滇龍膽GrHMA基因的克隆和原核表達(dá)
2014-04-29 11:51:28王彩云李富生李濤李彩霞張曉東王元忠
生命科學(xué)研究 2014年3期
王彩云 李富生 李濤 李彩霞 張曉東 王元忠

摘要:HMA蛋白(heavy metal transporting ATPase)是一種在植物中廣泛存在的多功能蛋白。根據(jù)滇龍膽轉(zhuǎn)錄組GrHMA基因序列,設(shè)計(jì)特異性引物,通過RT-PCR擴(kuò)增GrHMA基因序列,并進(jìn)行TA克隆、測序及序列分析;構(gòu)建原核表達(dá)載體pGEX-4T-1-GrHMA,轉(zhuǎn)入E.coli Rosetta(DE3)中,并在37℃、1.Ommol/L IPTG下成功誘導(dǎo)表達(dá)。序列分析表明,GrHMA基因是HMA超家族的成員;GrHMA氨基酸序列系統(tǒng)發(fā)育分析表明,CrHMA與TcHMA處于同一進(jìn)化枝。SDS-PAGE結(jié)果表明所表達(dá)蛋白與預(yù)期蛋白大小一致。這些結(jié)果為GrHMA蛋白的進(jìn)一步純化及結(jié)構(gòu)和功能的研究奠定基礎(chǔ)。
關(guān)鍵詞:滇龍膽;GrHMA基因;克隆;原核表達(dá)
中圖分類號:Q786
文獻(xiàn)標(biāo)識碼:A
文章編號:1007-7847(2014)03-0211-07
滇龍膽(Gentiana rigescens)為龍膽科(Gen-tianaceae)龍膽屬多年生宿根草本植物,是我國特有物種,主要分布于云南、貴州、四川、廣西等省,其中云南是主要產(chǎn)區(qū)。野外主要生長在海拔1100~3000m雜木林下、荒坡地、山谷灌木叢旁[1]。滇龍膽在云南有悠久的藥用歷史[2],被《中國藥典》2010年版收錄,為傳統(tǒng)中藥材龍膽的植物主要來源之一[3]。目前,國內(nèi)外對滇龍膽的研究主要集中在栽培、農(nóng)藝性狀、化學(xué)成分研究、礦質(zhì)元素測定等方面[4-6],而對滇龍膽的重金屬轉(zhuǎn)運(yùn)蛋白及其逆境脅迫和抗性研究未見報(bào)道。
重金屬ATP酶(heavy metal transporting AT-Pase,HMA),又稱為P1B-ATPase,其能選擇性地運(yùn)輸Cu+、Cu2+、Zn2+和CO2+等必須的金屬離子,還能轉(zhuǎn)運(yùn)Cu2+、Cd22+和Pb2+等重金屬離子[7];因其含有一個(gè)保守的內(nèi)膜基序(CPx:Cys-Pro-Cys/His/Ser),又稱為CPx-ATPase[8,9]。植物HMA除參與重金屬離子的穩(wěn)態(tài)外,還涉及植物對重金屬的吸收、轉(zhuǎn)運(yùn)、解毒和富集等方面[10,ll]。HMA基因在植物中廣泛存在,如在低等植物綠藻(Chlamydomonas rein-hardtii)中克隆到3個(gè)基因(CrHMA1~3),紅藻(Cyanidioschizon merolae)中克隆到2個(gè)基因(CmHMA1,CmHMA2)。單子葉植物水稻(Oryzasativa)基因組中發(fā)現(xiàn)9個(gè)HMA基因(OsHMAl~9),大麥(Hordei vulgaris)中有10個(gè)HMA基因(HvHMAl-HvHMA10)[11~13]。于擬南芥(Arabidop-sis thaliana)中克隆到8個(gè)HMA基因(AtHMA l~8) [12,14],大豆(Glycine max)基因組中發(fā)現(xiàn)9個(gè)HMA基因(GmHMAl~GmHMA9)[15]。