細胞因子在萊姆病中作用的研究進展
2014-05-19 09:04:03王增田
中國藥理學通報 2014年3期
關鍵詞:研究
李 覃,邢 杰,王增田
(武警后勤學院1.病原生物與免疫學教研室、2.臨床醫學系,天津 300309)
萊姆病(Lyme disease)是一種由伯氏疏螺旋體(Borrelia burgdorferi,Bb)引起、經蜱傳播的人畜共患疾病,可感染馬、牛、羊等多種動物,能夠在人畜之間傳播流行,是嚴重危害人類健康的全球公共衛生問題,已于1992年被世界衛生組織(WHO)列入重點防治研究對象。近年來,對萊姆病流行病學、病原生物學、致病機制、診斷及綜合防治等方面的研究雖然已取得不少進展,但由于新的疫源地被不斷發現,Bb抗原結構的復雜性、疾病臨床癥狀的多樣性等原因,使得人們對該病的快速診斷和有效治療仍存在諸多挑戰。闡明其致病機制不但有助于建立高效、快速的診斷手段,而且有望找到新的藥物作用靶點,提高對疾病的防治能力。本文將就細胞因子在萊姆病中的作用作一綜述。
1 概述
萊姆病是美國和歐洲最常見的蜱媒傳染病,因1977年耶魯大學Steere博士在美國萊姆鎮(Lyme Town)首次發現而得名。據美國疾病控制中心(Centers for Disease Control and Prevention,CDC)統計[1],2009年全美有 29 959例萊姆病確診病例和8 509例疑似病患,位列十大傳染性疾病的第7名。目前亞、歐、美、非、大洋洲等世界五大洲70多個國家均報道有萊姆病存在,全世界每年感染人數在30萬左右,且發病區域和發病率呈迅速擴大和上升的趨勢[2]。萊姆病在我國同樣分布廣泛[3],自從艾承緒于1986年在黑龍江省海林縣林區發現萊姆病以來,全國已有29個省、市、自治區、直轄市人群中發現存在Bb感染的風險,受威脅人群不少于5億人。萊姆病臨床表現復雜多樣,在不同感染階段、不同基因型螺旋體均可引起不同癥狀,早期(<1月)多出現皮膚、神經系統、心血管系統病變,晚期(數月~數年)主要表現為關節炎、慢性萎縮性肢皮炎、淋巴細胞瘤、腦脊髓炎等,甚至可終生致殘、死亡,嚴重威脅人類健康和生活質量[4]。
2 炎癥因子在萊姆病發病過程中的作用
近年研究表明,萊姆病的病理進程與Bb感染和宿主免疫系統之間復雜的交互作用密切相關。Bb含有100多種不同的蛋白質,特別是外膜蛋白(outer surface protein,Osp)可誘導機體產生細胞和體液免疫應答,其中OspA和OspB抗原可能在疾病的發生發展中發揮關鍵作用,它們不僅通過Toll樣受體(Toll-like receptors,TLR)誘導多種免疫細胞分泌細胞因子,作為絲裂原刺激T淋巴細胞增殖和細胞因子分泌[5],還能通過分子模擬 hLFA-1與HLA-DR4/-DR2交叉反應,引起自身免疫應答,發展至抗生素抵抗性的慢性炎癥,損害肌骨骼系統、神經系統等[4,6]。目前,以 IL-6、IL-8、CCL2等炎癥因子為代表的細胞因子參與萊姆病的發病機制,并在其病程進展中發揮重要作用已獲得共識(Fig 1)。基因組學分析發現,Bb感染可導致CXC及CC類趨化因子、前炎癥因子以及TNF超家族的基因表達水平明顯增加,提示炎癥性細胞因子可能通過多種機制參與萊姆病的發病過程[7-8]。深入研究顯示,Bb進入機體后可通過與單核/巨噬細胞表面TLR結合,或誘導NK細胞、NKT細胞等固有免疫細胞產生IFN-γ、CCL2、CXCL9、CXCL10等致炎性細胞因子,引起組織細胞的損害[9];此外,這些細胞因子還可間接誘導T淋巴細胞在炎癥部位聚集,啟動機體適應性免疫應答。例如,萊姆性關節炎患者的免疫系統識別Bb后經免疫應答能產生大量TNF-α、IL-1β等炎癥因子,進而誘導滑膜細胞產生膠原酶和前列腺素,形成正反饋,加重關節炎癥反應和組織損傷[10-11]。
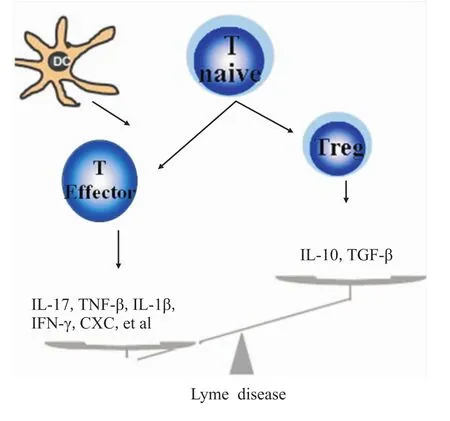
Fig 1 Action of cytokines in Lyme disease
