植物乳桿菌AY01 luxS基因表達模式與功能的研究
2014-04-29 11:51:28王良才姜黎明康子騰柳陳堅李曉然羅義勇
生命科學研究 2014年3期
關鍵詞:表達
王良才 姜黎明 康子騰 柳陳堅 李曉然 羅義勇
摘 要:植物乳桿菌(Lactobacillus plantarum)是發酵食品中常見的一種益生菌,群體感應(quorum sensing,Qs)是細菌進行信號交流的一種普遍方式,Lb.plantarum是否通過QS介導著其益生特性尚不清楚。為了探索Lb.plantarum AY01是否通過AI-2/LuxS QS系統介導其對常見食源性病原菌的抑制,首先運用半定量PCR方法,研究了AY01在不同生長階段luxS基因(合成Qs信號分子AI-2的關鍵基因)的表達情況;然后,利用牛津杯抑菌法研究了AY01對食源性病原菌腸出血性大腸桿菌(enterohemorrhagic Escherichia coli,EHEC)0157:H7、單核細胞增生李斯特菌(Listeria monocytogenes)和金黃色葡萄球菌(Staphylococcus aureus)的抑菌效果。研究發現:1)Lb.plantarum AY01 luxS基因在指數早期沒有表達,指數中期出現表達,指數末期表達量明顯增加,并于穩定中期達到最大值;2)AY01培養上清液對EHEC 0157:H7、L.monocytogenes、S.aureus均有抑茵效果,抑菌效果與;luxS基因表達趨勢一致,即隨AY01生長時間的增加抑菌呈現增加變化,到穩定期達到最大抑茵效果。結果表明,Lb.plantarum AY01可能通過AI-2/LuxS QS系統介導其對常見食源性病原菌的抑制。
關鍵詞:植物乳桿菌;luxS基因;表達;抑茵;群體感應
中圖分類號:Q786
文獻標識碼:A
文章編號:1007-7847(2014)03-0199-06
乳桿菌(Lactobacillus)是乳酸菌(lactic acidbacteria,LAB)最大的一個屬,因其形態呈桿狀或球狀以及能發酵碳水化合物(主要為葡萄糖)并產生大量乳酸而得名[1]。作為乳桿菌一個主要的種,植物乳桿菌(Lb.plantarum)具有很多益生功能[2],其中對病原菌的抑制功能得到了最廣泛的研究。2011年Moslehi-Jenabian等[3]將病原菌單核細胞增生李斯特菌(Listeria mono-cytogenes)與嗜酸乳桿菌(Lb.acidophilus)NCFM共培養,發現L.monocy-togenes生長明顯受到影響,而Lb.acidophilusNCFM的生長幾乎沒有變化,說明Lb.acidophilusNCFM對病原菌L.monocytogenes具有很好的抑菌效果。Kim等[4]研究發現,Lb. acidophilus A4除了能抑制L.monocyogenes的生長外,對腸出血性大腸桿菌(enterohemorrhagic Escherichia coli,EHEC)0157:H7同樣具有抑制功能。此外,本實驗室之前的研究顯示,Lb.plantarum,是一種具有廣譜抑菌性的乳酸菌,不僅能抑制同種或近親源關系的細菌,還能抑制大多數病原菌如E.coli、L.mono-cytogenes、金黃色葡萄球菌(Staphylococcus aureus)和鼠傷寒沙門氏菌(Sal-monella typhimurium)等菌株[5,6]。