此外,Southron、Pa-poyan等16,17]從甘藍(lán)型油菜(Brassica napus)克隆到了P1B-ATPase基因BnRANl(HMA7);Courbot等[18]從重金屬Zn/Cd超富集植物鼠耳芥(Arabidopsishaller)中克隆到AhHMA4;Cobbett、Bernard等[8,19]從遏藍(lán)菜(Thlaspi caerulescens)中克隆到TcHMA4基因。植物基因組中有多個(gè)HMA基因存在,其蛋白在植物的根、莖、葉和細(xì)胞中的葉綠體、內(nèi)質(zhì)網(wǎng)、高爾基體、液泡和質(zhì)膜上均有定位[20]。根據(jù)重金屬的底物特異性可以將P1B-ATPase分為兩個(gè)亞類:Zn2+/CO2+/Cd2+/Pb2+P1B-ATPase(Zn亞類)和Cu+/Ag+P1B-ATPase(Cu亞類),其中動物只有Cu亞類[11,21]。植物中則包含這兩個(gè)亞類,如擬南芥AtHMAl-4屬于Zn亞類,而AtHMA5-8屬于Cu亞類[21];水稻OsHMAl-OsHMA3屬于Zn亞類,OsHMA4-OsHMA9屬于Cu亞類[13]。
雖然HMA蛋白在植物中廣泛分布,但目前的研究仍僅局限于擬南芥,在重金屬超富集植物中沒有進(jìn)行系統(tǒng)全面的研究,而在藥用植物滇龍膽中至今未有報(bào)道。本研究根據(jù)3年生滇龍膽的轉(zhuǎn)錄組中GrHMA基因序列設(shè)計(jì)特異性引物,從滇龍膽幼葉中提取RNA,反轉(zhuǎn)錄為cDNA,進(jìn)而擴(kuò)增到GrHMA基因序列,并進(jìn)行TA克隆、測序和序列分析;構(gòu)建GrHMA基因原核表達(dá)載體,在大腸桿菌Rosetta中進(jìn)行表達(dá),使用SDS-PAGE檢測其蛋白表達(dá)情況。結(jié)果顯示GrHMA基因?yàn)镠MA基因家族的成員,在37℃、1.0mmol/L IPTG誘導(dǎo)下成功表達(dá),這將為滇龍膽GrHMA蛋白的純化、結(jié)構(gòu)和功能研究奠定基礎(chǔ),也為植物、動物、微生物中HMA的研究提供參考。
1材料與方法
1.1植物材料
供試材料滇龍膽(Gentiana rigescens Franch.ex Hemsl.)種植于云南省農(nóng)科院藥用植物研究所種質(zhì)資源圃(25°08'04.50"N,102°46'15.05"E)。栽培地海拔1942m,年平均氣溫14.7℃,年平均降水量980~1050mm,極端高溫30.4℃,極端低溫-0.2℃。試驗(yàn)材料為滇龍膽長勢良好的幼葉。
1.2菌株、載體、酶等主要試劑
工程菌E.coli Trans 5α、E.coli Rosetta(DE3)、RNAiso for Polysaccharide-rich Plant Tissue Reagent、反轉(zhuǎn)錄試劑盒、限制性內(nèi)切酶BamH I和Xho I及異丙基-β-D-硫代吡喃半乳糖苷(IPTG)、X-gal等均購買于寶生物工程(大連)有限公司;高純質(zhì)粒小量制備試劑盒(離心柱型)、多功能DNA純化回收試劑盒均購自北京百泰克生物技術(shù)有限公司;質(zhì)粒pGEX-4T-l由玉溪師范學(xué)院分子生物學(xué)實(shí)驗(yàn)室保存;引物由上海捷瑞生物工程有限公司合成,測序由生工生物工程(上海)股份有限公司完成。
1.3引物設(shè)計(jì)
根據(jù)3年生滇龍膽轉(zhuǎn)錄組中GrHMA基因序列和原核表達(dá)載體pGEX-4T-l多克隆酶切位點(diǎn),設(shè)計(jì)一對特異引物GrHMABamH I-F:5'一GG -ATCCATGTCTATGGTGGAAGTTTTGG-3':GrHMAXho I -R: 5'-CTCGAGTCACATAATAGAACAAG-CATGGG-3'。