2.1 趨化因子與萊姆病 趨化因子(chemokines,CXC)是一類能夠吸引白細胞移行到感染部位的細胞因子,具有白細胞趨化性,使單核/巨噬細胞等炎癥細胞遷移到炎癥部位,增強炎癥細胞的吞噬殺傷功能,促進炎癥介質釋放,在固有免疫和適應性免疫中均發揮重要作用。基因芯片檢測發現,3種不同的致病性Bb感染人表皮成纖維細胞后均可誘導出明顯上調的趨化因子類(CXCL1、IL-8)細胞因子基因表達,這些炎癥因子進一步誘導免疫應答,引起組織細胞損傷,其它如IL-6、TNF-α、NF-κB、MMPs(MMP-1、MMP-3、MMP-12)等分子水平亦明顯增加[12]。此外,研究人員還采用基于微球的多通道分析檢測了49名萊姆病患者的細胞因子含量,結果顯示抗生素耐藥的萊姆病患者CXCL9、CXCL10明顯增高[13]。Shin等[14]以Bb體外感染萊姆關節炎患者外周血CD14+單核/巨噬細胞,可刺激 CCL2產生,而分泌 CCL4、CXCL9和CXCL10等趨化因子還需要PBMC中其它細胞參與,提示Bb可能通過直接/間接刺激單核/巨噬細胞分泌趨化因子,從而參與萊姆病的固有免疫應答。對萊姆病小鼠的表達譜研究發現[15],Bb感染不同類型的關節炎動物模型后,多種趨化因子配體(chemokine ligand,CCL)均明顯上調(CCL2、CCL7、CCL8、CCL9、CCL10、CCL11、CCL12、CCL13、CCL14、CCL16)。此外,趨化因子CXCL1、CXCL9、CXCL10等在炎癥組織中大量表達,可引起T細胞增加,誘導單核細胞及其它免疫細胞向炎癥組織浸潤,加重萊姆病的發病過程。可見,趨化因子在萊姆病的致病機制中發揮著至關重要的作用,既可介導固有免疫應答,又能介導適應性免疫應答,其與萊姆病的關系日益引起國內外學者的關注。
2.2 IL-17與萊姆病 現已明確,Bb感染可明顯增加IL-6、IL-8、CCL2等產生,這些炎癥因子主要參與Th1型細胞免疫應答,促進細胞損傷,加重萊姆病的病理進程,在疾病起始階段發揮重要作用[7-8]。隨著研究不斷深入,一些新型細胞亞群及其效應因子的發現為細胞因子與萊姆病之間關系的研究提供了新的思路。例如,新型CD4+T淋巴細胞亞群Th17主要分泌產生IL-17,具有強大的募集和激活中性粒細胞的能力,同時調控組織炎性反應,已成為近年的研究熱點。IL-17家族至少有6個成員(IL-17A-F),目前,對該家族中IL-17A(以下稱為 IL-17)研究較多,主要通過與其受體(IL-17R)結合,并活化MAPK和NF-κB等信號傳導通路發揮生物學活性。除了直接參與炎癥反應之外,IL-17還能誘導促炎因子的產生(如 TNF-α、IL-6及 IL-1β),對炎癥反應局部化和擴大化起重要作用[16]。有研究人員為感染了Bb的小鼠注射IL-17抗體或IL-17R抗體后,臨床癥狀明顯減輕,并且得到組織病理學證實,說明IL-17在Bb感染后機體的適應性免疫應答過程中發揮重要作用[17-18]。此外,Bb進入機體后還能與宿主細胞相互作用促進IL-6釋放,進而誘導Th17細胞分泌 IL-17,引起下游細胞因子——IL-1β、TNF-α等大量釋放,放大炎癥反應,加重萊姆病的臨床表現。隨著對IL-17研究的不斷深入,人們發現,由樹突狀細胞、巨噬細胞等免疫細胞分泌的IL-23可刺激na?ve CD4+T細胞活化,在Th17細胞存活和維持中起重要作用,并能擴增和穩定Th17細胞表型,敲除IL-12p40和(或)IL-12p19即造成IL-23缺失,能降低IL-17表達、減少Th17細胞數量,使動物模型不易發生膠原誘導性關節炎等自身免疫性疾病[19]。Oosting等[20]研究證實,Bb的中性粒細胞激活蛋白A能夠誘導固有免疫應答產生IL-23,進而促進Th17細胞分化,使IL-17在萊姆病患者體內高水平表達。在進一步研究中,Oosting使caspase-1缺陷小鼠感染Bb,結果IL-17含量明顯升高,提示Bb誘導IL-17產生的作用機制可能與炎癥依賴的caspase-1途徑活化密切相關。盡管螺旋體表面膜蛋白被認為與萊姆病發病密切相關,但目前對能誘導Th17細胞活化及其效應因子IL-17釋放的致萊姆病的毒力因子仍不清楚,尚需進一步研究闡明。
3 抗炎因子與萊姆病