群體感應(quorum sensing,QS)是細菌間通過化學信號分子進行信息傳遞的一種形式。根據細菌信號分子種類的不同,QS系統可分為4種代表性的類型:1)革蘭氏陽性(G+)細菌中的種內信號通訊使用的語言是寡肽;2)革蘭氏陰性fG)細菌利用酰基高絲氨酸內酯作為信號分子實現種內細胞通訊;3)許多革蘭氏陰性菌還可以使用擴散信號因子系統來調節多種生物學功能,例如毒力和生物膜形成等[7];4)不同種的細菌之間也能進行交流,交流所用的語言是AI-2,這種信號分子存在于大多數細菌中,是許多細菌進行種內和種間交流的通用語言[8.9]。AI-2在細菌體內由LuxS催化合成,LuxS基因廣泛存在于G+和G-細菌中,且具有很高的保守性[10]。代謝學研究發現,LuxS是細菌硫代謝中甲基循環中一個代謝酶,在代謝過程中AI-2作為副產物被合成,所以LuxS肩負著代謝和信號傳遞的雙重功能[11,12]。
AI-2/LuxS QS系統介導乳酸菌發揮益生特性已有一定研究。Moslehi-Jenabian等[13]發現luxS基因可能參與某些Lactobacillus的酸適應過程,使細菌可以通過胃腸道而存活下來,并且在腸道微生態系統中可能在細菌與細菌之間的交流中具有一定作用。Buck等[11]得出AI-2/LuxS QS系統在Lb.acidophilus粘附腸表皮,定植腸道過程中具有重要作用;Lebeer等[14]研究推測AI-2/LuxS QS系統可能與Lactobacillus在動物消化道中的適應性密切相關。AI-2/LuxS QS系統與LAB抑菌的相關性研究很少。據報道,一些G+細菌能夠通過QS系統激活Lb. plantarum特異信號途徑,增加細菌素(一種具有抑菌作用的多肽)產量[15,16]。Man等[17,18]不僅發現4種LAB(稱為誘導LAB)與Lb. plan-tarum, KLDSl.0391共培養能使Lb.plantarumKLDSl.0391的細胞數增多,同樣能顯著提高Lb.plantarum KLDSl.0391產生細菌素的能力;以此為基礎,通過進一步基因敲除和表達研究,他們建立了AI-2/LuxS QS系統介導Lb. plantarum KLDS-1.0391細菌素產生的模型:誘導LAB作為環境刺激被Lb. plantarum, KLDSl.0391識別,導致Lb.plantarum KLDSl.0391細胞數顯著增加,造成LuxS合成的信號分子AI-2的大量積累,當該濃度達到一定閾值時,它能夠開啟下游調節元件的轉錄,調控細菌素編碼基因合成細菌素。
在前期的研究中,我們從云南石林路南鮮羊奶中,分離鑒定了一株益生效果較好的乳桿菌。16S rRNA基因測序鑒定其為Lb. plantarum,命名為AY01,并且利用焦磷酸測序策略測定了其全基因組序列[19]。本研究以Lb. plantarum AY01為實驗材料,首先制作它的生長曲線,然后選取跟細菌QS密切相關的指數早期(4.5h)、指數中期(12h)、指數晚期(18h)和穩定中期(27h)4個點為研究對象,應用半定量PCR方法和牛津杯抑菌法分別測定其在這4個點的luxS基因表達水平和對常見食源性病原菌EHEC 0157:H7、L monocytogenes、S.aureus的抑制情況,探討Lb.plantarum AY01luxS基因表達與抑菌的相關性,為AI-2/LuxS QS系統介導乳桿菌的抑菌關系提供理論依據。
1材料與方法
1.1菌株與培養基
Lb.pLantarum AY01由本實驗室從云南石林路南鮮羊奶中分離獲得。常見食源性病原菌E-HEC 0157:H7、L monocytogenes、S.aureus為本實驗室保存菌株。Lb.plantarum培養用MRS培養基(Oxoid,英國),E.coli用營養肉湯培養基(杭州天河微生物試劑有限公司,中國),L.monocytoge-nes、S.aureus用腦心浸液肉湯培養基(青島高科園海博生物技術有限公司,中國)培養。
1.2生長曲線和產酸試驗