1.4滇龍膽葉片總RNA的提取及GrHMA基因的擴(kuò)增
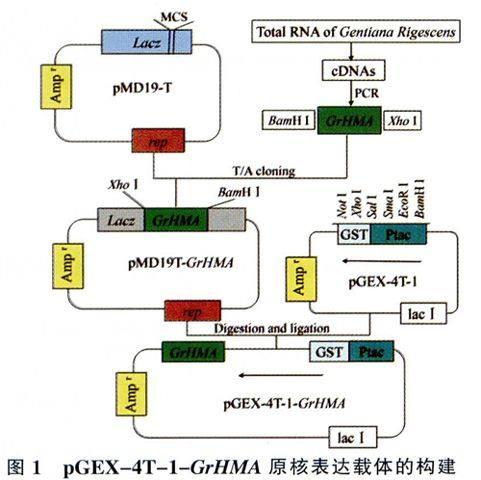
采用植物RNAiso for Polysaccharide-rich PlantTissue提取試劑盒,按照說明書提取滇龍膽葉片的總RNA,參照Reverse Transcriptase M-MLV(RNase H-)試劑盒說明書反轉(zhuǎn)錄為cDNA,-20℃保存?zhèn)溆谩R詂DNA為模板進(jìn)行PCR擴(kuò)增,反應(yīng)條件為:94℃ 3min;94℃ 30s,54℃ 30s,72℃30s,30個(gè)循環(huán);72℃延伸7min。利用BioTekeCorporaltion高純質(zhì)粒小量制備試劑盒回收RT-PCR產(chǎn)物,隨后將其連接到pMD19-T載體上,轉(zhuǎn)化E.coli Trans5a感受態(tài)細(xì)胞,涂布于添加氨芐青霉素(Amp)(100mg/L)、IPTG、X-gal的LB平板上37℃培養(yǎng)12~16h后隨機(jī)挑選陽性克隆搖菌并提取質(zhì)粒,經(jīng)PCR檢測和酶切驗(yàn)證正確后送出測序,即可獲得pMD19T-GrHMA重組載體。使用BarnH I和Xho I雙酶切pMD19T-GrHMA重組質(zhì)粒和pGEX一4T-l載體,回收目的基因和載體片段,按1:4(摩爾比)混合后經(jīng)Ligation SolutionI連接并轉(zhuǎn)化E.coli Trans Sa感受態(tài)細(xì)胞,涂布于添加100mg/L Amp的LB固體平板。次日隨機(jī)挑選陽性克隆搖菌并提取質(zhì)粒,經(jīng)PCR檢測和酶切驗(yàn)證正確后測序,即可獲得pGEX-4T-1-GrHMA原核表達(dá)載體(圖1)。
1.5GrHMA的生物信息學(xué)分析
利用NCBI網(wǎng)站上的BLAST程序?qū)rHMA蛋白進(jìn)行分析,應(yīng)用DNAMAN推測并比對氨基酸序列,利用在線軟件預(yù)測GrHMA蛋白的相對分子質(zhì)量、氨基酸含量、等電點(diǎn)、二級結(jié)構(gòu)、三級結(jié)構(gòu)和稀有密碼子等;應(yīng)用軟件Mega4.0構(gòu)建系統(tǒng)進(jìn)化樹。
1.6pGEX-4T-1-GrHMA重組質(zhì)粒在大腸桿菌Rosetta中的表達(dá)
利用熱激法將pGEX-4T-l-GrHMA重組質(zhì)粒轉(zhuǎn)化大腸桿菌Rosetta( DE3)感受態(tài)細(xì)胞,挑取單菌落并接種于4mL含100mg/L Amp的LB液體培養(yǎng)基中,37℃ 200r/min培養(yǎng)過夜。次日以l:100接種到新的不含Amp的LB液體培養(yǎng)基中,37℃200r/min培養(yǎng)至OD6≈0.6~0.8,在37℃條件下,加入IPTG(終濃度為lmmol/L)進(jìn)行誘導(dǎo)表達(dá),并以pGEX-4T-l質(zhì)粒轉(zhuǎn)化菌(含1mmol/L IPTG)作對照。誘導(dǎo)0、1、2、4、6、8、18h后分別收集2mL菌液。常溫13000r/min離心lmin,棄上清,加入20μL5xSDS-PAGE蛋白上樣緩沖液,100μL Milli-Qwater,震蕩混勻,煮水沸1 5min。室溫13000r/min離心5min,取20μL蛋白樣品上樣,進(jìn)行SDS-PAGE(12%分離膠和4%濃縮膠)電泳檢測。