除了炎癥性細胞因子外,細胞因子還包括抗炎因子,炎癥因子與抗炎因子之間比例失衡導致的免疫調節紊亂,可引起或加重萊姆病的發生發展。其中一種重要的抗炎因子IL-10可由CD4+T淋巴細胞、單核巨噬細胞等多種細胞合成,既能直接抑制炎癥細胞黏附、浸潤,也能抑制單核巨噬細胞合成、釋放炎性細胞因子,具有較強的抗炎及免疫抑制活性。Giambartolomei等[21]以LPS刺激的單核細胞THP-1作為研究對象發現,Bb感染后細胞產生的IL-10可抑制炎癥因子釋放,而抑制IL-10產生則導致前炎癥因子IL-6、IL-12含量增加,提示IL-10可能在萊姆病的發病過程中發揮重要的負調節作用。最近,基因表達譜研究顯示,IL-10能抑制Bb感染的巨噬細胞中多種基因的功能性表達,包括TLR1、TLR2、MMP-9,從而控制炎癥反應[22];蛋白組學分析同樣證實,IL-10在 CXCL2、CXCL10、CCL3、CXCL1、CXCL5、CCL2、CCL4、GCSF、IL-6、TNF-α、IL-1β等炎癥因子的產生過程中可發揮重要的調節作用,其作用機制可能與抑制巨噬細胞激活或T細胞活化有關[23]。Sonderegger等[24]進一步研究證實,IL-10基因敲除小鼠往往伴隨著NK細胞、巨噬細胞等炎癥細胞在Bb感染部位的大量浸潤,更易出現萊姆性關節炎,這些數據表明,IL-10可通過負調節炎癥反應抑制萊姆病的發生發展。但是,也有研究人員指出[25],當Bb感染具有遺傳易感性的萊姆性關節炎小鼠后,IL-10并不能延緩關節炎的病理進程,對螺旋體清除率或疏螺旋體特異性抗體生成亦無影響。因此,IL-10在萊姆病中究竟發揮何種作用及其具體調節機制仍需進一步深入研究。
TGF-β是另一種多功能細胞因子,可影響細胞生長、分化、凋亡,在炎癥因子TNF-α的協同作用下可調節細胞轉化,對調控機體免疫應答具有重要作用。研究發現,TGF-β及下游信號分子的不恰當激活可導致炎癥激活,其作用機制可能是作為化學趨化劑募集炎癥細胞、促進前炎性細胞因子生成、使細胞疏松化、氣球樣變[26]。研究人員檢測了40例萊姆病患者血清,結果顯示患者體內TGF-β水平明顯低于正常對照組,并且與MMP-2含量具有一定的相關性,說明炎癥因子和抗炎因子的交互作用能夠參與萊姆病的發病過程[27]。
值得一提的是,近年來CD4+CD25+調節性T細胞(regulatory T cell,Treg)與萊姆病的關系引起越來越多的關注。Treg是新發現的重要免疫調節細胞,在維持炎癥免疫性疾病的自身平衡中發揮重要作用。雖然目前Tregs免疫抑制的機制尚未完全明確,現多認為可能有以下幾種形式[28]:①直接細胞接觸途徑。Tregs通過細胞表面的膜分子CTLA-4和膜TGF-β與效應細胞相應的配體結合,直接抑制炎癥因子分泌;②分泌抑制性細胞因子。Tregs可在某些趨化因子作用下遷移至免疫應答的局部,通過細胞接觸作用后分泌IL-10、TGF-β抗炎因子等發揮抑制作用;③間接抑制途徑。Tregs可通過下調抗原提呈細胞表面CD80/CD86、CD40、MHC-Ⅰ/MHC-Ⅱ等分子表達,進而抑制對效應細胞的抗原提呈與活化。最近,Vudattu等[29]在關節炎方面的權威期刊《Arthritis Rheum》報道,抗生素耐藥的萊姆性關節炎患者關節滑液中Treg亞群數量明顯降低,糖皮質激素誘導的腫瘤壞死因子受體(TNFR)表達明顯增加,因此不能有效抑制前炎性因子產生,導致患者免疫調節功能失衡。Nardelli等[30]發現,為感染了Bb的萊姆病小鼠注射IL-17抗體進行治療后,局部淋巴結Treg細胞數量明顯增加,臨床癥狀得到明顯緩解;而同時給予IL-17抗體和CD25抗體治療后,局部淋巴結Treg細胞數量下降,病變加重,說明Treg可有效預防和治療Bb引起的萊姆病發病,靶向Treg的免疫干預策略能夠針對萊姆病發病過程中的免疫應答,減輕炎癥反應,可能成為有效防治萊姆病的新靶點,并為開發新型抗萊姆病藥物提供研究方向。
4 結語與展望
蜱傳播引起的萊姆病是重要的人畜共患疾病,嚴重威脅人類健康和畜牧業生產,如何快速診斷和加強防治已成為迫切需要解決的問題。隨著高通量表達檢測技術的發展,科研人員不斷發現參與萊姆病發病過程的細胞因子,有的促進炎癥發生,有的抑制炎癥反應。因此,鑒定萊姆病中的異常的細胞因子可為人們提供研制新型防治藥物的作用靶標,還可為針對細胞因子的萊姆病治療提供新策略。但是,目前對細胞因子在萊姆病發病過程中作用的研究還處于起步階段,包括明確細胞因子的作用通路、提高利用細胞因子檢測萊姆病的靈敏度和特異性、確定萊姆病致病機制中相關的細胞因子靶蛋白等問題還有待進一步研究闡明。相信這一系列問題的解決,不僅有利于人們對萊姆病發生發展機制的了解,還將大大促進細胞因子在萊姆病預防、診斷和新藥研發中的應用。
