過表達GBA誘導SH-SY5Y細胞自噬和降低α-synuclein的表達
2014-11-27 07:32:02范成河王雪晶楚廣磊馬耀華滕軍放
基礎醫學與臨床 2014年4期
關鍵詞:基因突變
范成河,王雪晶,楚廣磊,馬耀華,荊 婧,滕軍放
(鄭州大學 第一附屬醫院 神經內科, 河南 鄭州 450052)
過表達GBA誘導SH-SY5Y細胞自噬和降低α-synuclein的表達
范成河,王雪晶,楚廣磊,馬耀華,荊 婧,滕軍放*
(鄭州大學 第一附屬醫院 神經內科, 河南 鄭州 450052)
目的探討葡萄糖腦苷脂酶(GBA)對SH-SY5Y細胞自噬及α-突觸核蛋白(α-synuclein)表達水平的影響。方法將EGFP-N1、EGFP-GBA質粒轉染SH-SY5Y細胞,Western blot檢測Beclin 1、LC3-Ⅱ/LC3-Ⅰ及α-synuclein蛋白的表達,熒光顯微鏡觀察自噬泡的形成及LC3與GBA在細胞內的共定位。結果與轉染EGFP-N1組相比,轉染EGFP-GBA組上調Beclin 1和LC3-Ⅱ/LC3-Ⅰ 蛋白的表達水平(Plt;0.05),而α-synuclein表達水平則下降(Plt;0.05);與轉染EGFP-N1組相比,轉染EGFP-GBA組自噬空泡數目明顯增加,外源性LC3免疫熒光表達明顯增強,且GBA與LC3存在共定位。結論GBA上調自噬水平,降低α-synuclein蛋白的表達。
葡萄糖腦苷脂酶;α-突觸核蛋白;自噬
大量病理的、遺傳的及臨床的研究都證實葡萄糖腦苷脂酶基因(glucocerebrosidase gene, GBA)基因突變是帕金森病及相關疾病的重要危險因素[1]。帕金森病(Parkinson’s disease, PD)的病理標志是黑質致密部選擇性多巴胺神經元的丟失及殘存的神經元內蛋白聚集體的形成,而α-突觸核蛋白是這些蛋白聚集體的主要組成部分,PD的發生、發展以及受累腦區的變性均與α-突觸核蛋白的異常聚集有密切的聯系[2-3]。GBA基因的突變可促進α-突觸核蛋白相互聚集并形成穩定的寡聚體,積聚的α-突觸核蛋白影響了GBA蛋白的活性,而GBA活性的改變反過來又影響了α-突觸核蛋白的清除,由此可見α-突觸核蛋白與GBA蛋白的缺乏在PD的發病中起到了一個雙向環路的作用[4]。自噬-溶酶體途徑在清除異常聚集的α-突觸核蛋白中發揮著重要作用[5],GBA是溶酶體內的一種酶,且GBA與α-突觸核蛋白可相互影響,因此有學者推測GBA蛋白在自噬-溶酶體和調控α-突觸核蛋白的表達中發揮著某種作用。為此,設計本實驗以探討GBA對自噬的調控及對α-突觸核蛋白表達水平的影響。
1 材料與方法
1.1 材料
細胞及試劑:質粒EGFP-N1、EGFP-GBA和RFP-LC3(蘇州大學藥學院王光輝教授饋贈),人神經母細胞瘤細胞(SH-SY5Y)(中南大學湘雅醫院唐北沙教授饋贈),DMEM培養基、胎牛血清(FBS)(Gibco公司),抗LC3抗體(Abcam公司)、抗GAPDH抗體(Santa Cruz Biotechnology公司)、抗Beclin抗體(Eptimics公司),過氧化物酶標記山羊抗兔IgG(Promega公司),Lipofectamine2000、單丹磺酰尸胺(monodan-sylcadaverine,MDC)和Hochest33258(Sigma公司),LysoTracker(Invitrogen公司)。
1.2 方法