2結(jié)果與分析
2.1滇龍膽GrHMA基因的克隆與分析
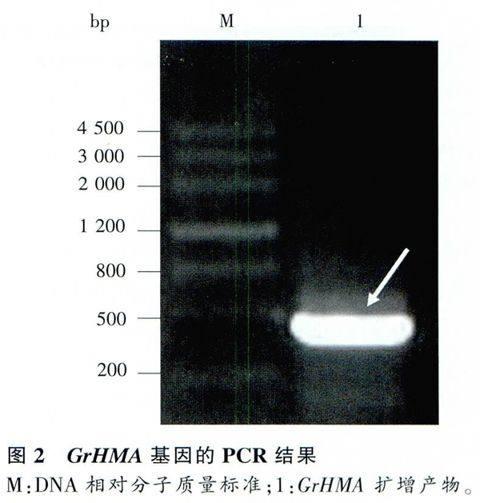
以滇龍膽cDNA為模板擴(kuò)增出的PCR結(jié)果表明,擴(kuò)增片段在500bp附近,是預(yù)期的目的片段(圖2)。pMD19T-GrHMA經(jīng)酶切驗(yàn)證連接正確,有明顯的、單一的目的條帶,且目的條帶與預(yù)期條帶的大小吻合。
cDNA序列測序結(jié)果分析表明,滇龍膽GrHMA基因(GenBank登錄號為KF922376)全長429bp,編碼142個(gè)氨基酸。
使用在線軟件ExPASy ProtParam tool(http://web.expasy.org/protparam/)對GrHMA蛋白進(jìn)行分析,結(jié)果顯示GrHMA蛋白的相對分子質(zhì)量為15.56kD,理論等電點(diǎn)為6.58,化學(xué)方程式為:C705H1069N179O205S7,不穩(wěn)定指數(shù)為31.78,脂肪族指數(shù)為71.14,總平均疏水性(grand average of hy-dropathicity,GRAVY)為-0.182,半衰期為30h。通常依據(jù)蛋白的GRAVY值來預(yù)測蛋白的疏水性,GRAVY值在2到-2之間,若為負(fù)值,則為親水蛋白;若為正值,則為疏水蛋白。GrHMA蛋白的GRAVY值為-0.182,說明GrHMA為親水蛋白。
通過使用ExPASy ProtParam tool對GrHMA蛋白的氨基酸含量進(jìn)行預(yù)測,結(jié)果顯示GrHMA蛋白含有20種氨基酸,其中丙氨酸(Ala)、纈氨酸(Val)的含量最高,為10.6%,其次是甘氨酸(Gly)、絲氨酸(Ser),為7.7%;谷氨酰胺(Gln)、色氨酸(Trp)含量最低,僅為0.7%。
采用InterProScan在線工具對GrHMA蛋白的保守結(jié)構(gòu)域進(jìn)行預(yù)測。結(jié)果顯示GrHMA蛋白有6個(gè)保守結(jié)構(gòu)域(圖3),主要是HMA(heavy met-al-associated domain,HMA)結(jié)構(gòu)域(IPR006121),分別為位于6~63位HMA( PF00403)結(jié)構(gòu)域、3~67位HMA-2(PS50846)結(jié)構(gòu)域和1~70位HMA(heavy metal-associated domain)結(jié)構(gòu)域(SSF55008)。另外還有在IPR中沒有明確分類的結(jié)構(gòu)域,主要為位于2~69位未明確描述的結(jié)構(gòu)域(3.30.70.100)、1~70位Copper Transport Protein Atoxl-Related結(jié)構(gòu)域(PTHR22814)和1~70位OS04G0401000 Pro-tein(PTHR22814:SF44)結(jié)構(gòu)域。
選擇GrHMA氨基酸序列,在NCBI中進(jìn)行BLASTp分析,結(jié)果表明滇龍膽GrHMA蛋白屬于HMA超家族(圖4)。
GrHMA氨基酸序列與NCBI中經(jīng)比對后一致性較高的可可(Theobroma cacao)、擬南芥(Ara-bidopsis thaliana)、蓖麻(Ricinus communis)、蒺藜狀苜蓿(Medicago truncatula)HMA氨基酸序列進(jìn)行比對,結(jié)果表明GrHMA蛋白與可可TcHMA同源性最高(74%),其次是擬南芥AtHMA,同源性為70%,與蒺藜狀苜蓿HMA蛋白的同源性較低(39%),見表1。
