擬目烏賊卵子發(fā)生與卵巢發(fā)育
2014-11-05 06:33:00蔣霞敏柳敏海彭瑞冰
水生生物學報 2014年6期
羅 江 蔣霞敏 柳敏海 唐 鋒 彭瑞冰
(1. 寧波大學海洋學院, 寧波 315211; 2. 浙江省舟山市水產研究所, 舟山 316000)
卵子發(fā)生是個體發(fā)育的重要環(huán)節(jié)之一, 卵子的質量好壞直接影響到受精過程以及受精卵的發(fā)育,而卵巢發(fā)育程度的好壞直接決定卵子的質量[1]。關于頭足類卵子發(fā)生和卵巢發(fā)育的研究, 國外較多,主要集中在皮氏槍烏賊(Loligo pealei)[2]、巴塔哥尼亞槍烏賊(Loligo gahi)[3]、福氏槍烏賊(Loligo forbesi)[4]、狼烏賊(Lycoteuthis lorigera)[5]、尖盤艾爾斗蛸(Eledone cirrhosa)[6]和真蛸(Loligo vulgaris)[7]等,國內鮮有報道, 僅見曼氏無針烏賊(Sepiella maindroni)[8]、金烏賊(Sepia esculenta)[9]和長蛸(Octopus variabilis)[10], 迄今為止, 未見擬目烏賊(Sepia lycidas)相關報道。
擬目烏賊隸屬于軟體動物門(Mollusca)、頭足綱(Cephalopoda)、鞘亞綱(Coleoidea)、烏賊目(Sepiida)、烏賊科(Sepiidae)、烏賊屬(Sepia), 為淺海暖水性較強的底棲種, 20世紀80年代前我國東海有分布, 目前主要分布在南海[11]。因其個體大, 生長快, 具有較高的營養(yǎng)價值和人工養(yǎng)殖前景, 已引起國內外學者的關注[12—23]。2013年5月, 本課題組率先突破了擬目烏賊規(guī)模化人工育苗技術, 為其人工養(yǎng)殖的研究奠定了基礎。本文采用解剖學和組織學的方法,對水泥池養(yǎng)殖條件下擬目烏賊卵子發(fā)生和卵巢發(fā)育進行了研究, 闡明了擬目烏賊卵子發(fā)生和卵巢發(fā)育規(guī)律, 可豐富頭足類發(fā)育生物學理論知識, 對擬目烏賊人工育苗和養(yǎng)殖具有十分重要的指導意義。
1 材料與方法
1.1 材料
試驗于2013年2—11月進行, 擬目烏賊親體捕自廣東省湛江市硇洲島海域(21.11°N, 110.24°E), 經自然交配產卵, 在浙江省舟山市水產研究所朱家尖基地進行孵化, 孵化水溫為(19—21)℃, 鹽度為24—26, 烏賊幼體孵出后在4.0 m×5.0 m×1.2 m的水泥池內進行培育, 培育水溫為(20—32)℃, 鹽度22—30, 連續(xù)充氣, 充氣頭1個/m2。