穩定表達Syncytin的EL4細胞系的建立及鑒定
2014-04-29 14:26:10鄭志牛華張桂前范欣高玉紅孫鹥
生命科學研究 2014年3期
鄭志 牛華 張桂前 范欣 高玉紅 孫鹥

摘要:Syncytin是人類內源性逆轉錄病毒W家族的囊膜蛋白。近期研究發現Syncytin與白血病密切相關。為研究Syncytin的生物學功能,我們克隆了人syncytin,并連接到pIRES2-EGFP質粒上,轉化該質粒至感受態大腸桿菌DH5a,挑選陽性克隆進行PCR、酶切電泳和DNA測序鑒定,成功構建了表達syncytin基因的真核表達載體。利用羅氏轉染試劑轉染重組質粒至EL4細胞,并通過G418選擇性培養基篩選,在熒光顯微鏡下觀察細胞中Syncytin表達,用RT-PCR、Western blot檢測Syncytin表達水平,結果顯示我們成功構建了穩定表達人Syncytin的ELA細胞系。穩定表達人Syncytin的EL4細胞系的建立,為進一步研究人Syncytin功能及其與白血病免疫逃逸的關系提供了重要的細胞模型和實驗基礎。
關鍵詞:Syncytin蛋白;pIRES2-EGFP質粒;真核表達載體;EL4細胞;白血病
中圖分類號:Q786
文獻標識碼:A
文章編號:1007-7847(2014)03-0205-06
人內源性逆轉錄病毒(human endogenousretrovlruses,HERV)是幾百萬年前整合到人類基因組中,并以孟德爾方式遺傳至今的逆轉錄病毒的殘余物,約占人類基因組DNA的8%[1]。大多數HERVs在進化過程中由于突變、缺失等的積累失去了編碼能力[2,3],但仍有少數HERVs的開放閱讀框(open reading frames,ORFs)被完整保留了下來。這些完整的ORFs可以編碼逆轉錄病毒的蛋白,在一些特定的組織或分化發育的特定階段表達,可能具有重要的生理意義。Syncytin是HERVs-W家族的囊膜蛋白,主要在胎盤中表達,介導合胞滋養層的形成,維持胎盤正常的生理功能[4]。HERVs已被證實與腫瘤形成密切相關,如乳腺癌[5]、慢性骨髓瘤白血病[6,7],研究發現syncytinmRNA和Syncytin在9種白血病或淋巴瘤細胞系中均有表達[8],具有潛在的研究價值。
Syncytin的373-397殘基是一個具有免疫抑制活性的多肽[9],其在細胞表面的大量表達有利于癌變細胞逃避免疫打擊,同時其具有的融合活性有利于細胞的遷移[10]。本研究擬構建syncytin,真核表達載體,并通過羅氏轉染試劑轉染小鼠淋巴瘤細胞細胞株(EL4),并通過G418抗性培養基篩選出穩定表達Syncytin的細胞系,通過RT-PCR和Western blot分別從mRNA與蛋白水平驗證syncytin在EL4細胞中的表達情況。穩定表達syncytin基因的EL4細胞系的建立為進一步探討syncytin基因異常表達對白血病免疫逃逸的機制提供細胞模型和實驗基礎。
1材料和方法
1.1材料
PIRES2-EGFP質粒、C8166細胞由本實驗室保存,E.coli DH5α感受態細胞、膠回收試劑盒、質粒提取試劑盒,Marker 2500購自天根生物技術公司;EL4細胞購自上海細胞庫;DNA Marker、限制性內切酶(EcoR I、BamH I)、T4 DNA連接酶、Taq DNA聚合酶、SYBR MASTER Mixture、Trizol試劑均購于日本Takara公司;X-tremeGENE HPDNA Transfection Reagent購自羅氏公司;PCR引物及RT-PCR引物由華大基因合成;RPMI1640液體培養基、胎牛血清購自美國Gibco公司;Trizol、Opti-MEM均購于美國Invitrogen公司;Syncytin(H-280)抗體購自美國SANTA公司;Real time PCR儀(日本TAKARA公司);凝膠成像儀(天能公司)攝像分析,Nanodrop分光光度儀(美國Thermo公司),熒光顯微鏡(H本Olympus公司),CO2培養箱(日本三洋),其他試劑均為進口或國產分析純。
1.2pIRES2-syncytin-EGFP重組質粒的構建及鑒定