參考文獻:
[1]Shen A K,Mead P S,Beard C B.The Lyme disease vaccine-a public health perspective[J].Clin Infect Dis,2011,52(Suppl 3):s247-52.
[2]Williams S,Matthews M,Barnett T.Lyme disease update:30 years after the first cases,the clinical picture remains cloudy[M].Adv NPs PAs,2011,2(7):27-8,30-2,38.
[3]牛慶麗,殷 宏,羅建勛.國內萊姆病研究進展 [J].動物醫學進展,2009,30(10):89-93.
[3]Niu Q L,Yin H,Luo J X.Progress on Lyme disease in China[J].Prog Veterinary Med,2009,30(10):89-93.
[4]Wright W F,Riedel D J,Talwani R,Gilliam B L.Diagnosis and management of Lyme disease[J].Am Fam Physician,2012,85(11):1086-93.
[5]Schempp C,Owsianowski M,Lange R,et al.Comparison of Borrelia burgdorferi ultrasonicate and whole B,burgdorferi cells as a stimulus for T-cell proliferation and GM-CSF secretion in vitro[J].Zentralbl Bakteriol,1993,279(3):417-25.
[6]Gross D M,Forsthuber T,Tary-Lehmann M,et al.Identification of LFA-1 as a candidate autoantigen in treatment-resistant Lyme arthritis[J].Science,1998,281(5377):703-6.
[7]Krupka M,Zachova K,Weigl E,et al.Prevention of lyme disease:promising research or sisyphean task[J]?Arch Immunol T-her Exp(Warsz),2011,59(4):261-75.
[8]Wang G,Petzke M M,Iyer R,et al.Pattern of pro-inflammatory cytokine induction in RAW264.7 mouse macrophages is identical for virulent and attenuated Borrelia burgdorferi[J].J Immunol,2008,180(12):8306-15.
[9]Kawai T,Akira S.The role of pattern-recognization receptors in innate immunity:Update on Toll-like receptors[J].Nat Immunol,2010,11(5):373-84.
[10]Strle K,Shin J J,Glickstein L J,et al.Association of a Toll-like receptor 1 polymorphism with heightened Th1 inflammatory responses and antibiotic-refractory Lyme arthritis[J].Arthritis Rheum,2012,64(5):1497-507.
[11]Nardelli D T,Callister SM,Schell R F.Lyme arthritis:Current concepts and a change in paradigm[J].Clin Vaccine Immunol,2008,15(8):21-34.