1.2.1 細胞培養及轉染:SH-SY5Y細胞常規培養于含有體積分數15%新鮮胎牛血清的DMEM完全培養液中,置于37 ℃、體積分數5% CO2細胞培養箱中培養,細胞以4×108個/L接種于6孔板里,次日轉染,轉染具體操作步驟參考脂質體LipofectamineTM2000說明書進行。
1.2.2 MDC染色:MDC是一種化學熒光染料,可以標記自噬空泡,通過其染色可觀察判斷細胞自噬的發生。SH-SY5Y細胞分別轉染EGFP-N1、EGFP-GBA 24 h后,用質量分數4%多聚甲醛室溫固定10 min,磷酸鹽緩沖液(1×PBS)洗3次,加入終濃度為0.05 mmol/L的MDC染色液,37 ℃孵育1 h后加入1×PBS洗3遍,在熒光倒置顯微鏡(Olympus 1×70)下觀察熒光情況,并在400倍視野下,各組細胞隨機抽取20個不相重復的視野進行觀察拍照。
1.2.3 細胞免疫熒光化學:將SH-SY5Y細胞按1∶4接種比例接種至6孔板進行培養,次日轉染后置于37 ℃、體積分數5% CO2細胞培養箱中培養48 h,棄去培養基,1×PBS洗2遍,用質量分數4%的多聚甲醛,室溫下固定10 min,1×PBS洗3遍,0.25%聚乙二醇辛基苯基醚(Trito X-100)封閉10 min,1×PBS洗3遍,Hochest33258(1∶10 000)染細胞質,室溫下避光孵育3 min,1×PBS洗3次,用熒光倒置顯微鏡觀察并分析熒光情況(568 nm激發,探測RFP紅色熒光;488 nm激發,探測EGFP綠色熒光),并在400倍視野下,各組細胞隨機抽取20個不相重復的視野觀察和拍照。
1.2.4 蛋白印跡(Western blot):SH-SY5Y細胞轉染48 h后,收集細胞提取總蛋白制成所需樣品;十二烷基硫酸鈉聚丙烯酰胺凝膠電泳(SDS-PAGE)后轉移到聚偏二氟乙烯(PVDF)膜上;PVDF膜用5%脫脂奶粉室溫封閉1 h,TBST洗3次,加入抗Beclin1(1∶800)、抗α-synuclein(1∶1 000)、抗LC3(1∶300)抗體,4 ℃過夜。次日加入HRP結合的二抗(山羊抗兔IgG,1∶5 000),室溫孵育2 h后行ECL光化學法顯色,暗室下行X線膠片曝光,內參為GAPDH。各組Western blot蛋白條帶用AlphaEaseFC(FluorChem8900)軟件分析圖像,采用平均吸光度值比較各組蛋白的表達。
1.3 統計學分析
2 結果
2.1Westernblot檢測細胞內源性Beclin1、LC3-Ⅱ/LC3-Ⅰ、α-synuclein的表達
與轉染EGFP-N1組相比,轉染EGFP-GBA組內源性Beclin 1蛋白的表達明顯上調(Plt;0.05),且LC3-Ⅱ/LC3-Ⅰ比值增高(Plt;0.05),而α-synuclein表達減少(Plt;0.05)(圖1,表1)。
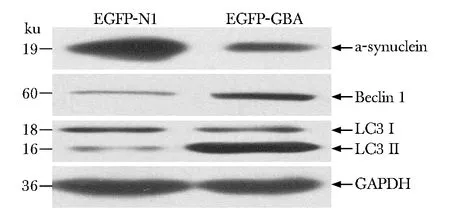
圖1 過表達GBA后對α-synuclein、Beclin1和LC3蛋白的影響Fig 1 The influence of GBA overexpression on α-syn- uclein, Beclin1 and LC3 protein expression
表1 GBA對α-synuclein、Beclin 1、LC3-Ⅱ/LC3-Ⅰ蛋白表達的影響Table 1 Effect of GBA on protein of α-synuclein, Beclin 1 and the ratio of LC3-Ⅱ/LC3-Ⅰ
*Plt;0.05 compared with EGFP group.
