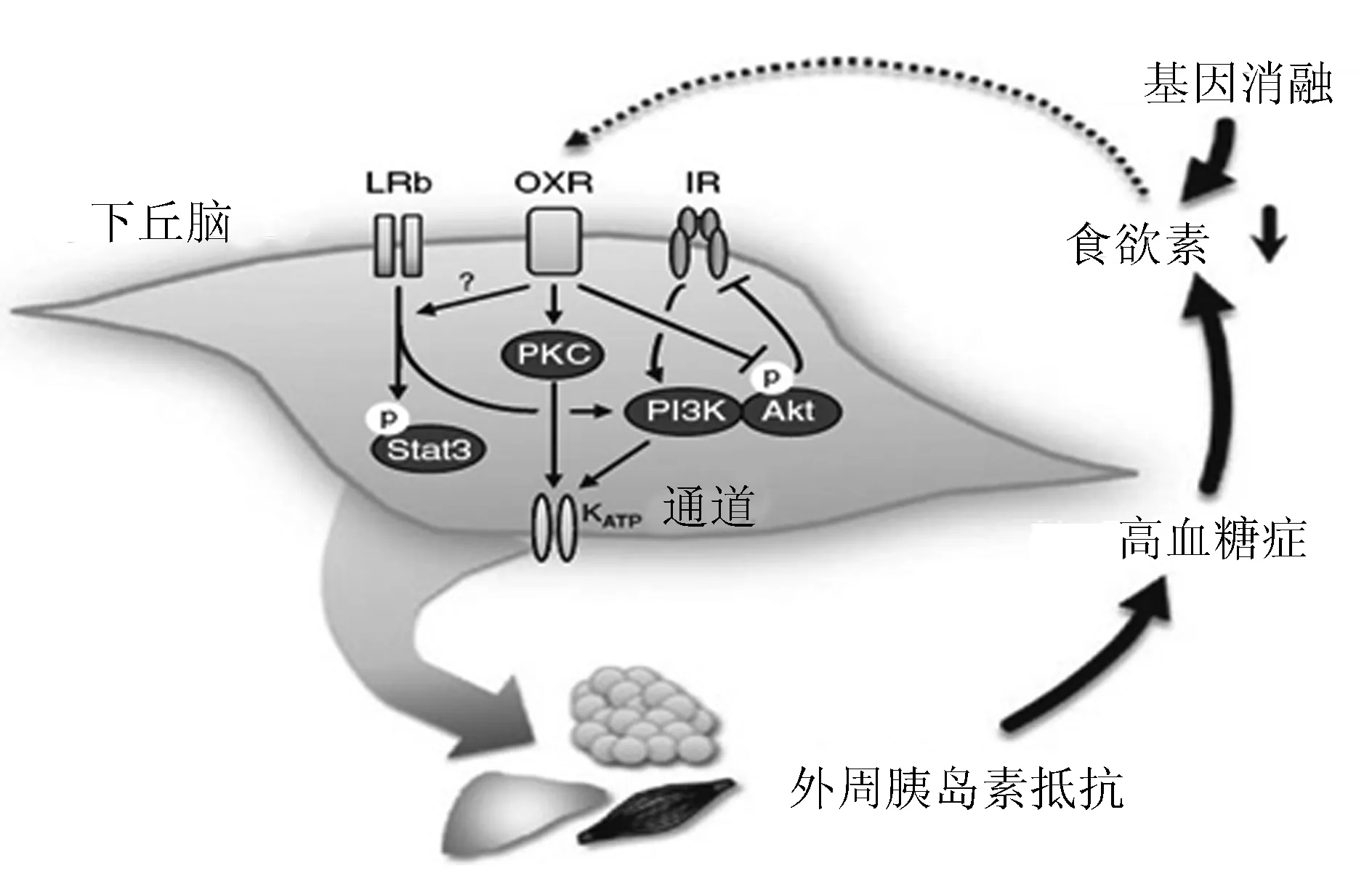
LRb:瘦素受體;OXR:食欲素受體;IR:胰島素抵抗;PI3K:磷脂酰肌醇3-激酶;Akt:蛋白激酶B;Stat3:信號轉導與轉錄激活因子3;PKC:下丘腦蛋白激酶C
圖2食欲素、瘦素、胰島素循環軸與血糖調節的關系
1.3.3食欲素影響胰島素、瘦素敏感性的機制 雖然食欲素影響胰島素、瘦素敏感性的確切機制仍不明確,但是很可能在食欲素缺乏的情況下(例如前食欲素原基因消融或者高血糖癥),下丘腦蛋白激酶B磷酸化水平會隨著年齡增長而增高,這種長期的磷酸化作用破壞了胰島素在下丘腦的作用,從而影響外周胰島素敏感性。
在正常飲食條件下,食欲素基因剔除的小鼠出現隨年齡增長呈進行性發展的糖耐量受損和胰島素抵抗,觀察對象包括非肥胖的雄性小鼠和輕度肥胖的雌性小鼠。9月齡的食欲素基因剔除的小鼠,在正常飲食條件下,通過蛋白激酶B通路的胰島素信號在骨骼肌和肝臟中斷。2~6月齡的食欲素基因剔除的小鼠,下丘腦蛋白激酶B(Ser473)磷酸化水平出現異常增加,且胰島素的刺激不能提升蛋白激酶B磷酸化水平,而在9月齡的食欲素基因剔除小鼠中也觀察到胰島素誘導的胰島素受體酪氨酸磷酸化減少,提示食欲素是維持下丘腦和外周胰島素敏感性所必須的,尤其隨著年齡的增長更是如此。運用示蹤稀釋法結合胰島素鉗夾技術的研究證實,下丘腦蛋白激酶C的激活對于降低肝臟葡萄糖生成是非常重要且必須的,主要通過下丘腦磺脲類藥物受體1/Kir6.2 KATP的激活/磷酸化,而食欲素A可強烈激活CHO細胞上的蛋白激酶Cδ,穩定表達人OXR1[9]。因此,食欲素可能是通過促進KATP的激活來增強胰島素在下丘腦的作用,從而抑制肝臟葡萄糖生成。
近幾年的研究證實,在嚙齒動物中食欲素可強烈抑制食源性肥胖以及后續出現的胰島素抵抗。食欲素過表達的轉基因小鼠可通過增加能量支出、減少能量消耗而預防高脂飲食誘導的肥胖、胰島素不敏感、高胰島素血癥和高瘦素血癥。將食欲素過表達的OXR1和OXR2基因敲除小鼠進行分子遺傳分析發現OXR2主要介導了這種抗肥胖效應和改善胰島素敏感性的效應。持續應用一種OXR2選擇性興奮劑食欲素B([Ala11,d-Leu15]-Orexin B)進行中樞干預可預防食源性肥胖。