根據(jù)推測的氨基酸序列進(jìn)行的系統(tǒng)發(fā)育分析,結(jié)果表明GrHMA與TcHMA在同一進(jìn)化枝上,表明GrHMA與TcHMA蛋白的親緣關(guān)系較近(圖5)。GrHMA與NCBI中HMA已知序列進(jìn)行多序列比對,結(jié)果表明滇龍膽GrHMA蛋白保守性較強(qiáng),與已知序列存在較高的同源性(圖6)。
利用ExPASy SignalP 4.0 Server在線軟件(http://www.cbs.dtu.dk/services/SignalP/)對GrHMA蛋白進(jìn)行分析,未發(fā)現(xiàn)信號肽,表明GrHMA不是分泌蛋白。
利用在線軟件Expasy中的TMHMM工具(http://www.cbs.dtu.dk/services/TMHMM-2.0/)預(yù)測GrHMA蛋白的跨膜螺旋區(qū),結(jié)果表明GrHMA蛋白不是膜蛋白,無跨膜螺旋區(qū)。
使用SSpro在線軟件(http://scratch.proteomics.ics.uci.edu/)分析GrHMA的二級結(jié)構(gòu)。結(jié)果顯示該蛋白二級結(jié)構(gòu)中無規(guī)則卷曲(C)占61.98%,β-折疊(E)占17.6%,α-螺旋(H)占20.42%。
利用CPHmodels 3.2 Server在線軟件(http://www.cbs.dtu.dk/services/CPHmodels/)預(yù)測GrHMA蛋白的三級結(jié)構(gòu)(圖7),從圖中可以看到GrHMA的三級結(jié)構(gòu)形成了多個(gè)無規(guī)則卷曲,整個(gè)三維結(jié)構(gòu)呈“口袋”狀,表明該蛋白可能通過該“口袋”與金屬離子和DNA結(jié)合。
用ProtFun在線軟件基于已知的具有相似功能蛋白的搜索和比對,對GrHMA編碼的蛋白質(zhì)進(jìn)行功能預(yù)測,結(jié)果顯示GrHMA蛋白參與復(fù)制與轉(zhuǎn)錄、能量代謝、翻譯、轉(zhuǎn)錄調(diào)控的可能性遠(yuǎn)高于其他功能,其可能性分別為0.204、0.184、0.167和0.118;另外其參與金屬離子轉(zhuǎn)運(yùn)的可能性為0.009,這些數(shù)據(jù)能為GrHMA蛋白的后續(xù)研究提供參考。
對GrHMA基因進(jìn)行稀有密碼子分析(http://molbiol.edu.ru/eng/scripts/01_11.html),結(jié)果表明GrHMA基因中不含稀有密碼子,因此可選用Rosetta菌或BL21進(jìn)行原核表達(dá)。
2.2原核表達(dá)重組質(zhì)粒的鑒定
利用BamH I和Xho I雙酶切pGEX-4T-l一GrHMA重組質(zhì)粒,可檢測到約429bp的片段(圖8),表明GrHMA基因已插入pGEX-4T-1載體中。對pGEX-4T-1-GrHMA重組質(zhì)粒進(jìn)行測序,結(jié)果顯示連入原核表達(dá)載體中的基因片段與目的序列一致,酶切位點(diǎn)處序列連接無誤,沒有移碼及堿基突變等現(xiàn)象出現(xiàn),表明已獲得正確的CrHMA原核表達(dá)載體。
2.3 SDS-PAGE分析