日投餌2次, 投餌種類和投喂量隨烏賊個體的增長而改變: 0—45日齡(胴長 0.72—3.83 cm)投喂活糠蝦(0.5—1.0 cm), 投喂量(10—20) ind./L; 45—90日齡(胴長3.83—6.52 cm)投喂冰鮮小雜魚, 切成長條狀(1.5 cm×1.0 cm), 日投喂量為體重的18%; 90日齡以后(胴長6.52 cm以上)投喂冰鮮小雜魚, 切成長條狀(3.0 cm×1.0 cm),日投喂量為體重的12%。每天換水1—2次, 換水量80%—100%。
1.2 方法
取樣 從剛孵化出膜的小烏賊開始, 1d取樣一次, 10d后每隔3天取樣一次, 45d后每隔15d取樣一次, 每次隨機取樣10只, 對于肉眼不能分辨性腺的個體, 用波恩氏液整體固定4h后, 切取胴部后段,換新的波恩氏液保存于 4℃冰箱中; 對于肉眼可分辨性腺的個體, 直接取性腺固定, 24h后換新的波恩氏液保存于4℃冰箱中。
組織學切片 酒精梯度脫水, 二甲苯透明,石蠟包埋, KD-2508型輪轉式切片機連續(xù)切片, 切片厚度(5—8) μm, HE染色, 中性樹膠封片, OlympusBX-50型熒光顯微鏡觀察并拍照。
數(shù)據(jù)測量與處理 記錄烏賊日齡(Age)、體重(W)、胴長(ML)和卵巢重(Wo), 細胞直徑和細胞核直徑直接在顯微照片上測量, 按以下公式分別計算核質比(NP)和性腺指數(shù)(GSI):
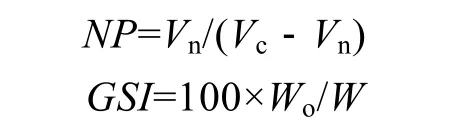
其中, Vn為細胞核的體積, Vc為細胞體積, 由公式V=4/3πR3求得。
2 結果
2.1 卵子發(fā)生分期
擬目烏賊卵子發(fā)生不同步, 根據(jù)細胞形態(tài)、細胞大小、濾泡細胞形態(tài)和卵黃形成情況可以分為卵原細胞階段(卵原細胞期)、原生質生長階段(無濾泡期、單層濾泡期和雙層濾泡期)、間質生長階段(濾泡內折早期、濾泡內折中期和濾泡內折晚期)和營養(yǎng)質生長階段(卵黃發(fā)生早期、卵黃發(fā)生晚期和成熟期), 共4個階段, 10個時期。
卵原細胞階段 第1期 前期卵原細胞期。前期卵原細胞形狀不規(guī)則或略呈圓形, 直徑為(12—15) μm, 細胞核呈圓形, 直徑為(10—12) μm, 細胞核占據(jù)整個細胞的絕大部分(圖版Ⅰ-1), NP為1.049—1.373。
原生質生長階段 一部分卵原細胞經過有絲分裂繼續(xù)保持增殖能力, 另一部分卵原細胞經過減數(shù)分裂發(fā)育為卵母細胞。此時, 細胞直徑和細胞核直徑均不斷增加, 細胞質繼續(xù)增多, 根據(jù)濾泡細胞的形態(tài)和分布的變化將此階段分為無濾泡期、單層濾泡期和雙層濾泡期3個時期。