另外,食欲過表達的轉基因小鼠表現為對外源性輸注的瘦素的敏感性增加,食欲素過表達或OXR2興奮劑均不影響瘦素缺乏的肥胖ob/ob小鼠的體質量增加,說明食欲素經OXR2的作用與瘦素敏感性狀態相關[10]。需要注意的是,單獨的OXR1缺乏可預防高脂飲食誘導的高血糖癥和高胰島素血癥,但無法防止肥胖,而食欲素過表達可預防OXR1和OXR2基因敲除的小鼠在高脂飲食的情況下發生肥胖。這些結果提示,內源性食欲素-OXR1信號可能介導了高脂飲食對糖代謝和肥胖的有害作用。事實上,已經有報道一種OXR1拮抗劑SB334867A可能成為抗肥胖和糖尿病的因子,因為它能減少肥胖ob/ob小鼠的攝食量,降低體質量、空腹血糖和血漿胰島素水平[11]。而食欲素-OXR2信號的增加可防止食源性肥胖,提高瘦素敏感性。
因此,食欲素可通過增加下丘腦胰島素和(或)瘦素信號來預防年齡相關及飲食誘導的胰島素抵抗,OXR可作為未來治療2型糖尿病患者高血糖癥的重要靶點。
2 食欲素與能量代謝調節
關于食欲素對能量代謝的調節機制,目前仍不明確。下丘腦食欲素誘導的能量代謝的增加并不僅僅是由于覺醒和身體活動的增加。事實上,食欲素A會引起氧耗增加和體溫上升,即使麻醉的大鼠也是如此,小鼠腦室內注射食欲素A能在不改變攝食和體育活動的情況下增加代謝率。另外,食欲素神經元缺失的小鼠表現為能量消耗減少,并且這一改變與睡眠/覺醒、運動和進食無關。
已知線粒體解偶聯蛋白1(uncoupling protein 1,UCP1)與非寒戰產熱有關,并且主要受交感神經系統支配,其原因是β去甲腎上腺素能遞質直接支配棕脂肪組織進行調控[12]。這種產熱過程在機體能量代謝平衡中非常重要。食欲素A可激活交感神經系統,食欲素基因剔除的小鼠表現為較低的交感縮血管纖維緊張性[13]。然而,長期緩慢室旁核內注射食欲素A并不改變大鼠棕脂肪組織中UCP1的水平[14]。大鼠第三腦室內注射食欲素A后也觀察到了類似的結果。此外,低脂飲食的情況下,食欲素過表達的轉基因小鼠的棕脂肪組織中UCP1 mRNA的水平與對照組小鼠比較差異無統計學意義[15]。因此,食欲素可能不是通過棕脂肪組織中的UCP1來調控能量代謝的。