將pGEX-4T-l-GrHMA重組質(zhì)粒轉(zhuǎn)化大腸桿菌Rosetta(DE3)并誘導(dǎo)表達(dá)。37℃、IPTG終濃度為1mmol/L時(shí)分別誘導(dǎo)0、1、2、4、6、8、18h后,提取大腸桿菌總蛋白進(jìn)行SDS-PAGE檢測。結(jié)果顯示,插入外源片段的pGEX-4T-1一GrHMA重組質(zhì)粒經(jīng)IPTG誘導(dǎo)后,在預(yù)期蛋白相對分子質(zhì)量41.56kD(包含GST蛋白)處有1條蛋白帶,而未誘導(dǎo)的pGEX-4T-l對照質(zhì)粒和轉(zhuǎn)化重組質(zhì)粒均未出現(xiàn)該條帶,表明pGEX-4T-1-GrHMA重組質(zhì)粒在大腸桿菌Rosetta中誘導(dǎo)表達(dá)了GrHMA蛋白。誘導(dǎo)時(shí)間為8h時(shí),蛋白表達(dá)量已很大,18h時(shí)達(dá)最大(圖9),表明相同條件下,一定時(shí)間范圍內(nèi),隨著誘導(dǎo)時(shí)間的延長,蛋白表達(dá)量逐漸增加。
3討論
HMA是一個(gè)在植物中廣泛存在的蛋白家族,具有參與葉綠體Cu運(yùn)輸、Cu穩(wěn)態(tài)、毒性重金屬轉(zhuǎn)運(yùn)等功能。植物中有關(guān)HMA蛋白純化的報(bào)道較少,其確切功能還有待進(jìn)一步研究。研究表明HMA可以調(diào)節(jié)植物穩(wěn)態(tài),具有重金屬轉(zhuǎn)運(yùn)、吸收、富集等作用。如Mills等[22]研究發(fā)現(xiàn)擬南芥AtHMA4的T-DNA插入突變體對Cd、Zn離子濃度的升高敏感,AtHMA4在野生型酵母(Saccharomyces cere-visiae)中的異源表達(dá)可提高其對Cd離子的抗性,還能使Zn敏感型大腸桿菌zntA突變體存活;Andres-Colas等[23]研究發(fā)現(xiàn)AtHMA4在擬南芥中過表達(dá)可增加葉片中Zn和Cd的含量;Verret F等[24]研究發(fā)現(xiàn)AtHMA4在野生型酵母和Co超敏感突變體cotl中的表達(dá)能提高酵母對Co的敏感性。Papoyan等[17]研究表明遏藍(lán)菜TcHMA4在酵母中的過表達(dá)可提高其對重金屬離子的耐受性,并使其輸出Cd、Zn、Pb和Cu等重金屬;鼠耳芥AhHMA4在酵母中的過表達(dá)可降低細(xì)胞中Zn和Cd的含量,表明AhHMA4蛋白具有跨膜轉(zhuǎn)運(yùn)Zn和Cd的作用。本研究利用RT-PCR技術(shù),從滇龍膽幼葉中分離到GrHMA基因,序列分析表明GrHMA屬于HMA蛋白超家族。系統(tǒng)發(fā)育分析顯示,GrHMA與SIHMA26位于同一進(jìn)化枝,親緣關(guān)系最近;與CsHMA同源性最低,親緣關(guān)系最遠(yuǎn)。但要明確GrHMA蛋白在滇龍膽中的具體功能,必須從蛋白水平進(jìn)一步研究,而原核表達(dá)是目前最常用的研究蛋白結(jié)構(gòu)和功能的方法。
本研究首次從滇龍膽中克隆到GrHMA基因,成功將其連接到pGEX-4T-l原核表達(dá)載體上并順利在大腸桿菌表達(dá)菌Rosetta(DE3)中表達(dá)到目的蛋白。大腸桿菌中外源目的基因能否順利表達(dá)及表達(dá)效率的高低取決于原核表達(dá)載體、大腸桿菌菌株、誘導(dǎo)溫度、誘導(dǎo)時(shí)間以及誘導(dǎo)劑濃度等。本研究在大腸桿菌Rosetta中進(jìn)行表達(dá),獲得預(yù)期效果,在37℃、60r/min、IPTG終濃度為1mmol/L時(shí),成功誘導(dǎo)了目的蛋白的表達(dá)。
本研究克隆了滇龍膽GrHMA基因,成功在大腸桿菌Rosetta中表達(dá)并獲得融合蛋白。這為GrHMA蛋白結(jié)構(gòu)和功能的深入研究以及進(jìn)一步開發(fā)和利用植物HMA蛋白都具有重要的理論和應(yīng)用價(jià)值。