第 2期 無濾泡期。細胞呈圓形或橢圓形, 直徑為(40—170) μm, 細胞核呈橢圓形, 直徑為(25—90) μm (圖版Ⅰ-2), 細胞外圍沒有濾泡細胞,NP為0.174—0.492。
第3期 單層濾泡期。細胞呈圓形或橢圓形, 直徑為(220—310) μm, 細胞核呈橢圓形, 直徑為(100—120) μm (圖版Ⅰ-3), NP 為 0.062—0.103。隨著卵母細胞體積的增大, 扁平的濾泡細胞(長9—12 μm)開始從卵母細胞的一側, 逐漸附于細胞膜上, 包圍整個卵母細胞, 形成單層濾泡細胞層(圖版Ⅱ-1)。
第4期 雙層濾泡期。細胞呈圓形或橢圓形, 直徑為(290—380) μm, 細胞核呈橢圓形, 直徑為(110—130) μm (圖版Ⅰ-4), NP 為 0.042—0.073。卵母細胞體積繼續(xù)增大, 濾泡細胞繼續(xù)附于細胞膜上,形成雙層濾泡細胞層, 內層濾泡細胞變?yōu)闄E圓形,直徑為(7—10) μm, 外層濾泡細胞呈扁平狀, 長為(10—15) μm (圖版Ⅱ-2)。
間質生長階段(濾泡內折階段) 間質生長階段也可以稱為濾泡內折階段, 主要特征是濾泡細胞開始向卵母細胞內部折疊, 并逐漸占據(jù)整個卵母細胞。根據(jù)濾泡細胞的分布情況, 可以將此階段分為早期、中期和晚期3個時期。
第5期 濾泡內折早期。細胞呈圓形或橢圓形,直徑為(340—400) μm, 細胞核呈圓形或橢圓形, 直徑為(150—170) μm (圖版Ⅰ-5), NP 為 0.083—0.094。位于內層的橢圓形濾泡細胞先向卵母細胞內部延伸,形成 8—10個指狀突起, 這些指狀突起的長度大體一致, 長為(30—40) μm (圖版Ⅰ-5、圖版Ⅱ-3)。
第6期 濾泡內折中期。細胞呈圓形或橢圓形,直徑為(390—470) μm, 細胞核呈圓形或橢圓形, 直徑為(160—180) μm (圖版Ⅰ-6), NP 為 0.060—0.074。濾泡細胞繼續(xù)向卵母細胞內部延伸, 占據(jù)了整個細胞的一半左右, 指狀突起的數(shù)量增加到 10—12個,長(80—170) μm 不等, 且有些指狀突起出現(xiàn)分支(圖版Ⅰ-6), 位于外層扁平的濾泡細胞直徑縮短至(8—10) μm, 隨著指狀突起的深入, 也延伸至卵母細胞內部(圖版Ⅱ-4)。
第 7期 濾泡內折晚期。細胞呈橢圓形, 體積急劇增大, 直徑為(1000—1200) μm, 細胞核呈橢圓形, 直徑為(150—180) μm, 偏向細胞一側(圖版Ⅰ-7), NP為 0.003—0.005。指狀突起不斷延伸和分支, 相互連接呈網狀, 占據(jù)整個卵母細胞, 扁平的濾泡細胞消失(圖版Ⅱ-5)。