2.2 免疫熒光觀察MDC和Lysotracker染色
與轉染HA組相比,轉染HA-GBA組MDC染色信號增強,自噬空泡聚集增多(圖2A)。同時利用溶酶體標志物LysoTracker Red DND-99來觀察溶酶體熒光強度的變化(圖2B)。
2.3轉染后觀察細胞內RFP-LC3和EGFP-GBA的免疫熒光信號及共定位
與轉染EGFP組相比,轉染EGFP-GBA組RFP-LC3的免疫熒光信號明顯增強,外源性LC3的表達明顯上調,自噬水平上調;且RFP-LC3和EGFP-GBA存在亞細胞共定位(圖3)。
3 討論
研究證實GBA基因是PD重要的危險因素,其與PD發病機制的關系主要有兩種假設:一種是GBA基因突變導致蛋白錯誤折疊,抑制了泛素-蛋白酶體系統的活性,導致異常蛋白的降解減少[6];另一種是GBA基因突變導致GBA在體內聚集,引起自噬障礙或溶酶體機能不全,使α-突觸核蛋白的聚集增多[7]。GBA蛋白與α-突觸核蛋白在體內外都存在相互作用,且二者共定位在組織蛋白酶D陽性的細胞結構內[8]。在一種GBA基因突變的鼠模型的海馬區域中發現,GBA基因的突變和下調GBA的表達都能上調α-突觸核蛋白的表達[9];在本實驗中,過表達GBA使α-突觸核蛋白的表達水平明顯降低,這與GBA基因突變導致GBA缺乏引起α-突觸核蛋白聚集、濃度增加的結果相對應。
越來越多的研究證實,自噬調節途徑的病理改變可能是易聚集蛋白(如α-突觸核蛋白)在受損神經元內積聚的主要原因,與PD等神經系統變性疾病的發生、發展密切相關[10-11]。當自噬發生時,LC3-Ⅱ/LC3-Ⅰ比值明顯增高[12-13],Beclin1表達明顯上調[14]。在一個過表達GBA基因突變(D409V)的動物模型研究中發現,GBA基因的突變導致α-突觸核蛋白的聚集, 但這種聚集可被自噬誘導劑雷帕霉素逆轉[9]。 在該實驗中,過表達GBA使Beclin1蛋白表達明顯上調,LC3-Ⅱ/LC3-Ⅰ比值增高,且MDC染色結果也顯示細胞內自噬空泡數目明顯增加,溶酶體標志物也顯示溶酶體的數量明顯增加。共轉染HA-GBA和RFP-LC3,LC3熒光強度增加,且HA-GBA和RFP-LC3存在明顯的共定位,由此表明過表達GBA可誘導細胞自噬的發生。
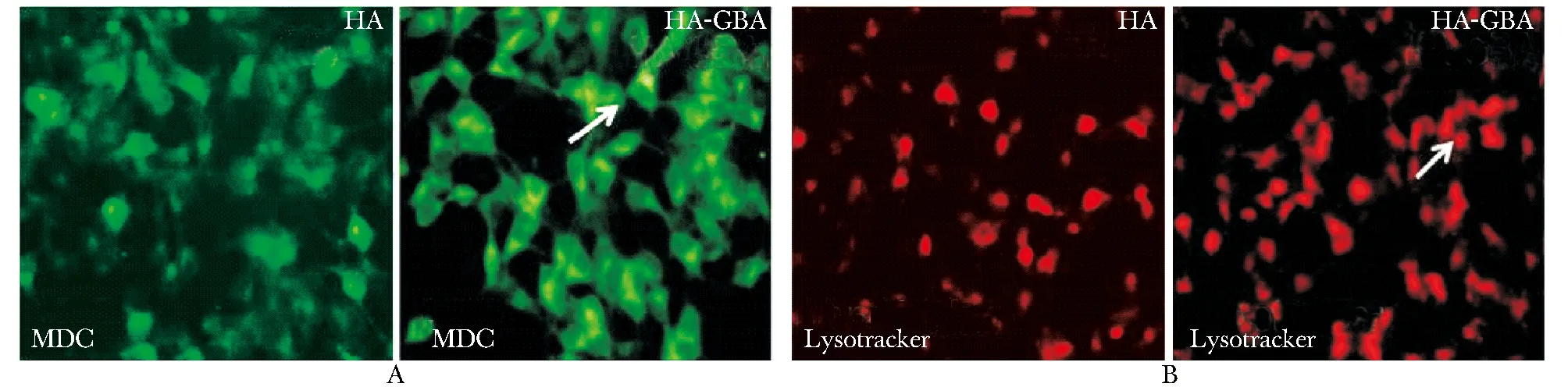
A.in GBA overexpressing groups,the bright green dots of MDC staining were increased in the morphologic picture, which marked autophagy; B.immunofluorescene showed that the bright red dots of Lysotracker staining were increased in GBA overexpressing groups
圖2共聚焦倒置顯微鏡觀察MDC和Lysotracker染色情況