參考文獻(xiàn)(References):
[1]何廷農(nóng),劉尚武,吳慶如.中國植物志(第62卷)[M].北京:科學(xué)出版社(HE Ting-nong, LIU Shang-wu, WU Qing-ru. TheFlora of China (Vo1.62)[M]. Beijing:Science Press), 1988.1OO-101.
[2]蘭茂.滇南本草(第2卷)[M].昆明:云南人民出版社(LANMao.South Yunnan Medica (Vol.2)[M]. Kunming Yunnan People'sPublishing House),1977,317-318.
[3]國家藥典委員會.中華人民共和國藥典(一部)[s].北京:化學(xué)工業(yè)出版社(National Pharmacopoeia Committee. ChinesePharmacopoeia(a)[S].Beijing. Chemical Industry Press),2010.65.
[4]沈濤,張金渝,楊維澤,等.滇龍膽主要農(nóng)藝性狀的相關(guān)及通徑分析[J].中國中藥雜志(SHEN Tao, ZHANG Jin-yu, YANGWei-ze.et al. Correlation and path analysis on agronomictraits of Gentiana rigescens[J]. China Journal of Chinese Ma-teria Medica), 2011,36(12):1568-1571.
[5]楊美權(quán),張金渝,沈濤,等.不同栽培模式對滇龍膽中龍膽苦苷含量的影響[J].江蘇農(nóng)業(yè)科學(xué)(YANG Mei-quan, ZHANCJin-yu, SHEN Tao, et al.Different cultivation mode influenceon gentiopicroside in Gentian,a rigescens[J]. Jiangsu AgriculturalSciences), 2011(1):287-289.
[6] 沈濤,楊美權(quán),趙振玲,等.滇龍膽中萜類物質(zhì)積累的動態(tài)變化[J]植物學(xué)報(bào)(SHEN Tao, YANG Mei-quan, ZHAO Zhen-ling, et al.Dynamic changes in terpenoid contents in Gentian.arigescens[J]. Bulletin of Botany), 2011,46(6):652-657.
[7]ARGUELLO J M,EREN E,GONzALEZ-GUERRERO M.Thestructure and function of heavy metal transport P1B-ATPases[J].Biometals, 2007,20(3/4):233-248.
[18]BERNARD C,ROOSENS N,CZERNIC P,et al.A novel CPx-ATPase from the cadmium hyperaccumulator Thlaspi caeru-lescens[J]. Febs Letters, 2004,569(1/3):140-148.
[9]SOLIOZ M. VULPE C.CPx-type ATPases:a class of P-typeATPases that pump heavy metals[J]. Trends in Biochemical Bcie-nces,1996,21(7):237-241.