營養(yǎng)質生長階段 濾泡細胞占據(jù)整個卵母細胞后逐漸解體, 細胞核也消失不見, 與此同時, 卵黃物質開始積累, 根據(jù)濾泡細胞的分布和卵黃的積累情況, 將此階段分為卵黃發(fā)生早期、卵黃發(fā)生晚期和成熟期3個時期。
第 8期 卵黃發(fā)生早期。細胞呈橢圓形, 直徑為(1500—1800) μm, 網狀的濾泡細胞從卵母細胞的中間開始解體(圖版Ⅰ-8), 解體的濾泡細胞邊界不明顯, 直徑為(7—10) μm, 相互堆積(圖版Ⅱ-6), 細小的顆粒狀卵黃物質相互聚集, 呈小塊狀, 被HE染成淡藍色(圖版Ⅰ-8)。
第9期 卵黃發(fā)生晚期。細胞體積急劇增大, 呈橢圓形, 直徑為(2300—2900) μm, 大部分濾泡細胞已經解體, 整個卵母細胞充滿了卵黃物質, 呈塊狀,被染HE成橘紅色(圖版I-9)。
第10期 成熟期。成熟卵子呈黃色透明的橢圓形, 直徑為(6000—7000) μm, 內充滿了大量卵黃,切片時容易破碎, 因而未獲得組織學結構。
2.2 卵巢發(fā)育分期
根據(jù)卵巢外觀形態(tài)、性腺指數(shù)變化和切面上各期細胞所占的比例, 將擬目烏賊卵巢發(fā)育分為形成前期、形成期、小生長期、大生長期、成熟前期和成熟期6個時期。
Ⅰ期 形成前期 烏賊日齡 23—31d, 體重(3.03—4.82) g, 胴長(2.43—3.22) cm, 性腺肉眼不可見, 組織切片觀察到卵巢位于胴體基部, 切面上生殖細胞均為卵子發(fā)生第1期的細胞, 少量被HE染成深藍色的扁平濾泡細胞散布于卵巢中(圖版Ⅲ-1、表 1)。
Ⅱ期 形成期 烏賊日齡 31—45d, 體重(4.82—8.56) g, 胴長(3.22—3.83) cm, 性腺位于體腔后部, 包被于體腔膜中, 肉眼不容易觀察到, 呈乳白色, 細條狀, 長約 4 mm, 重(0.01—0.02) g, 性腺指數(shù)為 0.16—0.28, 組織切片觀察, 卵巢內生殖細胞以卵子發(fā)生第 2期細胞為主, 占整個切面的83.66%, 還分布著少量第 3期(13.96%)和第 1期(2.38%)的細胞(圖版Ⅲ-2、表 1)。
Ⅲ期 小生長期 烏賊日齡 45—150d, 體重(8.56—133.81) g, 胴長(3.83—11.20) cm, 肉眼明顯可見卵巢呈乳白色, 扁平狀, 緊貼體腔膜, 大小(6—8) mm, 重(0.02—0.23) g, 性腺指數(shù)為 0.12—0.26, 組織切片觀察, 卵巢內生殖細胞以卵子發(fā)生第4期細胞為主, 占整個切面的50.01%, 其次為第3期細胞(36.33%), 仍然可見少量第2期(11.34%)和第1期(1.09%)細胞, 且切面上出現(xiàn)了部分退化吸收的卵母細胞, 占整個切面的1.23% (圖版Ⅲ-3、表1)。