首先從-80℃冰箱中取出Lb.plantarumAY01種子液,然后按4‰(V/V)接種于MRS液體培養基中,30℃活化24h后,同樣按4‰(V/V)接種到新鮮的MRS液體培養基中,30℃靜止培養。隔1.5h取樣(0~36h),利用分光光度計測定600nm波長處的吸光值(OD6oo),同時利用pH計測定各樣品的pH值。以培養時間為橫坐標,OD6oo和pH為縱坐標,制作Lb.plantarum AY01生長曲線和產酸趨勢圖。
1.3生長階段luxS基因表達分析
1.3.1cDNA第一條鏈的合成
依據Lb.plantarum AY01生長曲線,用RNAprep pure培養細胞/細菌總RNA提取試劑盒(TIANGEN,中國)提取其指數早期(a點,培養4.5h)、指數中期(b點,培養12h)、指數晚期(c點,培養18h)和穩定中期(d點,培養27h)4點的菌體總RNA,利用瓊脂糖凝膠電泳和吸光值測定法分別分析總RNA的完整性、純度和濃度。最后,以總RNA為模板,利用mRNA selective PCR kit (TAKARA,日本)進行逆轉錄反應,生成第一條鏈cDNA。
1.3.2引物設計
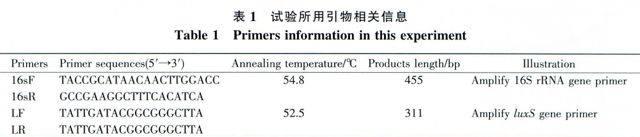
選取看家基因16S rRNA基因作為相對半定量的內參照基因。根據Lb.plantarum AY01全基因組序列[19],利用引物設計軟件Primer Premier5.0(PREMIER Biosoft International,美國),設計擴增16S rRNA和luxS基因的引物(表1)。引物由上海生工生物有限公司合成。
1.3.3相對半定量測定luxS基因表達
以逆轉錄得到的單鏈cDNA為模板,按照mlRNA selective PCR kit (TAKARA,日本)說明書進行相對半定量分析。首先調節內參16S rRNA基因PCR時的cDNA加樣量,通過較少循環數PCR擴增(85℃ 1 min;54.8℃ 1 min;72℃1min,25個循環),使PCR產物濃度在a、b、c、d 4點相同(利用GeneTools from SynGene軟件(Syngene,英國)測定瓊脂糖凝膠中PCR產物的濃度),記錄a、b、c、d 4點相應的模板cDNA加樣量。然后,按照上一步相同的cDNA加樣數加量,同樣利用較少循環數PCR擴增(90℃l min;52.5 ℃ 1 min;72℃lmin,25個循環)a、b、c、d 4點的luxS基因,最后通過瓊脂糖凝膠電泳與GeneTools fromSynGene軟件測定和分析luxS基因在a、b、c、d 4點相對表達量。
1.4抑菌試驗
Lb.plantarum AY01種子液經活化培養后,按40‰ (V/V)接種到新鮮的MRS液體培養基中,30℃靜止培養至a、b、c、d 4點,9000r/min離心2min,收集上清液和菌體。菌體用1mL無菌水懸浮后用細胞破碎儀破碎(程序為“在25℃下,按60%總功率,超聲開3.5s,關9s”處理5min)。上清液和菌體破碎液通過牛津杯法進行抑菌實驗:首先,將滅菌好的2%水瓊脂倒入直徑為90mm的平板中,凝固后,將4個無菌牛津杯垂直放置在同一個平板上;然后,將含有約l.Oxl06cfu/mL食源性病原菌的培養基小心倒人上述平板中,凝固后,用滅菌鑷子夾出牛津杯;最后,分別取250μL上清液和菌體破碎液加入到牛津杯孔中,30℃過夜培養后(約15h),用尺子按垂直測量的方法測定抑菌圈直徑,取兩次垂直測量值的平均數。每個樣品做3個平行平板,3個平板的平均值作為最終的抑菌值[20]。
2結果
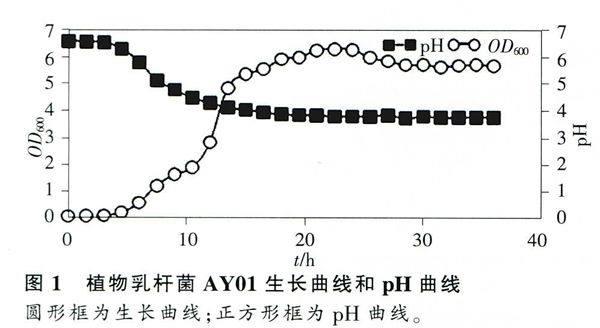
2.1生長曲線和pH相關曲線