[lO]CLEMENS S.Mecular mechanisms of plant metal toleranceand homeostasis[J]. Planla, 2001,212(4):475-486.
[11]WILLAMS L E,MILLS R F.PIB-ATPases-an ancient familyof transition metal pumps with diverse functions in plants[J].Trends in Plant Science, 2005,10(10):491-502.
[12]PUIG S,ANDRES-COLAS N,GARcfA-MINA A,et al.Cop-per and iron homeostasis in Arabidopsis: responses to metaldeficiencies interactions and biotechnological applications[J].Plant.Cell&Fnvironment, 2007,30(3):271-290.
[13]LEE S,KIM Y Y,LEE Y,et al.Rice P1B-type heavy-metalATPase, OsHMA9, is a metal efflux protein[J]. Plant Physiolo-gy, 2007,145(3):831-842.
[14]HUSSAIN D, HAYDON M J, WANG Y, et al. P-type ATPaseheavy metal transporters with roles in essential zinc homeosta-sis in A rabidopsis[J].The Plant Cell Online, 2004, 16(5):1327-1339.
[15]BERNAL M, TESTILLANO P S, ALFONSO M, et al. ldentifi-cation and subcellular localization of the soybean copper P1B—ATPase CmHMA 8 transporter[J]. Journal of Structural Biology,2007,158(1):46-58.
[16]SOUTHRON J L, BASU U, TAYLOR C J. Complementation ofSaccharomyces cerevisiae ccc2 mutant by a putative P1B-AT-Pase from Brassica napus supports a copper-transporting func-tion[J]. Febs Letters, 2004,566(1/3):218-222.
[17]PAPOYAN A, KOCHIAN L V. Identification of Thlaspicaerulescen,s genes that may be involved in heavy metal hy-peraccumulation and tolerance. Characterization of a novelheavy metal transporting ATPase[J]. Plant Physiology, 2004,136(3):3814-3823.
[18]COURBOT M, WILLEMS G, MOTTE P, et al. A major quanti-tative trait locus for cadmium tolerance in Arabidopsis hallericolocalizes with HMA4, a gene encoding a heavy metal AT-Pase[J]. Plant Physiology, 2007,144(2):1052-1065.
[19]COBBETT C S, HUSSAIN D, HAYDON M J. Structural andfunctional relationships between type 1B heavy metal-trans-porting P-type ATPases in Arabidopsis[J]. New Phytologist, 2003,159(2):315-321.
[20]張玉秀,張媛雅,孫濤,等.植物重金屬轉(zhuǎn)運(yùn)蛋白P1B-AT-Pase結(jié)構(gòu)和功能研進(jìn)展[J].生物工程學(xué)報(bào)(ZHANC Yu-xiu, ZAHNG Yuan-ya, SUN Tao, et al. Structure and functionof heavy metal transporter P1B-ATPase in plant: a review[J].Chinese Journal of Biotechnology), 2010,26(6):715-725.
[21]AXELSEN K B, PALMGREN M G. Inventory of the superfami-ly of P-type ion pumps in Arabidopsis [J]. Plant Physiology, 2001,126(2):696-706.
[22]MILLS R F, KRIJCER G C, BACCARINI P J, et al. Functionalexpression of AtHMA4, a P1B-type ArrPase of the Zn/Co/Cd/pbsubclass[J]. The Plant Journal, 2003,35(2):164-176.
[23]ANDRES-COLAS N, SANCENoN V, RODRfCUEZ-NAVAIR-RO S, et al. The Arabidopsis heavy metal P-type ATPaseHMA5 interacts with metallochaperones and functions in cop-per detoxification of roots[J]. The Plant Journal, 2006,45(2):225-236.
[24]VERRET F, GRAVOT A, AUROY P, et al. Heavy metaltransport by AtHMA4 involves the N-terminal degeneratedmetal binding domain and the C-terminal His11 stretch[J].Febs Letters, 2005,579(6):1515-1522.