Ⅳ期 大生長期 烏賊日齡 150—180d, 體重(133.81—256.34) g, 胴長(11.20—13.55) cm, 卵巢呈乳白色, 扁平狀, 肉眼可觀察到卵巢表面具有凹凸不平的卵粒, 此期卵巢開始快速增長, 大小(12—18) mm, 重 (0.23—1.56) g, 性 腺指 數(shù) 為0.17—0.72, 組織切片觀察, 卵巢內生殖細胞以卵子發(fā)生第6期細胞為主, 占整個切面的50.27%, 其次為第5期細胞(18.83%), 第1期細胞不再出現(xiàn)(圖版Ⅲ-4、表1)。
Ⅴ期 成熟前期 烏賊日齡 180d后, 體重達256.34 g以上, 胴長達13.55 cm以上, 卵巢繼續(xù)增長, 乳白色, 葡萄狀, 大小(20—25) mm, 重 1.56 g以上, 性腺指數(shù) 0.65以上, 肉眼可見橢球形的卵母細胞呈串排列。組織切片觀察, 卵巢內生殖細胞以卵子發(fā)生第 8期細胞為主, 占整個切面的 61.85%,退化吸收的卵母細胞增多, 占整個切面的2.05% (圖版Ⅲ-5、表1)。
Ⅵ期 成熟期 成熟卵巢從親體中獲得, 此時烏賊已經性成熟, 體重(950.33—2010.28) g, 胴長(20.80—27.75) cm, 卵巢呈淡黃色, 葡萄狀, 充滿整個胴體后部, 重(31.35—75.23) g, 性腺指數(shù)為2.82—4.05, 淡黃色的成熟卵子游離到生殖腔中, 偶見的未成熟卵子呈乳白色(圖版Ⅲ-6、表1)。
3 討論
3.1 關于性成熟和繁殖周期
頭足類繁殖周期不同種類之間存在較大差異,韋柳枝等認為金烏賊為一年性成熟, 繁殖周期為一年[24]; Goff和 Daguzan研究發(fā)現(xiàn)烏賊(Sepia officinalis)有兩種繁殖群體, 第一種群體的數(shù)量和個體均較小, 繁殖周期為一年, 第二種群體的數(shù)量和個體均較大, 繁殖周期為 2年[25]; 曼氏無針烏賊在自然條件下一年只繁殖一次, 而在人工養(yǎng)殖條件下, 春季產出的幼體在秋季即可達到性成熟, 一年有兩次繁殖期[26]。本實驗表明, 擬目烏賊孵化出膜后, 經過 4—6個月的水泥池養(yǎng)殖, 平均體重為 256.34 g,最大體重可達到 457.08 g, 大部分個體的卵巢發(fā)育至第Ⅳ期, 個別已發(fā)育至第Ⅵ期, 達到性成熟, 因此我們推測人工養(yǎng)殖條件下擬目烏賊一年可達到性成熟, 繁殖周期為一年。