以培養時間為橫坐標,以OD600和pH值為縱坐標,制作Lb.plantarum AY01的生長曲線和產酸曲線,結果顯示AY01具有典型微生物生長曲線特征(圖1)。在O~3h,AY01的OD600≈0,說明AY01在該時間段處于延滯期;隨后AY01進入指數期,3~4.5h處于指數早期,4.5~12h處于指數中期,12~18h處于指數晚期;大約從18h開始,AY01生長進入穩定期,持續時間約為22h(18~36h);最后OD600有下降的趨勢,說明AY01開始進入衰亡期(圖1)。Lb.plantarum AY01培養液的pH值具有和生長曲線完全相反的特征(圖1)。在0~3h,pH穩定維持在6.5;在3~18h時間段,pH值處于迅速下降狀態,由6.29下降到3.86;18h后,AY01培養液pH處于一個穩定的低pH值狀態,pH≈3.8。
2.2生長階段luxS基因表達分析
2.2.1RNA提取與分析
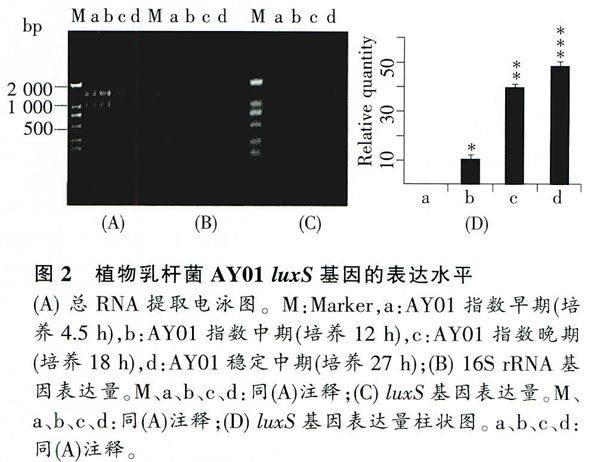
2%瓊脂糖凝膠電泳結果顯示,a、b、c、d 4點沒有降解,且都具有清晰的兩條帶,根據Marker可判斷該兩條帶分別為23S、16S rRNA(圖2A)。此外,利用分光光度計測定a、b、c、d 4點的A260/A280比值分別為1.876、1.959、1.871、1.856,說明有微量DNA污染。因為mRNA selective PCR kit能去除少量DNA的干擾,所以對該RNA不需DNA酶處理,直接進行逆轉錄反應。
2.2.2 16S rRNA基因內參設定
運用半定量PCR方法擴增內參16S rRNA基因,確定a、b、c、d 4點內參16S rRNA PCR擴增量相同時模板cDNA的加樣量。通過結果分析得知,當a、b、c、d 4點分別加1.26、1.52、1.05、0.94 μL模板后,其PCR擴增條帶亮度基本一致(圖2B)。
2.2.3luxS基因表達分析
根據內參設定結果,在luxS基因PCR擴增過程中,a、b、c、d 4點分別加入1.26、1.52、1.05、0.94 μL的模板cDNA,經25個循環后,發現Lb.plantarum, AY01 luxS基因的表達水平隨生長時間的增加呈遞增趨勢,即指數早期沒有表達,指數中期出現表達,相對表達量約為10,指數晚期表達量明顯增加,相對表達量約為39.5,穩定期表達達到最大值,相對表達量約為48.5 (圖2C、D)。
通過DPS數據處理系統一單因素試驗統計分析,結果顯示luxS基因a、b、c、d 4點表達量相互之間存在顯著性差異(n=3,P<0.0)(圖中*的多少表示它們之間具有顯著性差異)。
2.3抑菌結果
Lb.plantarum AY01上清液對EHEC 0157:H7、L.monocytogenes、S.aureus均有抑菌效果,且具有隨著生長時間的增加呈增大變化的特征(表2)。另外,AY01菌體在a、b、c、d 4點對3種病原菌(EHEC 0157:H7、L.monocytogenes、S.aureus)都沒有抑菌效果(結果未顯示1,說明AY01的抑菌物質可能主要是分泌型代謝產物或酶。
注:a:AYOl指數早期(培養4.5 h),b:AYOl指數中期(培養12h);c:AY01指數晚期(培養18h);d:AYOl穩定中期(培養27h)。
Notes:a:AYOl early exponential phase (cultured 4.5h);b:AY01 middle exponential phase (cultured 12h);c:AYOl lateexponential phase (cultured 18h);d:AYOl middle stationaryphase (cultured 27h).