根據GenBank中的人syncytin基因編碼區序列(核苷酸庫中的編號:AF072506.2)設計了兩套引物,外圍引物(s-1,s-2)和內圍引物(S,A),內圍引物中在上游引物5'端引入EcoR I酶切位點,下游引物5'端引入BamH I酶切位點。S-l:5'-AGGACCCTACCCAGTCATTTTATCT-3'、S-2:5'-GAAGACTTGGGTTTATATCCCGATC-3,:S:5'-CG-GAATTCATGGCCCTCCCTTATCAT-3'、A:5'-CGGGATCCCTAACTGCTTCCTGCTG-3'。培養C8166細胞并提取細胞總RNA,以其為模板利用一步法逆轉錄試劑盒擴增,反應參數為:94℃ 30s、62℃ 30s、72℃ 1min 30s,共40個循環;以擴增產物為模板,用高保真DNA聚合酶PirmeSTARMAX進行PCR擴增帶有酶切位點的目的片段syncytin,擴增片段大小為1.6kb。PCR反應參數為:94℃ 10s、62℃ 5s、72℃ 10s,共35個循環。將PCR產物經瓊脂糖凝膠電泳回收后,用EcoR I/BamH I雙切后純化回收,隨后用T4 DNA連接酶將其與EcoR I /Bam,H I雙切后電泳凝膠回收的PIRES2-EGFP線性質粒進行16℃過夜連接,連接產物轉化DH5a感受態細胞,接種至含卡那霉素(Kan+)的LB瓊脂糖培養板上培養,挑取單菌落接種于LB培養基(含Kan+)中,37℃振搖過夜。用質粒小量提取試劑盒重組質粒,分別進行PCR及EcoR I/BamH I限制性內切酶雙切后電泳鑒定,陽性質粒送樣測序后經DNA序列分析驗證。構建的重組質粒命名為pIRES2-syncytin-EGFP。
1.3穩定表達Syncytin的EL4細胞系的建立
1.3.1EL4細胞的培養和質粒轉染
ELA細胞在含10%胎牛血清的1640培養基中,于37℃、5% CO2條件下培養。參照羅氏轉染試劑盒說明書分別完成重組載體pIRES2 -syn-cytin-EGFP和空載體pIRES2-EGFP的轉染。1)轉染前4h,將處于對數生長期的EIA細胞以lx105密度鋪于24孔板,使細胞匯合度至80%~90%;2)轉染前用無血清無雙抗1640洗滌細胞;3)50μL OPTI-MEM分別稀釋0.5μg DNA,室溫孵育5min;4)50μL OPTI-MEM分別稀釋1.5μL X-tremeGENE HP DNA Transfection Reagent,室溫孵育5min;5)將3)、4)中的液體混合,室溫孵育15~20min;6)將轉染復合物逐滴加入細胞孔板中,輕柔震蕩混勻,放人37℃、5% CO2培養箱培養;7)轉染48h后熒光顯微鏡下觀察熒光。
1.3.2穩定表達Syncytin的EL4細胞系的獲得
將對數生長期的EL4細胞接種于24孔板,每孔5000個,分別加入0、100、200、300、400、500、600、700、800、900、1000、l 10022、1200mg/L的G418,3d換一次含有相同濃度的培養液,篩選14d,以細胞全部死亡的最低G418質量濃度為最佳篩選劑量。重組質粒pIRES2-syncytin-FGFP轉入EI4細胞48h后,進行細胞傳代,并更換含有G418的l640培養液進行篩選,每隔3d更換培養液1次。2周后對篩選的細胞進行有限稀釋法挑選細胞,鋪96孔板,挑出單個細胞并擴大培養,最終獲得具有G418抗性的穩定表達syncytin的EL4細胞系,命名為ELA-Syncytin細胞和空載體質粒穩定轉染細胞。
1.4RT-PCR檢測syncytin轉錄水平的表達
收集EL4-Syncytin細胞和空載體質粒穩定轉染的細胞,按Trizol試劑說明書抽提細胞總RNA,反轉錄制備cDNA,進行PCR反應。利用Prime5.0軟件設計syncytin,上下游引物,上游引物5'-ATCCCCCGCAACTGCTATC-3',下游引物5'-AGACAGTGACTCCAAGTCCTC-3',擴增產物大小為112bp。內參基因GAPDH上游引物5'-GGTGAAGCJTCGGTGTGAACG-3',下游引物5'-CTCGCTCCTGGAAGATGGTG-3',擴增產物大小為233bp。擴增條件為:94℃預變性2min;94℃變性15s;55℃退火15s;72℃延伸20s,共30個循環;72℃繼續延伸5min。對實驗數據進行統計學分析。
1.5Western印記檢測細胞中Syncytin蛋白的表達
收集EL4-Syncytin細胞和空載體質粒穩定轉染的細胞,用含蛋白酶抑制劑的單去污裂解液裂解細胞,冰上放置5min,超聲充分碎裂,4℃,12000r/min離心10min,收集上清,即為細胞總蛋白。用BCA進行蛋白定量,再加入5xloadingbuffer混勻后煮沸5min,離心后-80℃凍存備用。各取20μL蛋白樣品進行SDS-PAGE電泳,電轉使蛋白至PVDF膜上,5%脫脂奶粉室溫震蕩封閉2h,與兔抗人Syncytin抗體(美國Santa公司)孵育過夜,用TBST洗PVDF膜3次,每次5min;再與HRP標記的山羊抗兔的抗體室溫孵育2h,TBST洗PVDF膜3次,每次5min,化學發光劑顯影并保存圖像。