一般來說頭足類達到性成熟后, 即交配、產卵, 隨后大量死亡, 特別是曼氏無針烏賊性早熟現(xiàn)象普遍, 養(yǎng)殖3—4個月, 長至50 g左右就性早熟, 交配產卵, 伴隨大量死亡, 這嚴重挫傷了養(yǎng)殖戶的積極性, 阻礙了烏賊養(yǎng)殖業(yè)的發(fā)展,而擬目烏賊生長快, 養(yǎng)殖 4—6個月就可長至250—500 g, 達到商品規(guī)格, 而且只出現(xiàn)個別性成熟, 這為養(yǎng)殖戶帶來了福音。
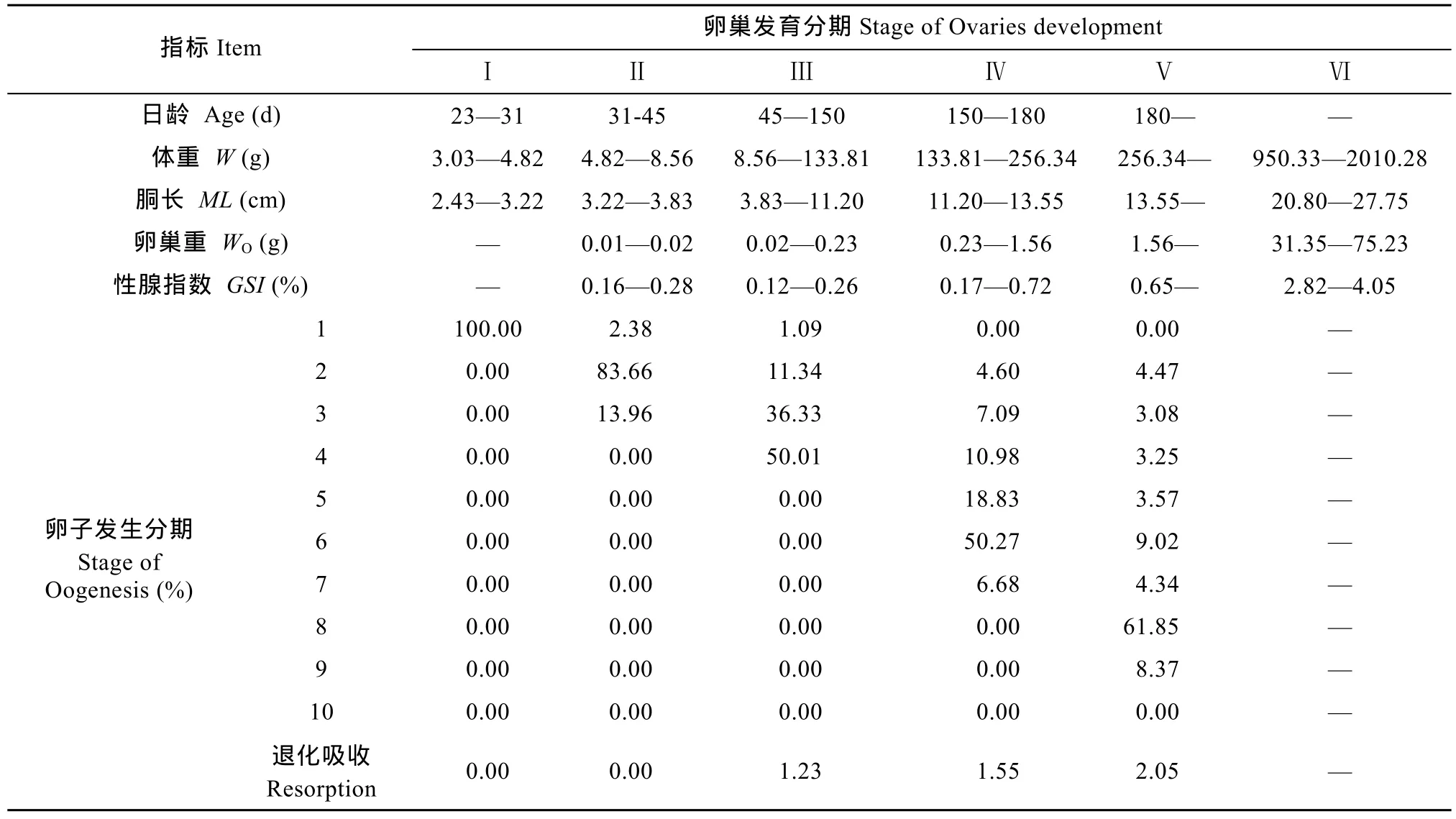
表1 擬目烏賊卵巢發(fā)育不同時期切面上各期細胞所占的比例Tab. 1 Percentage of each oocyte at the sections during ovaries development in Sepia lycidas
3.2 關于卵子發(fā)生和卵巢發(fā)育的分期
關于頭足類卵子發(fā)生的分期, 不同學者的分類依據(jù)不盡相同。Bottke[27]根據(jù)濾泡細胞形態(tài)變化, 將錐異尾槍烏賊(Alloteuthis subulata)的卵子發(fā)生分為了Ⅰ、Ⅱ和Ⅲ 3個階段, 而Selman和Arnold[2]根據(jù)濾泡細胞形態(tài)變化將皮氏槍烏賊卵子發(fā)生分為了Ⅰ—Ⅴ 5個階段, Boyle和Chevis[6]根據(jù)卵黃發(fā)生情況將尖盤艾爾斗蛸卵子發(fā)生也分為了Ⅰ、Ⅱ和Ⅲ3個階段, 蔣霞敏等[8]和李來國[10]根據(jù)細胞大小、細胞核形態(tài)和濾泡細胞形態(tài)變化分別將曼氏無針烏賊和長蛸卵子發(fā)生分為卵原細胞期、卵母細胞期、成熟期和退化期4個階段。本文綜合根據(jù)細胞形態(tài)、細胞大小、濾泡細胞形態(tài)和卵黃發(fā)生情況, 將擬目烏賊卵子發(fā)生分為卵原細胞、原生質生長、間質生長和營養(yǎng)質生長 4個階段, 這與 Laptikhovsky和Arkhipkin[3]對巴塔哥尼亞槍烏賊及Hoving和 Laptikhovsky[5]對狼烏賊卵子發(fā)生的分期類似。
關于頭足類卵巢發(fā)育的分期, 尚未有統(tǒng)一的標準。Perez和Haimovici[28]根據(jù)卵巢的厚度、顏色和卵細胞直徑將大愛爾斗蛸(Eledone Massyae)卵巢發(fā)育分為了Ⅰ—Ⅳ4個時期, Estefanell等[7]根據(jù)性腺指數(shù)變化將真蛸卵巢發(fā)育也分為了Ⅰ—Ⅳ4個時期,Boyle和 Chevis[6]根據(jù)卵黃發(fā)生情況, 