與UCP1不同,UCP2在全身廣泛表達,包括中樞神經系統,增加UCP2引發的線粒體解耦聯可使活性氧類生成減少,產生神經保護效應。UCP2也能增加海馬細胞中線粒體的數量。外側下丘腦區表達的食欲素A和食欲素B為熱能的產生提供了一個解剖學上受限的位點[16]。食欲素神經元UCP2過表達的轉基因小鼠(orexin/UCP2-Tg mice)下丘腦溫度升高,伴隨身體核心溫度降低。但是,也有一些報道提示與之矛盾的結果,因此食欲素是如何通過UCP2調節能量代謝的機制仍有待進一步研究。
3 食欲素在外周組織的作用
食欲素大部分是在下丘腦發揮其功能,在神經系統以外的作用目前仍存在爭議,食欲素及其受體存在于廣泛的組織中,包括腸道、胰腺、腎臟、腎上腺、脂肪組織和生殖系統。
研究證實,隨著鏈脲佐菌素誘導的糖尿病的發生,大鼠胰島細胞中OXR1的表達增加,糖尿病病程越長,胰島細胞中OXR1的表達越強烈,并且與胰高血糖素共定位[17]。另外,免疫組織化學發現在胰島有被剪切的胱天蛋白酶3片段與OXR1共定位。食欲素基因剔除的小鼠中OXR1和胱天蛋白酶3的表達顯著降低,說明在胰腺組織中食欲素可能是通過OXR1介導促進鏈脲佐菌素誘導的糖尿病的發展和β細胞凋亡[18-19]。但是,關于食欲素在外周組織中的作用目前仍然知之甚少,有待進一步研究。
4 小結與展望
食欲素參與調控血糖平衡,調節胰島素/瘦素敏感性,食欲素可有效預防年齡相關或高脂飲食誘導的外周胰島素抵抗的進行性發展。食欲素-OXR2信號可通過增加瘦素敏感性而預防食源性肥胖和胰島素抵抗。隨著年齡增長,海馬、下丘腦、腦橋等組織中OXR2 mRNA水平著顯下降,另外,前食欲素原基因表達在肥胖的ob/ob和db/db小鼠出現下調,并且這種下調現象是由高血糖導致的。這些現象都提示,胰島素敏感性下降所導致的高血糖癥造成下丘腦食欲素的表達降低,從而加劇了外周胰島素抵抗。因此,控制下丘腦胰島素/瘦素信號的食欲素受體可作為2型糖尿病高血糖癥治療的新靶點。
[1] Burdakov D,González JA.Physiological functions of glucose-inhibited neurones[J].Acta Physiol(Oxf),2009,195(1):71-78.
[2] Marino JS,Xu Y,Hill JW.Central insulin and leptin-mediated autonomic control of glucose homeostasis[J].Trends Endocrinol Metab,2011,22(7):275-285.