Fig2ConfocalfluorescencemicroscopyimageofMDCandLysotrackerstaining(×400)
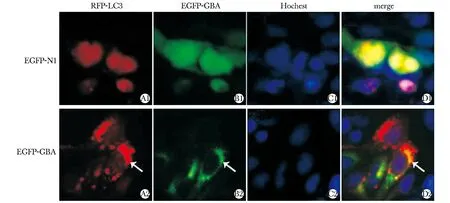
Immunoflourescence analysis showed that GBA(green) colocalized with LC3(red) in the cytoplasm of SH-SY5Y cells圖3 GBA和LC3在SH-SY5Y細胞中的共定位關系Fig 3 Co-localization of GBA and LC3 in SH-SY5Y cells(×400)
過表達GBA既可以上調自噬水平,又能使α-突觸核蛋白表達降低,二者是否存在關聯呢?自噬可通過降解那些觸發早期階段細胞死亡的損傷的異常蛋白(如α-突觸核蛋白),從而對神經元起保護作用,而抑制神經元內的自噬則會導致蛋白聚集體的積聚和神經元的變性。本實驗發現過表達GBA可以誘導自噬,自噬又是降解α-突觸核蛋白的重要途徑,且GBA與α-突觸核蛋白共定位在組織蛋白酶D陽性的細胞結構內[8],由此推測過表達的GBA可能部分通過自噬降低α-突觸核蛋白的表達水平,但該推測及具體的機制還需要進一步的研究證實。
[1] Sidransky E, Nalls MA, Aasly JO,etal. Multicenter analysis of glucocerebrosidase mutations in Parkinson’s disease[J]. N Engl J Med, 2009, 361: 1651-1661.
[2] 翟驍,段國禮,沈下賢,等. α-突觸核蛋白在帕金森病中的作用[J]. 基礎醫學與臨床,2012, 32: 1481-1483.
[3] Spillantini MG, Schmidt ML, Lee VM,etal. AlpHA-synuclein in Lewy bodies[J]. Nature, 1997,388:839-840.
[4] Tan EK, Tong J, Fook-Chong S,etal. Glucocerebrosidase mutations and risk of Parkinson disease in Chinese patients[J]. Arch Neurol, 2007,64:1056-1058.
[5] Pan T, Kondo S, Le W,etal. The role of autophagy-lysosome pathway in neurodegeneration associated with Parkinson’disease[J]. Brain, 2008, 131:1969-1978.
[6] Dawson TM. Parkin and defective ubiquitination in Parkinson’s disease[J]. J Neurla Transm Suppl, 2006, 70: 209-213.