2結果與分析
2.1syncytin基因ORF的擴增
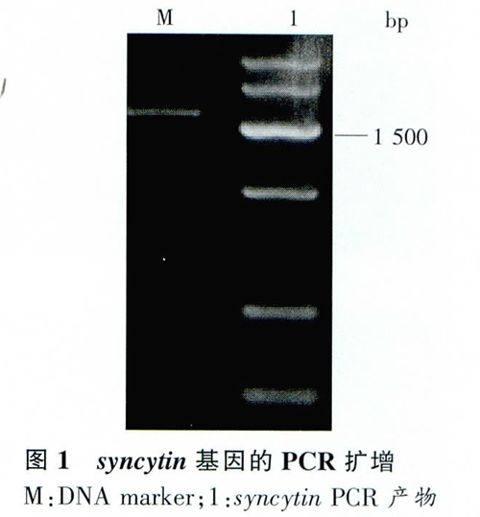
以C8166細胞總RNA為模板進行逆轉錄PCR擴增產物為模板擴增,產物用1%瓊脂糖凝膠電泳鑒定,所得PCR產物片段約為1621bp,與預期的syncytin基因ORF長度一致(圖1)。表明syncytin,基因ORF被成功克隆。
2.2重組質粒的鑒定
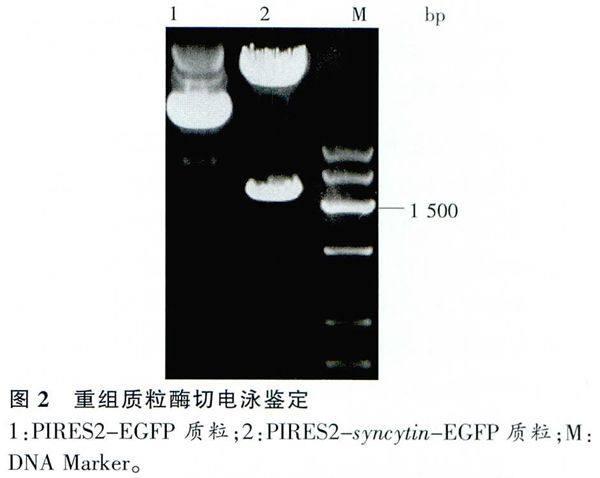
pIRES2-syncytin-EGFP重組質粒與空載體分別EcoR I /BamH I雙切后進行1%瓊脂糖凝膠電泳,可約見1.6kb與5.3kb大小片段,與預計大小相符(圖2)。DNA測序結果顯示與人的syncytin ORF序列一致。可表達syncytin基因的真核表達載體構建成功。
2.3穩定表達Syncytin的EL4細胞系的鑒定
2.3.1pIRES2-syncytin-EGFP非融合綠色熒光蛋白的表達
在EL4-Syncytin細胞中,可清楚地觀察到熒光蛋白(圖3A、B、C、D),表明所構建的重組synctin載體已在EL4細胞表達。
2.3.2RT-PCR檢測syncytin,基因的表達
提取EL4-Syncytin細胞和空載體轉染細胞總RNA,以小鼠GAPDH基因為內參,運用qRT-PCR對syncytin進行相對定量。每個實驗組兩個復孔,實驗重復3次。通過各組樣品Ct值,利用2-△△Ct法計算得出EL4-Syncytin組為空載體對照組的120倍,表明syncytin在EL4細胞穩定高表達。實驗結果見圖4(A、B、C、D)。
2.3.3Western印記檢測人Syncytin表達
EL4-Syncytin細胞和空載體質粒穩定轉染細胞所提總蛋白經Western印記檢測,內參顯影條帶亮度一致,EL4-Syncytin組條帶亮度明顯高于對照組,通過灰度值計算軟件處理計算出實驗組與對照組數據(見圖5、6),EL4-Syncytin組蛋白表達水平明顯高于空載體組蛋白表達水平,表明Syncytin在EL4細胞穩定表達且高表達。
3討論
腫瘤對人類的生命和健康造成了極大的威脅。以白血病為例,白血病是國內10個高發的惡性腫瘤之一,是35歲以下人群發病率的死亡率最高的惡性腫瘤。白血病的病因與發病機制非常復雜,目前還未完全清楚,病毒因素、遺傳因素、染色體及免疫功能遺傳都與白血病發病有關。由于對大多數白血病發病的病因、病理機制缺乏系統深入研究,致使臨床上多數患者得不到早期診斷和規范治療,療效和預后較差。盡管目前已發現多個基因與腫瘤發生有關,但是能真正用于白血病診斷及治療的基因為數不多。尋找與白血病相關的基因,研究它們在白血病發生和發展中的作用機制,對白血病的早期診斷和治療具有重要意義。
人類內源性病毒(HERV)是遠古逆轉錄病毒感染人類基因組后留下的遺跡。其組成約占人類基因組3%~8%。目前,可分為至少31個家族[1,2]。由于基因突變的原因,大多數人類內源性病毒基因已失去轉錄活性[3]。只要少數的基因保留了完整的開放讀碼框。多項研究表明,人類內源性病毒基因的異常表達與腫瘤形成密切相關。Syncytin是HERVs-W家族的囊膜蛋白,主要在胎盤中表達,介導合胞滋養層的形成,維持胎盤正常的生理功能[4]。本研究小組在最近的研究中,偶然發現syn-cytin, mRNA和Syncytin蛋白在9個白血病或淋巴瘤細胞系中均有不同程度的表達,而且syncytin基因的表達水平與白血病患者病情相關。通過對Syncytin的結構分析發現,Syncytin內存在一個具有免疫抑制活性的肽段,其373~397殘基是一個具有免疫抑制活性的多肽,其在細胞表面的大量表達利于癌變細胞逃避免疫打擊,同時其具有的融合活性有利于細胞的遷移。目前未見到syncytin(mRNA及蛋白)與血液系統腫瘤的文獻報道,此基因與白血病的相關性為我們首次發現并已獲得專利。syncytin是怎樣參與白血病腫瘤細胞免疫逃逸機制并發揮作用目前仍待進一步研究。
本研究以C8166細胞總RNA為模板,通過RT-PCR擴增獲得syncytin的cDNA全長,并成功將其克隆到pIRES2-EGFP載體CMV啟動子下游,從而成功構建了可在真核細胞中表達Syn-cytin蛋白的pIRES2-syncytin-EGFP表達載體。將該載體轉染至EL4細胞,并通過篩選建立了人syncytin基因穩定表達的EL4細胞系。以空載體質粒穩定轉染EL4細胞為對照,用RT-PCR和Western印記分別從mRNA和蛋白水平檢測了Syncytin的表達。結果顯示,syncytin基因能有效地在EL4細胞轉錄和翻譯,表明成功建立了穩定表達人Syncytin的EL4細胞系。該真核表達載體的構建和穩定表達細胞系的建立,為syncytin在白血病腫瘤中的作用和其免疫逃逸機制的研究提供了必要的細胞模型,為后續功能實驗奠定了基礎。