[3] Shen J,Tanida M,Yao JF,etal.Biphasic effects of orexin-A on autonomic nerve activity and lipolysis[J].Neurosci Lett,2008,444(2):166-171.
[4] Shiuchi T,Haque MS,Okamoto S,etal.Hypothalamic orexin stimulates feeding-associated glucose utilization in skeletal muscle via sympathetic nervous system[J].Cell Metab,2009,10(6):466-480.
[5] Ramadori G,Fujikawa T,Anderson J,etal.SIRT1 deacetylase in SF1 neuronsprotects against metabolic imbalance[J].Cell Metab,2011,14(3):301-312.
[6] Kalsbeek A,Yi CX,La Fleur SE,etal.The hypothalamic clock and its control of glucose homeostasis[J].Trends Endocrinol Metab,2010,21(7):402-410.
[7] Marston OJ,Williams RH,Canal MM,etal.Circadian and dark-pulse activation of orexin/hypocretin neurons[J].Mol Brain,2008,1:19.
[8] Ross R,Wang PY,Chari M,etal.Hypothalamic protein kinase C regulates glucose production[J].Diabetes,2008,57(8):2061-2065.
[9] Johansson L,Ekholm ME,Kukkonen JP.Multiple phospholipase activation by OX1 orexin/hypocretin receptors[J].Cell Mol Life Sci,2008,65(12):1948-1956.
[10] Funato H,Tsai AL,Willie JT,etal.Enhanced orexin receptor-2 signaling prevents diet-induced obesity and improves leptin sensitivity[J].Cell Metab,2009,9(1):64-76.
[11] Haynes AC,Chapman H,Taylor C,etal.Anorectic,thermogenic and anti-obesity activity of a selective orexin-1 receptor antagonist in ob/ob mice[J].Regul Pept,2002,104(1/3):153-159.
[12] Golozoubova V,Hohtola E,Matthias A,etal.Only UCP1 can mediate adaptive nonshivering thermogenesis in the cold[J].FASEB J,2001,15(11):2048-2050.
[13] Kayaba Y,Nakamura A,Kasuya Y,etal.Attenuated defense response and low basal blood pressure in orexin knockout mice[J].Am J Physiol Regul Integr Comp Physiol,2003,285(3):R581-R593.
[14] Russell SH,Small CJ,Sunter D,etal.Chronic intraparaventricular nuclear administration of orexin A in male rats does not alter thyroid axis or uncoupling protein-1 in brown adipose tissue[J].Regul Pept,2002,104(1/3):61-68.
[15] Conti B,Sanchez-Alavez M,Winsky-Sommerer R,etal.Transgenic mice with a reduced core body temperature have an increased life span[J].Science,2006,314(5800):825-828.
[16] Adeghate E,Fernandez-Cabezudo M,Hameed R,etal.Orexin-1 receptor co-localizes with pancreatic hormones in islet cells and modulates the outcome of streptozotocin-induced diabetes mellitus[J].PLoS One,2010,5(1):e8587.
[17] Sakurai T,Mieda M.Connectomics of rexinproducing neurons:interface of systems of emotion,energy homeostasis and arousal[J].Trends Pharmacol Sci,2011,32(8):451-462.
[18] Carter ME,Borg JS,de Lecea L.The brain hypocretins and their receptors:mediators of allostatic arousal[J].Curr Opin Pharmacol,2009,9(1):39-45.
[19] Yi CX,Serlie MJ,Ackermans MT,etal.A major role for perifornical orexin neurons in the control of glucose metabolism in rats[J].Diabetes,2009,58(9):1998-2005.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
保健醫苑(2022年6期)2022-07-08 01:26:34
家庭科學·新健康(2022年3期)2022-05-10 00:32:13
學苑創造·A版(2020年9期)2020-10-13 09:41:02
中國生殖健康(2019年2期)2019-08-23 08:12:10
媽媽寶寶(2017年3期)2017-02-21 01:22:30
飼料與畜牧(規模養豬)(2016年5期)2016-12-01 03:48:40
中國衛生標準管理(2015年1期)2016-01-14 03:41:27
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