將尖盤艾爾斗蛸卵巢發(fā)育分為了卵黃發(fā)生前期、卵黃發(fā)生早期、卵黃發(fā)生晚期和成熟期4個時期, Gabr等[29]根據(jù)卵巢的組織學結構將虎斑烏賊(Sepia pharaonis)和多氏烏賊(Sepia dollfusi)卵巢發(fā)育分為未成熟期、成熟期、產卵前期和產卵期 4個時期, 本文根據(jù)卵巢的外觀形態(tài)、性腺指數(shù)變化和切面上各期細胞所占的比例, 將擬目烏賊卵巢發(fā)育分為形成前期、形成期、小生長期、大生長期、成熟前期和成熟期6個時期,這種分期方式綜合了形態(tài)學、組織學和數(shù)據(jù)三方面依據(jù), 比較全面, 可作為日后研究頭足類卵巢發(fā)育分期的一個參考標準。
3.3 關于濾泡細胞的變化與卵黃的形成
濾泡細胞伴隨著卵母細胞發(fā)育的整個過程, 其形態(tài)和分布的變化是卵子發(fā)生分期的重要依據(jù)。關于濾泡細胞的起源, Nakamura[30]、張耀光等[31]和牟振波等[32]均認為濾泡細胞起源于非生殖細胞, 由卵巢基質分化而來, 而Broow[33]和Bolognari等[34]認為濾泡細胞起源于生殖細胞, 由早期卵原細胞分化而來, 在本研究中, 擬目烏賊濾泡細胞和卵原細胞同時出現(xiàn)于卵巢發(fā)育的第Ⅰ期, 濾泡細胞形態(tài)和大小均不同于卵原細胞, 因而我們認為擬目烏賊濾泡細胞起源于非生殖細胞, 由卵巢基質分化而來。在擬目烏賊卵子發(fā)生過程中, 濾泡細胞經歷了一個由包圍卵母細胞的扁平單層, 轉變?yōu)橥鈱颖馄絻葘訖E圓形的雙層, 內折形成指狀突起, 直至占據(jù)整個卵母細胞的網狀結構, 最后解體消失的過程, 這與曼氏無針烏賊[8]和長蛸[10]卵子發(fā)生過程中濾泡細胞的變化相類似, 在巴塔哥尼亞槍烏賊[3]卵子發(fā)生過程中濾泡細胞經歷了魚鱗狀和圓柱形, 這與本文描述的濾泡細胞經歷了扁平狀和橢圓形是相似的, Selman和 Arnold[2]研究發(fā)現(xiàn)皮氏槍烏賊濾泡細胞在卵子發(fā)生過程中是逐漸拉長的, Laptikhovsky和Arkhipkin[3]、Hoving 和 Laptikhovsky[5]也得到了類似的結果, 但本研究發(fā)現(xiàn), 擬目烏賊外層扁平濾泡細胞先拉長后縮短, 而內層橢圓形的濾泡細胞直徑變化不大。