3討論
早在2010年,我們實驗室從云南石林路南鮮羊奶中分離得到了一種益生效果較好的乳酸菌,16S rRNA基因序列分析證明其是植物乳桿菌,我們將該菌株命名為Lb. plantarum AY01。目前,AY01的全基因組序列已經測序完成[19]。本文研究發現,Lb.plantarum AY01 luxS基因表達在指數中期出現,隨著生長的進行,表達水平一直增加,至穩定中期達到最大值。Moslehi-Jenabian等[13】在研究AI-2/LuxS QS系統與Lactobacillus酸適應關系中發現了相似結果,即隨著Lactobacillus生長的進行,細胞數逐漸增多,造成的結果為luxS基因的表達水平和其合成產物-QS信號分子AI-2活性的逐漸增加,并于穩定期達到最大值。基于Lb.plantarum AY01的生長曲線和luxS基因表達相一致的趨勢(當細胞生長到一定數量時luxS基因才出現表達,并且隨著生長的進行,溶液中細胞濃度的增加,luxS基因表達也相應增加)以及luxS基因編碼產物能合成QS信號分子AI-2的背景知識[11],說明luxS基因與Lb.plantarum AY01QS密切相關。在AYOI的生長過程中,luxS基因表達水平的增加是因為AY01細胞數量的增加引起的,還是隨著合成的AI-2分子數目越來越多,最終啟動QS系統,而該QS系統又能促進luxS基因的表達這樣一種模型,還需要更深一步的研究,這也是我們下一步研究的方向。
牛津杯抑菌實驗結果顯示,Lb.plantarumAY01上清液的抑菌效果隨著生長進行也呈現增強變化。這與Lee[2l]的一本書中講到的類似,即乳酸菌與E.coli、L.monocytogenes、S.aureus等病原菌共培養時,隨著生長進行乳酸菌的數量明顯增加,而病原菌的數量明顯減少并最終消失。在本實驗中,隨著Lb.plantarum AY01生長的進行,細胞的濃度越來越高,luxS基因的表達量也隨之增加;在該過程中,AY01上清液對3種常見食源性病原菌的抑菌效果(EHEC 0157:H7、L.monocyto-genes、S.aureus)也越來越明顯,說明AYOl luxS基因表達可能與其抑菌相偶聯;此外,由產酸曲線得知,隨著AY01生長的進行,pH值呈現顯著下降,于18h后達到一個穩定的低pH值狀態,說明在該過程中,AY01的產酸量越來越多。由我們的實驗結果以及Moslehi-Jenabian[3].Diep[l5,16]和Man等[17,18]的前期研究,我們可以做如下推測:Lb.pLantarum AY01 luxS基因不但與QS密切相關,而且可能與發揮抑菌效果也有很大的關系,并且這些抑菌物質很可能是某些酸類或者細菌素等。Lb. plantarum AY01可能通過AI-2/LuxS QS系統調節著酸或者細菌素等物質的合成和分泌,最終介導著其對常見食源性病原菌的抑制。
參考文獻(References):
[1]劉振民,駱承庠,乳酸菌發酵劑生物工程技術[J]食品與發酵工業(LIU Zhen-min, LUO Cheng-xiang. Progress in biotech—nology of lactic acid bacteria starter culters[J].Food and Fer-mentation Industries), 2000, 26(4): 68-72.
[2]劉振民.乳酸菌益生特性及應用[C].杭州:中國奶業協會(LIUZhen-min. Lactic acid bacteria probiotic properties and appli-cations[C]. Hangzhou: China Dairy Industry Association), 2009199-201.
[3]MOSLEHI-JEIANENAB S,VOGENSEN F,JESPERSEN L.The quorum sensing luxS gene is induced in Lactobacillusacidophilus NCFM in response to Listeria mnnocytogenes[J].In-ternational Journal of Food Microbiology, 2011, 149(3): 269-273.
[4]KIM Y,OH S,PARK S,et al.Lactobacillus acidophilus re-duces expression of enterohemorrhagic Escherichia coli O157:H7 virulence factors by inhibiting autoinducer-2-like activity[J].Food Control,2008, 19(11): 1042-1050.
[5]熊濤,宋蘇華,黃濤,等,植物乳桿菌NCU116抑菌性能的研究[J].食品與發酵工業(XIONC Tao, SONC Su-hua,HUANGTao, et al.Antibacterial experimenb of Lactobacillus plan一tarum NCU116[J]. Food and Fermentation Industries), 2012, 38(6): 97-101.
[6] 熊駿.云南傳統發酵豆豉由來乳酸菌中主要有機酸的測定及其抑菌效果研究[D].昆明:昆明理工大學(XIONC Jun.Analy-sis of major organic acids from Laetic acid bacteria isolatedfrom Yunnan traditional fermented Douchi and study on bacteriostasis[D]. Kunming: Kunming Univcrsity of Science and Tech-nology), 2011.