[7] Wong K, Sidransky E, Verma A,etal. Neuropathology provides clues to the pathphysiology of Gaucher disease[J]. Mol Genet Metab, 2004, 82: 192-207.
[8] Yap TL, Gruschus JM, Velayati A,etal. AlpHA-synuclein interacts with glucocerebrosidase providing a molecular link between Parkinson and Gaucher diseases[J]. J Biol Chem, 2011,286: 28080-28088.
[9] Cullen V, Sardi SP, Ng J,etal. Acid beta-glucosidase mutants linked to Gaucher disease, Parkinson disease, and Lewy body dementia alter alpHA-synuclein processing[J]. Ann Neurol, 2011, 69: 940-953.
[10] 楊光,祝元剛,楊慧. 雷帕霉素抗神經退行性病研究進展[J]. 基礎醫學與臨床,2008, 28: 1336-1338.
[11] Cherra SJ, Chu CT. Autophagy in neuroprotection and neurodegeneration:a question of balance[J]. Future Neurol, 2008, 3:309-323.
[12] Klionsky DJ. Autophagy: from phenomenology to molecular understanding in less than a decade[J]. Nat Rev Mol Cell Biol, 2007, 8:931-937.
[13] Klionsky DJ,Cuervo AM, Seglen PO. Methods for monitoring autophagy from yeast to human[J]. Autophagy, 2007,3: 181-206.
[14] Luo S, Rubinsztein DC. Apoptosis blocks Beclin 1-dependent autophagosome synthesis: an effect rescued by Bcl-xL[J]. Cell Death Differ, 2010, 17: 268-277.
Overexpression of GBA induces autophagy and decreases α-synuclein expression in SH-SY5Y cells
FAN Cheng-he, WANG Xue-jing, CHU Guang-lei, MA Yao-hua, JING Jing, TENG Jun-fang*
(Dept. of Neurology, the First Affiliated Hospital of Zhengzhou University, Zhengzhou 450052, China)
ObjectiveTo investigate the mechanism of autophagy induced by GBA and the regulation of GBA on the expression of α-synuclein in SH-SY5Y cells.MethodsThe recombinated plamid EGFP-N1, EGFP-GBA were transfected into SH-SY5Y cells through eukaryotic cell transfection technique. The expression of beclin1, LC3-Ⅱ/LC3-Ⅰ and α-synuclein in the level of protein was detected through Western blot,MDC staining and cellular immunofluorescence microscopy were performed to observe the change of autophagic vacuolization and their subcellular colocalization.ResultsCompared to EGFP-N1 group, the expression of Beclin1 and LC3-Ⅱ/LC3-Ⅰ in group of overexpression EGFP-GBA were significantly increased, and the average number of autophagic vacuoles per cell increased, however, the expression of α-synuclein was significantly decreased (Plt;0.05). Besides, the signals for LC3, GBA and the extent of their co-localization increased in GBA groups.ConclusionsGBA protein decreases the expression of α-synuclein and induces autophagy of SH-SY5Y cells.
GBA; α-synuclein; autophagy
2013-07-18
2013-10-28
國家自然科學基金(81241046)
*通信作者(correspondingauthor): 13838210077@163.com
1001-6325(2014)04-0480-05
研究論文
R 741.02
A
猜你喜歡
英語世界(2023年6期)2023-06-30 06:29:10
中國醫學影像學雜志(2021年6期)2021-08-13 08:43:36
中國生殖健康(2020年2期)2021-01-18 02:51:26
小學生導刊(2018年13期)2018-06-29 03:49:00
中國生殖健康(2018年2期)2018-01-12 13:57:51
現代檢驗醫學雜志(2016年4期)2016-11-15 02:01:14
中國現代醫學雜志(2015年26期)2015-12-23 11:04:22
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:44
中華皮膚科雜志(2014年4期)2014-12-19 12:55:49
中國神經精神疾病雜志(2014年1期)2014-03-01 03:23:22