關于濾泡細胞的功能, 眾說紛紜, 龔啟祥和杜立勤[35]、張耀光和謝小軍[31]認為濾泡細胞參與卵膜的形成, Grier[36]認為濾泡細胞具有合成和運輸外源性卵黃前體物質的作用, 而Selman和Arnold[2]認為濾泡細胞同時具有合成和分泌卵膜物質和卵黃前體物質的作用, Thiaw和Mattei[37]認為濾泡細胞對退化吸收的卵母細胞還具有二次包裹和吞噬的功能。在本文中, 卵黃的形成與濾泡細胞解體同時發(fā)生, 推測濾泡細胞解體前合成了大量卵黃前體物質, 在解體時分泌到卵母細胞中, 從而在短時間內合成了大量卵黃顆粒, 因而我們認為濾泡細胞參與了卵黃的合成,這與Bottke[27]認為卵黃由濾泡細胞產生的觀點一致。
3.4 關于細胞大小的變化
卵細胞的體積在卵子發(fā)生過程中發(fā)生了較大的變化, 不同物種之間這種變化也不同, 表 2比較了不同物種卵子發(fā)生過程中卵細胞直徑的變化, 可看出各個時期卵細胞直徑在不同物種之間具有較大差異, 且在卵子發(fā)生過程中, 卵細胞的直徑會出現(xiàn)跳躍性的增長, 曼氏無針烏賊[8]卵細胞跳躍性增長出現(xiàn)在第6—7期, 此期是濾泡細胞大量內折占據(jù)整個卵母細胞的時期, 巴塔哥尼亞槍烏賊[3]和狼烏賊[5]卵細胞跳躍性增長出現(xiàn)在第10期, 此期是卵黃大量合成的時期, 擬目烏賊卵細胞出現(xiàn)了兩次跳躍性增長, 第一次出現(xiàn)在第 7期, 即濾泡細胞大量內折期,第二次出現(xiàn)在第 9—10期, 即卵黃大量合成期, 可見濾泡細胞的增長和卵黃的合成是卵細胞體積增長的主要動力。卵子的大小可能會影響精卵的結合以及受精卵的發(fā)育, 據(jù)悉擬目烏賊受精卵產出時大小為 17.1 mm×39.7 mm, 孵化出的烏賊幼體胴長為8.0 mm, 而曼氏無針烏賊[38]受精卵產出時大小僅有5.5 mm×10.0 mm, 孵化出的烏賊幼體胴長僅有4.0 mm,這與擬目烏賊卵子較曼氏無針烏賊大是密切相關的。
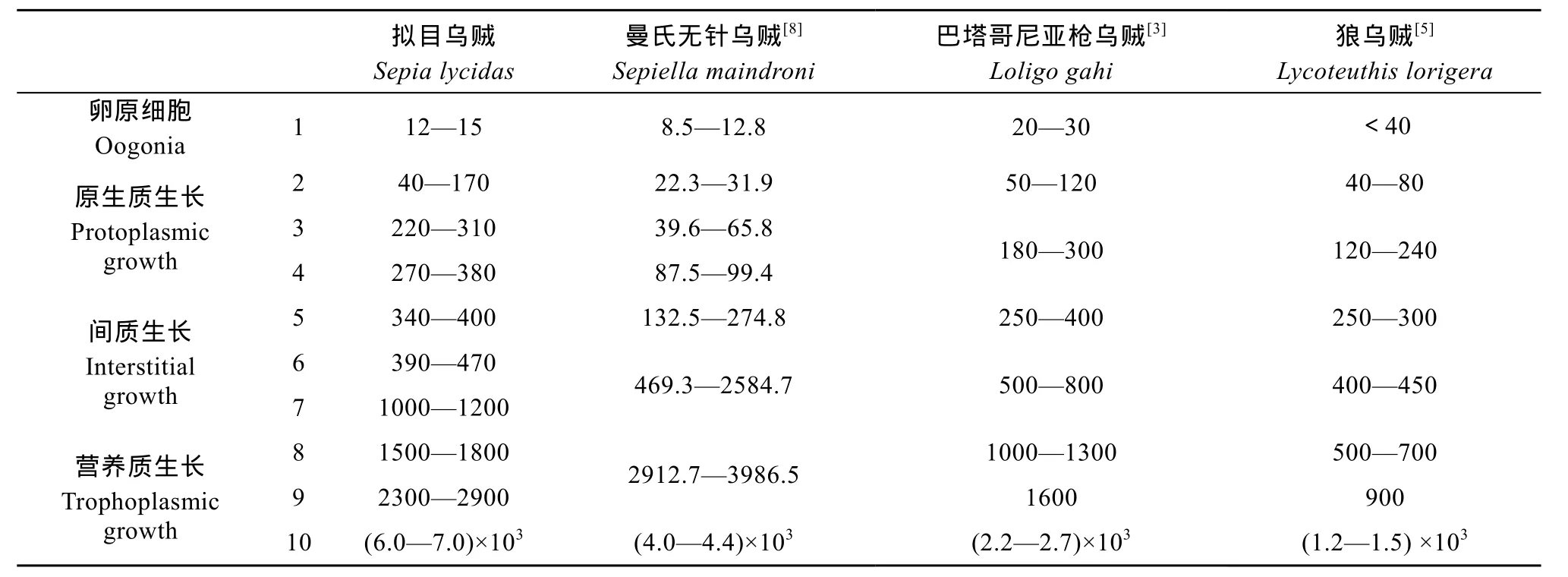
表2 幾種烏賊卵子發(fā)生過程中卵細胞直徑變化的比較(單位: μm)Tab. 2 Comparison of oocyte diameter of several cuttlefish during Oogenesis (Unit: μm)
細胞核的變化主要體現(xiàn)在體積和位置上, 在擬目烏賊卵子發(fā)生過程中, 細胞核體積不斷增長至第5期, 之后保持在(150—180) μm左右, 位置由細胞中央偏移到一側, 隨著濾泡細胞的解體而逐漸消失,這與皮氏槍烏賊[2]、巴塔哥尼亞槍烏賊[3]和狼烏賊[5]卵子發(fā)生過程中細胞核的變化相類似, 而曼氏無針烏賊[8]卵子發(fā)生過程中細胞核消失之前體積有一個減小的過程。在整個卵子發(fā)生過程中, 核質比是不斷減小的, 這與倪海爾和杜立勤[39]認為卵母細胞越趨向成熟, 核質比越小的觀點一致。