[12]Schramm F,Kern A,Barthel C,et al.Microarray analyses of inflammation response of human dermal fibroblasts to different strains of borrelia burgdorferi sensu stricto[J].PLoS One,2012,7(6):e40046.
[13]Strle K,Shin JJ,Glickstein L J,et al.Association of a Toll-like receptor 1 polymorphism with heightened Th1 inflammatory responses and antibiotic-refractory Lyme arthritis[J].Arthritis Rheum,2012,64(5):1497-507.
[14]Shin J J,Strle K,Glickstein L J,et al.Borrelia burgdorferi stimulation of chemokine secretion by cells of monocyte lineage in patients with Lyme arthritis[J].Arthritis Res Ther,2010,12(5):R168.
[15]Miller J C,Ma Y,Crandall H,et al.Gene expression profiling provides insights into the pathways involved in inflammatory arthritis development:murine model of Lyme disease[J].Exp Mol Pathol,2008,85(1):20-7.
[16]Qu N,Xu M,Mizoguchi I,et al.Pivotal roles of T-helper 17-related cytokines,IL-17,IL-22,and IL-23,in inflammatory diseases[J].Clin Dev Immunol,2013,2013(7):9685-49.
[17]Burchill MA,Nardelli DT,England DM,et al.Inhibition of interleukin-17 prevents the development of arthritis in vaccinated mice challenged with Borrelia burgdorferi[J].Infect Immun,2003,71(6):3437-42.
[18]Oosting M,ter Hofstede H,van de Veerdonk F L,et al.Role of interleukin-23(IL-23)receptor signaling for IL-17 responses in human Lyme disease[J].Infect Immun,2011,79(11):4681-7.
[19]楊金娜,劉曉光,李 覃,等.Th17/Treg平衡在類風濕關節炎中作用的研究進展 [J].中國藥理學通報,2013,29(8):1045-8.
[19]Yang JN,Liu X G,Li T,et al.Function of Th17/Treg balance in rheumatoid arthritis[J].Chin Pharmacol Bull,2013,29(8):1045-8.
[20]Oosting M,van de Veerdonk F L,Kanneganti TD,et al.Borrelia species induce inflammasome activation and IL-17 production through a caspase-1-dependent mechanism[J].Eur J Immunol,2011,41(1):172-81.
[21]Giambartolomei G H,Dennis V A,Lasater B L,et al.Autocrine and exocrine regulation of interleukin-10 production in THP-1 cells stimulated with Borrelia burgdorferi lipoproteins[J].Infect Immun,2002,70(4):1881-8.
[22]Gautam A,Dixit S,Philipp M T,et al.Interleukin-10 alters effector functions of multiple genes induced by Borrelia burgdorferi in macrophages to regulate Lyme disease inflammation[J].Infect Immun,2011,79(12):4876-92.
[23]Youn Y,Sung I K,Lee I G.The role of cytokines in seizures:interleukin(IL)-1β,IL-1Ra,IL-8,and IL-10[J].Korean J Pediatr,2013,56(7):271-4.
[24]Sonderegger F L,Ma Y,Maylor-Hagan H,et al.Localized production of IL-10 suppresses early inflammatory cell infiltration and subsequent development of IFN-γ-mediated Lyme arthritis[J].J Immunol,2012,188(3):1381-93.
[25]Brown CR,Lai A Y,Callen ST,et al.Adenoviral delivery of interleukin-10 fails to attenuate experimental Lyme disease[J].Infect Immun,2008,76(12):5500-7.
[26]Worthington J J,Fenton T M,Czajkowska B I,et al.Regulation of TGF-βin the immune system:an emerging role for integrins and dendritic cells[J].Immunobiol,2012,217(12):1259-65.
[27] Pancewicz S A,Izycka A,Klibingat M,et al.Transforming growth factor beta,metalloproteinase2 and its tissue inhibitor 2 in the serum from patients with early and late boreliosis[J].Pol Merkur Lekarski,2008,25(150):495-9.
[28]Shevach E M.Immunology.Regulating suppression[J].Science,2008,322(5899):202-3.
[29]Vudattu N K,Strle K,Steere A C,et al.Dysregulation of CD4+CD25(high)T cells in the synovial fluid of patients with antibiotic-refractory Lyme arthritis[J].Arthritis Rheum,2013,65(6):1643-53.
[30]Nardelli D T,Burchill M A,England D M,et al.Association of CD4+CD25+T cells with prevention of severe destructive arthritis in Borrelia burgdorferi-vaccinated and challenged gamma interferon-deficient mice treated with anti-interleukin-17 antibody[J].Clin Diagn Lab Immunol,2004,11(6):1075-84.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19