[7]DENG Y Y,WU J E,TAO F,et al Listening to a new lan-guage: DSF-based quorum sensing in gram-negative bacteria[J].Chemical Reviews, 2011,111(1):160-173
[8]趙麗萍,金黃色葡萄球菌AI-2群體感應系統的調控[D].合肥:中國科學技術大學 (ZHANG Li-ping. AI-2 quorum sensing regulation in Staphylococcus aureus[D]. Hefei: University ofScience and Technology of China), 2010.
[9]CHEN X. SCHAUDER S, POTIER N, et al. Structural identi-fication of a bacterial quorum sensing signal containing boron[J].Nature, 2002, 415(6871):545-549.
[10]de KEERSMAECKER S C J, SONCK K, VANDERLEYDEN J.Let LuxS speak up in AI-2 signaling[J]. Trends in Microbiolo-gy, 2006,14(3):114-119.
[11]BUCK B L. AZCARATE-PERIL M A, KLAENHAMMER T R.Role of autoinducer-2 0n the adhesion ability of Lactobacillusacidophilus[J]. Journal of Applied Microbiology, 2009, 107(1):269-279.
[12]di CAGNO R, de ANGELIS M, CALASSO M, et al. Proteomicsof the bacterial cross-talk by quorum sensing[J]. Journal of Pro-teomics,2011,74(1):19-34.
[13]MOSLEHI-JENABIAN S, GORI K, JESPERSEN L.AI-2 sig-naling is induced by acidic shock in probiotic strains of Lacto-bacillus spp[J]. International Journal of Food Microbiology, 2009,135(3):295-302.
[14]LEBEER S. CSLAES I J J, VERHOEVEN T L A, et al. Im-pact of luxS and suppressor mutations on the gastrointestinaltransit of Lactobacillus thamnosus GG[J]. Applied and Environ-mental Microbiology, 2008, 74(15):4711-4718.
[15]DIEP D B, MATHIESEN G, EIJSINK V G H, et al. Use ofLactobacilli and their pheromone-based regulatory mechanismin gene expression and drug delivery[J]. Current Pharmaceuti-cal Biotechnology, 2009, 10(1):62-73.
[16]DIEP D B, STRAUME D, KIOS M, et al. An overview of themosaic, bacteriocin pln loci from Lactobacillus plantarum[J].Peptides, 2009, 30(8):1562-1574.
[17]MAN L L, MENGX C, ZHAO R H. Induction of plantaricinMG under co-culture with certain lactic acid bacterial strainsand identification of LuxS mediated quorum sensing system inLactobacillus plantarum KLDSl. 0391[J]. Food Control, 2012,23(2):462-469.
[18]MAN L L, MENG X C, ZHAO R H, et al. The role ofplNC8HK-plnD genes in bacteriocin production in Lacto-bacillus plantarum KLDSl. 0391[J]. International Dairy Journal,2014,34(2):267-274.
[19]LI X R, GONG F M, ZHENG H J, et al. Draft genome se-quence of Lactobacillus plantarum strain AY01 from freshgoatso' milk[J]. Genome Announcements, 2013,1(5):1.
[20]錢存柔,黃儀秀.微生物學試驗教程[M].北京:北京大學出版社
(QIN Cun-rou, HUANG Yi-xiu. Microbiology Course[M]. Bei-jing: Peking University Press), 1999.176-182.
[21]LEE C H. Fermebtation Technology in Korea[M]. Seoul: KoreaUniversity Press, 2001.
猜你喜歡
甘肅教育(2016年22期)2016-12-20 22:28:05
藝術評鑒(2016年17期)2016-12-19 19:12:28
青年文學家(2016年17期)2016-12-17 20:26:03
中學課程輔導·教師教育(上、下)(2016年17期)2016-12-17 18:11:48
成才之路(2016年35期)2016-12-12 12:08:34
中學課程輔導·教師教育(上、下)(2016年19期)2016-12-07 20:59:07
藝術科技(2016年9期)2016-11-18 18:24:22
藝術科技(2016年9期)2016-11-18 16:48:09
校園英語·上旬(2016年10期)2016-11-16 19:03:00
人間(2016年28期)2016-11-10 00:08:22