參考文獻( References):
[1]BELSHAW R.KATZOURAKIS A,PACES J, et al.High copynumher in human endogenous retrovirus families is associatedwith copying mechanisms in addition to reinfection[J]. Molec-ular Biology and Evolution, 2005, 22(4):814-817.
[2]LOWER S,LOWER J,KURTH R.The viruses in all of us:characteristics and biological significance of human endogenous retrovirus sequences[J]. Proceedings of the National Acade-my of Sciences, 1996, 93(11):5177-5184.
[3]BANNERT N.KURTH R Retroelements and the humangenome: new perspectives on an old relation[J]. Proceedings ofthe National Academy of Sciences of the United States ofAmerica, 2004, 101:14572-14579.
[4]HARRIS J R.Placental endogenous retrovirus(ERV): structural,functional, and evolutionary significance[J]. BioEssays: News andReviews in Molecular, Cellular and Developmental Biology,1998, 20(4):307-316.
[5]LARSSON L.HOLCK S,CHRISTENSEN J Prognostic role ofsyncytin expression in breast cancer[J]. Human Pathology, 2007,38(5):726-731.
[6]BRODSKY I.FOLEY B,HAINES D,et al.Expression ofHERV-K provirus in human leukocytes[J]. Blood, 1993, 81(9):2369-2374.
[7]DEPIL S,ROCHE C,DUSSART P.et al.Expression of a hu-man endogenous retrovirus, HERV-K, in the blood cells ofleukemia patients[J]. Leukemia, 2002,16(2):254-259.
[8]SUN Y, OUYANG D Y.PANG W,et al.Expression of syncytin inleukemia and lymphoma cells[J]. Leukemia Research, 2010, 34(9):1195-1202.
[9]Ml S,LEE X,LI X,et al.Syncytin is a captive retroviral en-velope protein involved in human placental morphogenesis[J].Nature, 2000, 403(6771):785-789.
[10]BLOND J L,LAVILLETTE D,CHEYNETNV,et al.An enve-lope glycoprotein of human endogenous retrovirus HERV-W isexpressed in the human placenta and fuses cells expressingthe type D mammalian retrovirus receptor[J]. Journal of Virol-ogy, 2000, 74(7):3321-3329.
